CROP PROTECTION
Relationships between Four Neotropical Species of
Galling Insects and Shoot Vigor
SILMARY J. GONÇALVES-ALVIM, MAURÍCIO L. FARIAAND GERALDO W. FERNANDES
Ecologia Evolutiva de Herbívoros Tropicais/ DBG, ICB/ Universidade Federal de Minas Gerais, Caixa postal 486, 30161-970, Belo Horizonte, MG.
An. Soc. Entomol. Brasil 28(1): 147-155 (1999)
Relações entre Quatro Espécies de Insetos Galhadores Neotropicais e o Vigor de Ramos
RESUMO - A hipótese do vigor de plantas (HVP) prediz que plantas ou módulos de plantas mais vigorosos são preferencialmente selecionados por fêmeas de insetos galhadores como sítios de oviposição, pois aumentam a chance de sobrevivência da prole. A HVP foi testada para Neopelma baccharidis Buckhardt (Homoptera: Psyllidae) e Rhoasphondylia friburgensis Tavares (Diptera: Cecidomyiidae) que causam galhas em Baccharis dracunculifolia D.C. (Asteraceae), e para duas espécies diferentes de Cecidomyiidae (Diptera) que provocam galhas em Maytenus salicifolia Reiss. (Celastraceae) e Vernonia polyanthes Less (Asteraceae). A abundância de galhas aumentou com o aumento do comprimento do ramo nas quatro espécies, como previsto pela HVP. Entretanto, ao considerarmos a taxa de ataque por unidade de ramo (cm), que inclui nas análises o efeito da disponibilidade de recursos por unidade de comprimento, o padrão de resposta dos quatro galhadores desapareceu. R. friburgensis e o cecidomiídeo que induz galhas em M. salicifolia não responderam ao vigor do ramo (P > 0,05), enquanto as outras espécies responderam diferencialmente. A abundância de galhas de N. baccharidis correlacionou-se negativamente com o aumento do comprimento do ramo (y = 0,185 - 0,007x; r² = 0,21; F = 6,013, P < 0,05), enquanto o número de galhas provocadas pelo cecidomiídeo em V. polyanthes correlacionou-se positivamente com o vigor do ramo (y = 0,09 + 0,002x; r² = 0,34; F = 6,157; P < 0,05). Portanto, apenas o padrão de ataque do cecidomíideo galhador em V. polyanthes suportou a HVP.
PALAVRAS-CHAVE: Insecta, Diptera, Homoptera, galhas, interações inseto-planta.
species that induce galls on Maytenus salicifolia Reiss. (Celastraceae) and Ver-nonia polyanthes Less (Asteraceae), respectively. The abundance of galls in-duced by the four galling species increased with increasing shoot length, as predicted by the PVH. However, when we considered the rate of attack per shoot unit length (cm), which includes in the analyses the effect of availability of resource per unit length, the response patterns of the four gallers disappeared. R. friburgensis and Cecidomyiidae species which cause galls on M. salicifolia did not respond to shoot vigor (P > 0.05) while the other species responded differentially. The abundance of N. baccharidis galls correlated negatively with shoot length (y = 0.185 - 0.007x, r² = 0.21, F = 6.013, P < 0.05) while the number of galls caused by Cecidomyiidae on V. polyanthes showed a positive relationship with shoot vigor (y = 0.09 + 0.002x, r² = 0.34, F = 6.157, P < 0.05). Hence, only the attack pattern of the cecidomyiid galler on V. polyanthes cor-roborated the PVH.
KEY WORDS: Insecta, Diptera, Homoptera, galls, insect-plant interactions.
Studies on the effects of plant quality on the attack rates by herbivorous insects have shown contradictory results that have led to two major hypotheses. Many species show high preference and improved larval perform-ance on stressed hosts, which have resulted in the formulation of the “plant stress hypoth-esis” (PSH) ( e.g., White 1969, 1976). It pre-dicts that plants under abiotic stress become more suitable as food to herbivores due to an increase in the availability of nutrients. In addition, stressed plants may synthesize less chemical defenses (Rhoades 1979, White 1984). Some studies support White’s hypoth-esis (e.g., Waring & Cobb 1992, Bruyn 1995, Cobb et al. 1997). On the other hand, other studies have shown that several species of herbivorous insects respond positively to vig-orously growing plants or plant parts (e.g., Price et al. 1990, Hunter & Price 1992, Preszler & Price 1995), which have resulted in the “plant vigor hypothesis” (PHV) (Price 1991). Vigor is defined as any plant or plant module that grows rapidly and reaches a larger size in relation to the mean growth rate. Price (1991) postulated that insect herbivores, whose larval development is associated with their host-plant growth processes, should pre-fer to attack the most vigorous plants or plant modules where subsequent larval performance
is highest. The archetype herbivores with these characteristics are galling insects, whose association with host-plant vigor have impor-tant implications on their population dynam-ics (Preszler & Price 1995).
Price (1991) also suggested a solution for the divergence in the results of these two hy-potheses, proposing an alternative view in which there is a continuum of strategies. This approach predicts that insect herbivores more specialized and intimately related with the growth processes of plants (latent species) tend to select shoots at one extreme of the continuum, hence characterizing the PVH. Other herbivores, notably those with ovipo-sition and larval development spatially and temporally separated (eruptive species) tend to select modules at the other end of this con-tinuum, characterizing the PSH.
Faria et al. (1997) also verified a positive re-lationship between attack by a Scarabaeidae beetle and shoot length in Erythroxyllum suberosum Turcz. (Erythroxylaceae). Nev-ertheless, other studies have shown different results. Madeira et al. (1997) did not find any support for female preference for longer shoots, while Cornelissen et al. (1997) sug-gested idiosyncratic responses of the herbiv-ores that attack shoots of Bauhinia brevipes Vog. (Leguminosae).
In this study, we evaluate the pattern of attack by four galling insects on three species of host plants, Maytenus salicifolia Reiss. (Celastraceae), Baccharis dracunculifolia D.C. (Asteraceae) and Vernonia polyanthes Less (Asteraceae). The study aims to answer the following questions: Are the longest shoots in the available shoot population of the three plant species most attacked by the gallers? Do the four galling species respond similarly to plant vigor?
Material and Methods
Study Area. This study was done in the
Cam-pus of the Federal University of Minas Gerais (19º52’ S, 43º58’ W), Belo Horizonte (MG) in southeastern Brazil. The campus is at 800 m above sea level and has a heterogeneous vegetation with native, introduced and orna-mental species (Fernandes et al. 1988). The studied area has a marked dry period of 3 to 4 months with an average monthly precipitation of 50 mm and average temperature between 19ºC and 21ºC. The rainy season lasts eight to nine months and the average monthly pre-cipitation is 300 mm with an average tempera-ture of 23.9ºC.
Host Plants and Galling Insects. An uni-dentified species of Cecidomyiidae (Diptera) induces galls on the central leaf vein of M. salicifolia. Galls are spheroid, brownish, gla-brous, one-chambered and have one larva per chamber. These galls are frequently observed on the host plant throughout the year.
V. polyanthes is a weed species of culti-vated and disturbed areas (Leitão Filho et al.
1972). Spheroid galls are formed on the cen-tral vein of its leaves by an unidentified spe-cies of Cecidomyiidae (Diptera). Galls are green, glabrous, one-chambered with one or two larvae per chamber. They occur on the abaxial leaf surface and are very common in studied area.
Approximately 17 species of galling in-sects attack B. drancunculifolia (Fernandes et al. 1996 ). The host plant is a dioecious shrub which is widespread in Brazil. Two species of gallers were studied: Neopelma baccharidis Buckhardt (Homoptera: Psyllidae) is the most common galling insect on B. dracunculifolia which causes an elon-gated globular leaf gall (Lara & Fernandes 1994) and Rhoasphondylia friburgensis Tavares (Diptera: Cecidomyiidae) which pro-duces globular, green and glabrous galls on the stems of the plant (Gagné 1994).
Sampling and Statistical Analyses. Random
Results
Longer shoots were less abundant than shorter shoots on the three plant species. How-ever, high rates of attack by galling insects were always observed on the longest shoots (Fig. 1). The highest density of galls occurred
on M. salicifolia with 10,090 galls per 1,000 shoots. N. baccaridis induced 570 galls/1,000 shoots, R. friburguensis induced 215 galls/ 1,000 shoots, while the unidentified species of cecidomyiid induced 2,695 galls/1,000 shoots on V. polyanthes.
Galling insects showed divergent
re-Figure 1. Frequency distribution expressed as the percentage of total shoots ( ) and percentage of shoots galled ( ) in each shoot length class on A) B. dracunculifolia (Asteraceae) galled by R. friburgensis (Diptera: Cecidomyiidae); B) B. dracunculifolia (Asteraceae) galled by N. baccharidis (Homoptera: Psyllidae); C) V. polyanthes (Asteraceae) galled by Cecidomyiidae (Diptera) and D) M. salicifolia (Celastraceae) galled by Cecidomyiidae (Diptera).
3.5 5.5 7.5 9.5 11.5 13.5 15.5 17.5 20.5 23.5 25.5 34.5 0 0.02 0.04 0.06 0.08 0.1 0.12 0 0.2 0.4 0.6 0.8 1 1.2
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29
0 0.05 0.1 0.15 0.2 0.25 0.4 0.5 0.6 0.7 0.8 0.9 1 1.1 .5 2.5 4.5 6.5 8.5 10.5 12.5 14.5 16.5 18.5 20.5 26.5 0 0.02 0.04 0.06 0.08 0.1 0.12 0 0.2 0.4 0.6 0.8 1 1.2
2.8 6.2 9.8 13.2 16.8 20.2 23.8 27.2 30.8 34.2 37.8 41.2 44.8 48.2 0 0.02 0.04 0.06 0.08 0.1 0.12 0.14 0 0.2 0.4 0.6 0.8 1 1.2 A L L SHOO T S ( %
) AT
T A CKE D S HO OT S ( % )
SHOOT SIZE CLASS (cm)
A
B
C
D
sponses to shoot length classes. The attack by R. friburguensis did not correlate with shoot length (y = 0.082 + 0.021x, r² = 0.12, F = 2.934, P > 0.05). On the other hand, the number of galls per shoot positively corre-lated with shoot length in the other three gall-ing species: N. baccaridis (y = 0.183 + 0.043x,
r² = 0.52, F = 23.923, P < 0.05), cecidomyiid on V. polyanthes (y = - 0.77 + 0.19x, r² = 0.86, F = 71.617, P < 0.05), and cecidomyiid on M. salicifolia (y = 2.42 + 0.67x, r² = 0.30, F = 5.059, P < 0.05) (Fig. 2). These results cor-roborate the plant vigor hypothesis.
However, when the attack rates were
re-Figure 2. Observed distribution of the number of galls per shoot on A) B. dracunculifolia (Asteraceae) galled by R. friburgensis (Diptera: Cecidomyiidae); B) B. dracunculifolia (Asteraceae) galled by N. baccharidis (Homoptera: Psyllidae); C) V. polyanthes (Asteraceae) galled by Cecidomyiidae (Diptera) and D) M. salicifolia (Celastraceae) galled by Cecidomyiidae (Diptera).
5 10 15 20 25 30 35 40
0 0.5 1 1.5 2 2.5
5 10 15 20 25 30 35
0 0.5 1 1.5 2 2.5
12
GA
LLS
P
E
R
S
H
O
O
T
SHOOT SIZE CLASS (cm)
A
B
10 20 30 40 50 60
0 2 4 6 8
10 C
5 10 15 20 25 30
0 10 20 30 40 50
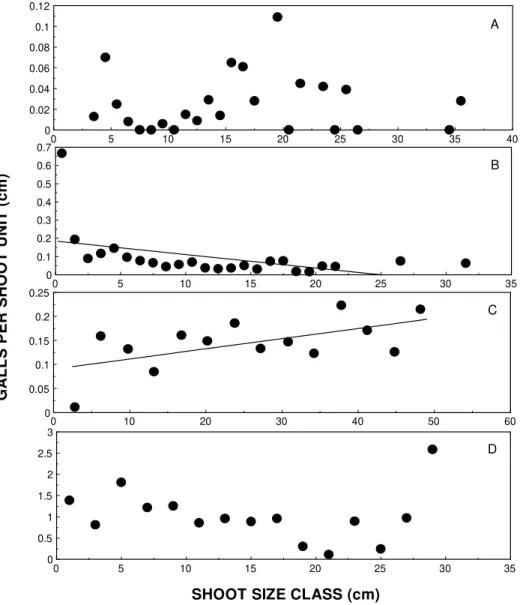
calculated as the number of galls per shoot unit length (cm), the patterns previously re-ported were modified (Fig. 3). The attack by N. baccaridis became negatively correlated with shoot length (y = 0.185 - 0.007x, r² = 0.21, F = 6.013, P < 0.05). The attack by the
cecidomyiid on V. polyanthes became posi-tively correlated with shoot length (y = 0.09 + 0.002x, r² = 0.34, F = 6.157, P < 0.05). The patterns of attack of R. friburguensis (y = 0.023, r² = 0.01, F = 0.038, P > 0.05) and that of the cecidomyiid on M. salicifolia (y = 1.13
Figure 3 . Observed distribution of the number of galls per shoot unit length (cm) on A) B. dracunculifolia (Asteraceae) galled by R. friburgensis (Diptera: Cecidomyiidae); B) B. dracunculifolia (Asteraceae) galled by N. baccharidis (Homoptera: Psyllidae); C) V. polyanthes (Asteraceae) galled by Cecidomyiidae (Diptera) and D) M. salicifolia (Celastraceae) galled by Cecidomyiidae (Diptera).
0 5 10 15 20 25 30 35 40
0 0.02 0.04 0.06 0.08 0.1 0.12
0 5 10 15 20 25 30 35
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
0 10 20 30 40 50 60
0 0.05 0.1 0.15 0.2 0.25
0 5 10 15 20 25 30 35
0 0.5 1 1.5 2 2.5 3
A
B
C
D
SHOOT SIZE CLASS (cm)
GA
LL
S
P
E
R
S
H
O
O
T
UNI
T
(c
- 0.008x, r² = 0.01, P > 0.05) did not show any relationship with shoot length. These re-sults indicate that the plant vigor hypothesis was not supported for most insect species when we considered the attack rate per shoot unit length.
Discussion
The distribution patterns of shoot sizes in the three species of plants corroborate with many other studies (e.g., Craig et al. 1986, Craig et al. 1989), including tropical species (e.g., Cornellissen et al. 1997, Madeira et al. 1997) where the longest shoots are always rare in the population of shoots. The longest shoots also tend to be more attacked by gall-ing insects than the shortest shoots. In the tropics, Madeira et al. (1997) studied galling insect preference on Baccharis concinna Barroso (Asteraceae) and observed that the herbivores preferred the most vigorous shoots. However, the longest shoots have a greater probability of being attacked than the smaller shoots even if no selection is at work. Longer shoots offer more leaves and buds to be used by gallers than shorter shoots (Madeira et al. 1997). Furthermore, other variables such as the availability of sites for colonization and shoot age tend to produce greater attack fre-quencies in longer shoots (Karban 1987). Therefore, we argue that resource availabil-ity per shoot unit length should be incorpo-rated when testing the plant vigor hypothesis to avoid repetition in the results of galling insect attacks. The grouping of shoot length into size classes may force the relationship where the longest shoots are always attacked due to chance alone (Cornellissen et al. 1997). When we considered attack rate per shoot unit length (cm), the four galling species did not respond similarly to plant vigor. Instead, each species responded differentially or did not respond at all to shoot vigor. Thus, the observed pattern which supported the vigor hypothesis disappeared after applying a cor-recting method. This work corroborates the study of Cornellisen et al. (1997) where the authors observed the existence of a continuum
of responses by herbivores to plants and that herbivores may have idiosyncratic responses to plant quality even when feeding on the same host.
Acknowledgments
We thank J. Lombardi for the identifica-tion of the plant species, G. R. Julião for field assistance, E. Marques for her good sugges-tions and two anonymous reviewers for their comments which improved the manuscript. This study was supported by CNPq (521772/ 95-8), FAPEMIG (1950/95) and International Foundation for Science (H-2487/1).
Literature Cited
Bruyn, L. De. 1995. Plant stress and larval performance of a dipterous gall former. Oecologia 101: 461-466.
Cobb, N. S., S. Mopper, C.A. Gehring, K. Chistensen & T.G. Whitham. 1997.
Increased moth herbivory associated with environmental stress of pinyon pine at local and regional levels. Oecologia 109: 389-397.
Cornelissen, T. G., B. G. Madeira, L.R. Allain, A.C.F. Lara, L.M. Araújo & G.W. Fernandes. 1997. Multiple
responses of insect herbivores to plant vigor. Ciênc. Cult. 49: 285-288.
Craig, T.P., P.W. Price & J.K. Itami. 1986.
Resource regulation by a stem-galling sawfly, Euura lasiolepis, on the arroyo willow, Salix lasiolepis. Ecology 67: 419-425.
Craig, T.P., J.K. Itami & P.W. Price. 1989.
A strong relationship between oviposition preference and larval performance in a shoot-galling sawfly. Ecology 70: 1691-1699.
selection by a Scarabaeidae beetle on Erythroxyllum suberosum in Serra do Cipó, Minas Gerais, Brazil. Ecotropicos 10: 29-32.
Fernandes, G.W., E. Tameirão Neto & R.P. Martins. 1988. Ocorrência e caracterização das galhas entomógenas na vegetação do Campus-Pampulha, UFMG, Belo Horizonte-MG. Rev. Bras. Zool. 5: 11-19.
Fernandes, G.W., M. A. A. Carneiro, A.C.F. Lara, L. A. Allain, T.C. Reis & I.M. Silva. 1996. Galling insects on neotropical species of Baccharis (Asteraceae). Trop. Zool. 9: 315-332.
Gagné, R. J. 1994. The gall midges of the Neotropical Region. Ithaca, NY, Comstock.
Hunter, M.D. & P.W. Price. 1992. Playing chutes and ladders: heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology 73: 724-732.
Karban, R. 1987. Herbivory dependent on plant age: a hypothesis based on acquired resistance. Oikos 48: 336-337.
Lara, A.C.F. & G.W. Fernandes. 1994.
Distribuição de galhas de Neopelma baccharidis (Homoptera: Psyllidae) em Baccharis dracunculifolia (Asteraceae). Rev. Bras. Biol. 54: 661-668.
Leitão-Filho, H.F., C. Aranha & O. Bacchi. 1972. Plantas invasoras de culturas no
Estado de São Paulo. São Paulo, Hucitec.
Madeira, B.G., T.G. Cornelissen, M.L. Faria & G.W. Fernandes. 1997. Insect
herbivore preference for plant sex and modules in Baccharis concinna Barroso (Asteraceae). p. 135-143. In A. Raman (ed.), Ecology and Evolution of Plant-Feeding Insects in Natural and
Man-Made Environments. New Delhi, India, International Scientific Publications.
Prada, M., O.J. Marini-Filho & P.W. Price. 1995. Insects in flower heads of Aspilia foliaceae (Asteraceae) after a fire in a central Brazilian savanna: evidence for the plant vigor hypothesis. Biotropica 27: 513-518.
Preszler, R.W. & P.W. Price. 1995. A test of plant-stress, and plant-genotype effects on leaf-miner oviposition and performance. Oikos 74: 485-492.
Price, P.W. 1991. Plant vigor hypothesis and
herbivore attack. Oikos 62: 244-251.
Price, P.W., N. Cobb, T.P. Craig, G.W. Fernandes, J.K. Itami, S. Mopper & R.W. Preszler. 1990. Insect herbivore population dynamics on trees and shrubs: new approaches relevant to latent and eruptive species and life table development. p. 1-38. In E. A Bernays (ed.), Insect-plant interactions. Boca Raton, Florida, CRC Press.
Rhoades, D.F. 1979. Evolution of plant chemical defense against herbivores. p. 3-54. In G. A. Rosenthal & D. H. Janzen (eds.), Herbivores: their interactions with secondary plant metabolites. New York, Academic.
Vieira, E.M., I. Andrade & P.W. Price. 1996. Fire effects on a Palicourea rigida (Rubiaceae) gall midge: a test of the plant vigor hypothesis. Biotropica 28: 210-217.
Waring, G.L. & N.S. Cobb. 1992. The
impact of plant stress on herbivore population dynamics. p. 167-227. In E. A. Bernays (ed.), Insect-plant inter-actions. Boca Raton, Florida, CRC Press.
White, T.C.R. 1969. An index to measure
with outbreaks of psyllids in Australia. Ecology 50: 905-909.
White, T.C.R. 1976. Weather, food and
plagues of locusts. Oecologia 22: 119-134.
White, T.C.R. 1984. The abundance of
invertebrate herbivores in relation to the availability of nitrogen in stressed food plants. Oecologia 63: 90-105.