In vitro initial immune response against Leishmania amazonensis infection is characterized by an
Texto
Imagem
Documentos relacionados
Para chegar a este intento, o estudo buscou as motivações que levaram o empreendimento a adotar o cultivo urbano em suas dependências; conhecer os produtos cultivados
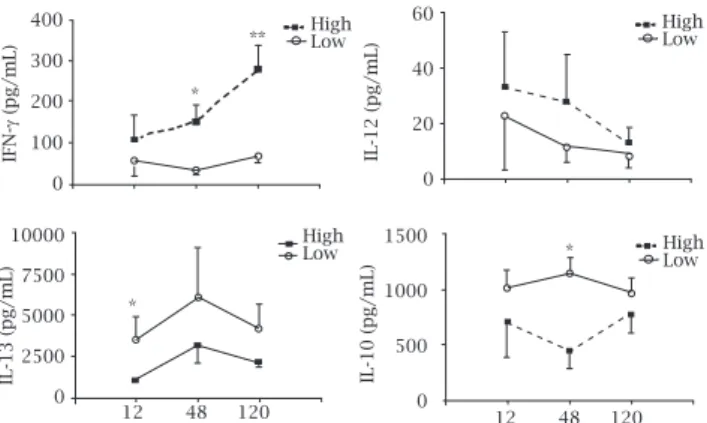
To investigate the mechanisms by which the decreased expres- sion of ecto-NTPDase activity contributes to lesion development in the host, mice were inoculated with short- or
Therefore, both the additive variance and the deviations of dominance contribute to the estimated gains via selection indexes and for the gains expressed by the progenies
In order to determine the in vitro nature of the association between ketoconazole and antimony on Leishmania (Leishmania) amazonensis amastigote, the fixed-rate isobologram
In this paper we evaluated the immune response of infected humans against two antigenic preparations: the total soluble extract (FhTSE) and the adult worm vomit (FhAWV) in order
We evaluated the expression of this molecule in logarithmic and stationary growth phase promastigotes, as well as in amastigotes, of Leishmania amazonensis by immunoblotting,
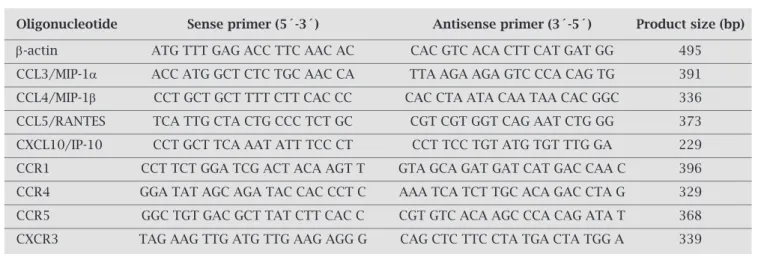
in the early acute phase of infection (5). Since a) IFN- γ produced by NK cells in the early acute phase of T. cruzi infection is important in mediating resistance, b) IL-12 has
As part of the strategies to advance in studies related with in situ detection of the virus proteins, we attempted to promote the in vitro expression of the ORF 3, the
