CASTANHEIROS
Selecção clonal na variedade Judia
Tese apresentada para o efeito de obtenção do grau de Doutor em Ciências Agronómicas e Florestais, de acordo com o disposto no nº 2 do artigo 8 do Decreto-Lei nº388/70 (Decreto-Decreto-Lei nº216/92, de 13 de Outubro)
Orientadores: Doutor José Carlos Esteves
Gomes-Laranjo, Professor Auxiliar da Universidade de Trás-os-Montes e Alto Douro, Doutor Francisco Manuel Peixoto, Professor Agregado da Universidade de Trás-os-Montes e Alto Douro e Doutora Rita Costa, Investigadora do INRB L-INIA.
AGRADECIMENTOS
Ao apresentar este trabalho não poderia deixar de expressar a minha gratidão a todos aqueles que, de forma diferente, contribuiram para a sua conclusão. A todos eles quero manifestar o meu sincero agradecimento.
Ao orientador desta dissertação, Professor Doutor José Carlos Esteves Gomes-Laranjo o meu agradecimento pela ajuda no meu crescimento científico e pela importante colaboração no planeamento e execução de todo o trabalho.
Ao meu co-orientador, Professor Doutor Francisco Manuel Peixoto pela indispensável disponibilização do laboratório que permitiu desenvolver parte dos estudos realizados e pelo sempre presente estímulo e rigor científico.
À Doutora Rita Maria Lourenço da Costa, Responsável do Laboratório de Biologia Molecular – INRB/L-INIA, o meu muito obrigada pela constante confiança depositada no meu trabalho, palavras de apoio e incentivo bem como pela disponibilidade com que sempre me presenteou.
Ao Professor Doutor Roberto Botta, pela ajuda no planeamento, execução e rigor científico no estudo da fenologia e biologia floral.
Ao Professor Doutor Jorge Ventura Ferreira Cardoso, pelo indispensável auxílio e orientação nas tarefas relacionadas com a bioquímica e morfologia do fruto.
Ao Professor Doutor João Coutinho, o meu agradecimento pela valiosa ajuda quer na execução quer no tratamento e discussão dos dados relativos à análise de solos e análise da composição mineral.
Ao Professor Doutor Fernando Hermânio Ferreira Milheiro Nunes, pelo auxílio no desenvolvimento de algumas metodologias relacionadas com a componente química do estudo.
À Professora Doutora Maria Manuel Oliveira, pela ajuda nos trabalhos de laboratório relativamente ao estudo da actividade antioxidante bem como pelo seu humor e constante incentivo.
Aos Professores Doutores Carlos Gomes Abreu e Luís Martins pelas variadas e valiosas contribuições nos trabalhos relacionados com a “doença da tinta”.
À Professora Doutora Teresa Pinto e ao Professor Doutor José Moutinho Pereira pela disponibilidade demonstrada no esclarecimento e planeamento de algumas tarefas ao longo do trabalho.
Ao Professor Doutor António Luis Crespí e ao Doutor Richard Nail Bennett pela correcção da escrita de alguns trabalhos científicos.
À Maria Natália Teixeira que, além de funcionária do DeBA, sempre se manifestou como uma grande amiga. Sinto-me profundamente grata pela total disponibilidade e vontade com que sempre me ajudou, pelas palavras e gestos de carinho, amor e incentivo que sempre me dirigiu.
A todos os funcionários do DeBA, nomeadamente ao Rui Martins, Helena Ferreira, Maria Irene Santos, Cesaltina Carvalho, Luís Fernando, Donzília e Ana Fraga, e à Ana Macedo da Protecção de Plantas por toda a ajuda nos trabalhos de laboratório e de campo.
A todos os estagiários com quem tive o prazer de trabalhar que de alguma forma permitiram o meu crescimento científico e social, nomeadamente à Marisa e ao Jorge.
Um agradecimento à FCT pelo apoio financeiro da minha bolsa de doutoramento. À minha família, particularmente aos meus Pais e Irmãos que nunca pouparam esforços para a minha educação e pelas dificuldades por que passaram por acreditarem que eu chegaria aqui. Muito obrigada pelo amor e palavras constantes de conforto.
Aos meus sogros e a todos os meus amigos, pelo carinho e amizade que sempre me dedicaram.
E como os últimos são os primeiros, ao meu marido Nuno João Pires Barbosa, agradeço o incentivo, a paciência, a companhia, as palavras de incentivo e os gestos repletos de carinho e amor com que sempre me presenteou.
RESUMO
O castanheiro é uma espécie multifuncional, cujo fruto, durante séculos, fez parte da dieta alimentar de muitos povos das regiões de montanha onde era cultivado, como sucedeu nos países do sul da Europa. Esta tendência foi evoluindo ao longo dos tempos e de alimento base de populações rurais, a castanha passou também a ser consumida pelas populações mais urbanas através de receitas tradicionais ou da alta cozinha. Finalmente, o conhecimento adquirido sobre o seu valor alimentar tem-na transformado num alimento dietético, a ponto de ser conhecida como uma “unNut”. Entre outros aspectos, é recomendada na alimentação para pessoas quer com problemas de diabetes quer celíacos. De entre as 13 espécies do género Castanea, a “europeia” é a Castanea sativa Mill. Infelizmente as alterações climáticas, principalmente a temperatura e o stresse hídrico, parecem condicionar o seu desenvolvimento em muitas regiões europeias.
Assim, pretendeu-se com este trabalho conhecer melhor a forma como diferentes condições edafoclimáticas poderão no futuro influenciar o desenvolvimento e consequente produtividade desta espécie em Trás-os-Montes, com destaque para a variedade Judia.
A tese está estruturada em dez capítulos. No primeiro é feita uma Introdução Geral (Capítulo 1). Seguidamente, é fornecida informação detalhada sobre cada uma das actividades de investigação desenvolvidas (Capítulos 2 a 9), terminando com o Capítulo 10 que é dedicado a uma breve síntese dos resultados essenciais, conclusões e importância deste estudo em futuros trabalhos.
Com a colaboração dos agricultores, e de associações de produtores de Trás-os-Montes, foram seleccionadas árvores da variedade Judia (5080 frutos/kg) em sete localidades: Alfândega da Fé, Bragança, Chaves, Macedo de Cavaleiros, Murça, Valpaços e Vinhais.
No primeiro ano (2006), foram seleccionadas 130 árvores de acordo com informações dos agricultores. Destas, tendo em conta os calibres (total: todos os frutos e correcto: só frutos viáveis) foram seleccionadas (Capítulo 2) as 25 melhores (Alfândega da Fé: 4, Bragança: 4, Chaves: 4, Macedo de Cavaleiros: 3, Murça: 2, Valpaços: 4 e Vinhais: 4). A distância genética entre as árvores (entre 0 e 0,227), obtida por genotipagem com marcadores moleculares, demonstrou a proximidade entre genótipos, provando que todas pertenciam à mesma variedade.
No segundo ano, o trabalho continuou com as 25 árvores seleccionadas no ano anterior. Os somatórios de graus-dia (ºD) de 2006 e 2007 foram 2551ºD e 2155ºD (Maio-Outubro), respectivamente. Em 2007, os frutos foram 38% maiores e contendo mais 50% de amido que os frutos colhidos em 2006. No entanto, o teor em gordura bruta foi 6,41% mais baixo. Em 2006, os frutos provenientes da localidade mais fria (Valpaços: 2300ºD) foram os maiores (62,5 frutos/kg) e os que apresentaram teores mais elevados de amido (400,7g/kg MS). Contrariamente, os frutos provenientes da localidade mais quente (Murça: 2751ºD), obtiveram o pior calibre (157,1 frutos/kg). Em 2007, os frutos de melhor calibre corrigido foram os de Macedo de Cavaleiros (2163ºD; 46,4 frutos/kg) tendo sido novamente os frutos de Murça a apresentarem pior calibre (66,0 frutos/kg).
Paralelamente, em 2007, foi feita a caracterização morfológica e histológica foliar das árvores de cada um dos 7 locais em estudo (Capítulo 3). Ainda no âmbito deste capítulo foi também feito o estudo fenológico. Neste capítulo, verificámos a adaptação morfológica e histológica de folhas de castanheiro às diferentes altitudes e condições climáticas. As altitudes variaram entre 709m e 860m acima do nível do mar, correspondendo a uma variação de valores de graus-dia (ºD) de 2751ºD para 2316ºD em 2006 e 2338ºD a 1700ºD em 2007 (entre Maio e Outubro, novamente). Os resultados gerais sugerem que as melhores condições de temperatura para a produção de fruto situam-se no intervalo 2100ºD/2200ºD (entre Maio e Outubro). Os locais onde este valor pode ser encontrado dependem das condições climáticas de cada ano. As alterações fenológicas verificadas entre locais foram registadas somente em 2007, no entanto o início do ciclo vegetativo bem como o início e o fim quer da floração feminina quer masculina variam consoante as localidades (mais quentes ou mais frias).
Baseados no estudo fenológico anterior (2007) e com intuito de obter melhor conhecimento sobre a influência da temperatura na biologia floral da variedade Judia, foram feitas polinizações controladas em 2008 (Capítulo 4). A polinização e crescimento do embrião são fortemente influenciados pelas condições climáticas que variam de local para local com diferentes altitudes e de ano para ano, sendo muitas vezes a principal causa da variabilidade na qualidade e quantidade da produção. Por outro lado, a fertilização e o desenvolvimento embrionário são etapas importantes que influenciam o rendimento e a qualidade da castanha, incluindo a percentagem de polispermia do fruto. As polinizações controladas da variedade Judia foram feitas com o pólen de Negral e Lada, duas variedades polinizadoras
compatíveis. Através das observações histológicas verificámos que a meados de Julho já tínhamos óvulos em desenvolvimento e continham já a célula mãe do megásporo. As flores de Judia polinizadas com Lada apresentaram óvulos maduros cinco dias antes das polinizadas com Negral. A influência dos polinizadores na polispermia e na formação de sulcos ainda não é inconclusiva, mas o tempo de polinização parece ser fundamental para a formação de frutos. Relativamente ao tamanho dos frutos obtidos através das polinizações controladas, a árvore cujo polinizador foi a variedade Negral apresentou melhor calibre que os resultados obtidos com a variedade Lada (168.2 e 281 frutos/Kg, respectivamente). A forma do fruto também apresentou alguma variação, uma vez que a variedade Negral parece originar, frutos mais alongados que a variedade Lada.
Nesse mesmo ano (2008, mais frio que os anteriores), foi também feita a análise do estudo da influência da temperatura nos metabolitos, ácidos gordos e propriedades tecnológicas, nas árvores das sete localidades (Capítulo 5). Os ºD variaram entre 1597ºD e 2077ºD. Neste estudo demonstrámos a correlação existente entre ºD e o teor de água das castanhas, proteína bruta e ácidos gordos. As localidades mais frias (1597ºD e 1971ºD), num ano frio (valor médio = 1959ºD), obtiveram frutos com teores mais elevados de água, proteína bruta, açúcares solúveis e amido e uma clara prevalência de ácidos gordos polinsaturados. Um estudo sobre a perda de água em frutos com casca mostrou diferenças significativas entre localidades. A localidade com ºD mais baixo apresentou maior percentagem de perda de água que a localidade com o maior valor de graus-dia. Esta diferença de valores reflecte maiores ou menores perdas a nível económico, respectivamente. Os melhores calibres
foram obtidos em Alfândega da Fé cujo valor do somatório de temperatura foi 2032ºD.
O Capítulo 6 destinou-se ao estudo da influência da temperatura na actividade antioxidante dos frutos colhidos em 2008. Os teores de fenóis totais variaram entre 9.6 mg/g de equivalentes em ácido gálico (GAE) na localidade mais quente (Murça) e 19.4 mg/g de GAE (ecotipo mais frio, Valpaços). O ácido gálico e o ácido elágico foram os compostos fenólicos predominantes, e mais uma vez a localidade mais fria obteve os maiores valores comparativamente à localidade mais quente, cujos valores obtidos foram os mais baixos. A actividade antioxidante foi avaliada através de diferentes ensaios: captação dos radicais de ABTS (ácido 2,2’-azinobis-(3-etilbenzotiazolina-6 sulfónico))e DPPH (2,2-difenil-2-picril-hidrazil), FRAP (poder antioxidante de redução do ferro) e inibição da hemólise oxidativa de eritrócitos. Os frutos provenientes de Valpaços (localidade mais fria), revelaram as melhores propriedades antioxidantes. Assim, as condições climáticas parecem ser um factor limitante para produção de compostos fenólicos e consequentemente para a actividade antioxidante do fruto.
Após analisarmos todos os resultados obtidos (Capítulo 2-6) seleccionámos, de entre as 25 árvores em estudo, as melhores 12 (2009) das quais foram utilizados ramos do ano para estudar a sua resistência à infecção com P.cinnamomi e C.parasitica (Capítulo 7). Após todos estes estudos, e ainda em 2009, seleccionámos as 9 melhores e enxertámo-las, em Ca90 (híbrido resistente: C. crenata C. sativa). Colocámos todas as plantas enxertadas nas mesmas condições edafoclimáticas, com o objectivo de efectuarmos uma caracterização fotossintética de cada clone (Capítulo 8). Em relação às trocas gasosas, obtivemos diferenças significativas quer no valor
da taxa fotossintética quer na temperatura óptima para a respectiva taxa, varia entre 3,8 µmol CO2m-2s-1 a 32,5ºC e 5,8 µmol CO2m-2s-1 a 22,0ºC para os
clones 136 (Murça) e 57 (Valpaços), respectivamente. Estes valores de temperatura estão em concordância com os valores de ºD obtidos nas árvores mãe, demonstrando alguma memória térmica por parte dos diferentes clones. Associando a termo-sensibilidade ao stresse hídrico obtivemos alterações nos teores de metabolitos nas folhas. Uma estratégia adoptada pelos clones com maior stresse foi o aumento de açúcares solúveis e redução na concentração de amido. Contrariamente, os clones mais bem adaptados mostraram um aumento quer no teor dos açúcares solúveis quer de amido, provavelmente devido à boa actividade fotossintética. Os pigmentos fotossintéticos também reflectiram a resistência/sensibilidade das plantas enxertadas bem como o teor em compostos fenólicos. A principal resposta ao stresse observado, em plantas sensíveis, foi o aumento do conteúdo em fenóis totais. A composição de ácidos gordos indicou que o baixo índice de insaturação encontrado nos clones 24 e 67 (Chaves e Alfândega da Fé) permitiram uma melhor termo-tolerância em contraste com o elevado nível encontrado nos clones 130, 136, (Bragança e Murça, respectivamente).
No estudo do Capítulo 9, o porta-enxerto escolhido pela sua conhecida resistência à “Doença da Tinta” provocada pela Phytophthora cinnamomi, foi o Ca90. Para melhor entender os mecanismos envolvidos nesta resistência efectuámos estudos fisiológicos e bioquímicos antes da inoculação e 9 e 16 dias pós-inoculação de plantas de Judia e Ca90. Todas as plantas de Ca90 sobreviveram ao oomiceta, enquanto 87,5% das plantas de Judia morreram 30 dias após a inoculação. Findos 16 dias, as plantas da variedade Judia apresentaram uma redução de 92,9% no seu potencial hídrico foliar
contrastando com os 35,8% das plantas de Ca90. Esta diminuição na Judia causou um decréscimo no teor de clorofila (23,2%) e um aumento na chla/b
(3,1-3,5). Igualmente, ocorreu uma quebra de 58,3% na taxa de fotossíntese das plantas de Judia. O aumento significativo no teor de açúcares solúveis e fenóis (50%) nos clones de Ca90 parece ser uma forma de resposta à invasão, resultando num aumento de resistência.
Para finalizar o Capítulo 10 refere-se às conclusões e discussões gerais onde é, também referida a importância deste estudo e quais as propostas de estudos futuros.
Palavras-Chave: Castanea sativa, Graus-dia, Calibre, Alteração foliar, Bioquímica básica, Compostos fenólicos, Actividade antioxidante, Selecção clonal, Phytophthora cinnamomi, Cryphonectria parasítica.
ABSTRACT
The chestnut is a multipurpose species whose fruits were for century’s part of the diet of many people who lived in mountainous regions where it was produced as well in the South of Europe. This trend has evolved over time and staple food of rural populations, the chestnut has also been consumed by urban populations using traditional recipes or haute cuisine. Finally, nowadays, the knowledge about its nutritional value, also turned it in a dietetic food (recommended for people with diabetic and celiac problems), leading to be known as an “unNut”.
Among the 13 species of the genus Castanea, the European one is Castanea sativa Mill. Unfortunately, climatic changes, particularly temperature and water stress, seem to limit its growth in many European regions.
The aim of this work was to contribute to better understand how different climatic conditions will influence the future development and consequently productivity of this specie in Trás-os-Montes region, especially the Judia variety.
With the collaboration of the farmers and producer’s associations from Trás-os-Montes region, trees from Judia variety (50 80 fruits / kg) were selected in the main chestnut localities. Then, ecotypes from seven localities were studied: Alfândega da Fé, Bragança, Chaves, Macedo de Cavaleiros, Murça, Valpaços e Vinhais.
In the first year (2006), 130 trees were selected. A second selection was performed according to the calibre (total: all fruits and correct: only viable
fruits) (Chapter 2) comprising a total of 25 trees, representing the best ones (Alfândega da Fé: 4, Bragança: 4, Chaves: 4, Macedo de Cavaleiros: 3, Murça: 2, Valpaços: 4 e Vinhais: 4). The genetic distance between trees obtained by genotyping with a set of molecular markers ranged between 0 and 0.227, demonstrating the proximity among genotypes, proving that all of them belonged to the same variety.
The study continued in 2007, with the 25 selected trees. Comparatively to the others under study, this year was colder than 2006. The mean amount of degree-days (°D) in 2006 and 2007 were 2551ºD and 2155ºD (May-October), respectively. Nevertheless, in 2007, fruits were 38% bigger and contained more 50% starch than the fruits harvested in 2006. However, the crude fat content was 6.41% lower. In 2006, the highest fruits (62.5 fruits/kg) and the highest starch content (400.7g/kg DM) were yielded in the coldest locality (Valpaços: 2300°D). Contrarily, the fruits from the hottest locality (Murça: 2751ºD), had the worst calibre (157.1 fruits/kg). In 2007, the fruits with the best correct calibre were obtained in Macedo de Cavaleiros (2163ºD; 46.4 fruits/kg), Murça had again the worst calibre (66.0 fruits/kg). The work continued with the morphological, histological leaf and phenological studies of each ecotype which are presented in Chapter 3.
The overall results suggest that the annual amount of temperature is a decisive factor in what concerns its influence on chestnut production. The optimal temperature conditions ranged from 2100°D/2200ºD (between May and October), and the best locality depending of the annual conditions.
In order to better understand why Judia nuts are frequently polyspermic controlled pollinations were done in 2008 (Chapter 4). Pollination and
embryo development are strongly influenced by climatic conditions that vary among localities with different altitudes and among years, being the main cause of variability in quality and quantity of production. On the other hand, fertilization and embryonic development are important steps that influence yield and quality of chestnut fruit, including the percentage of polyspermy.
Controlled pollinations of Judia variety were done with pollen from Negral and Lada, two compatible varieties. By histological observations the development of ovules was observed in the middle of July containing a megaspore mother cell. Judia’s flowers pollinated with Lada had mature ovules five days before the Judia´s flowers pollinated with Negral variety. The influence of pollinators on polyspermy and on the formation of wrinkles is not yet conclusive, but the time of pollination seems to be essential for the fruit growth. With regard to the fruit size achieved by the controlled pollination, the tree whose pollinator was the Negral variety showed better calibre than the results obtained with the Lada variety (168.2 fruits/kg and 281 fruits/kg, respectively). The fruit shape also showed some variation, since variety Negral seems to produce fruits more elongated than the ones produced from variety Lada.
In 2008, (average value = 1959°D; being colder than the previous ones), the analysis of the influence of temperature in the metabolite synthesis and technological properties was also done (Chapter 5). The ºD varied from 1597°D to 2077°D. This study demonstrated the positive correlation between ºD and the water, protein and fatty acids content of nuts. Trees from the coldest localities (1597°D and 1971°D), developed nuts with higher levels of water, protein, soluble sugars and starch content and a clear prevalence of polyunsaturated fatty acids. A study of water loss in unpeeled nuts showed
significant differences between localities. The locality with the lowest ºD had a higher percentage of water loss than the locality with the highest degree-days value. This difference in values reflects higher or lower levels in economic losses, respectively.
The aim of the Chapter 6 was to study the influence of different temperature on the antioxidant activity presented by the nuts .The total phenolic content ranged between 9.6 mg/g gallic acid equivalents (GAE) in the hottest locality (Murça) and 19.4 mg/g of GAE (coldest ecotype, Valpaços). The gallic acid and ellagic acid were the predominant phenolic compounds, and nuts from the coldest locality had the highest values. The evaluation of antioxidant activity was done by different ways: capture of the radical ABTS acid (2,2 '-azinobis-(3-ethylbenzothiazoline-6 sulfonic acid)) and DPPH (2,2-diphenyl-2-picryl-hydrazyl), FRAP (ferric reduction antioxidant power) and inhibition of oxidative hemolysis of erythrocytes. The fruits from Valpaços (coldest locality), showed the best antioxidant properties. Thus, the climatic conditions seem to be a limiting factor for production of phenolics and consequently to the antioxidant activity of fruit.
The best 12 trees were selected (which will be the mother plants of each clone) from the 25 old ones (2009). Six year stems of each selected ecotype were used to study their resistance to infection and P.cinnamomi and C.parasitica (Chapter 7).
At the end the best 9 trees were selected and were then grafted (2009) in Ca90 (resistant hybrid: C. Crenata C. sativa) and putted all plants in the same place, in order to be in future germobank for Judia variety. These plants were now characterized with photosynthetic traits (Chapter 8). Significant
differences were obtained in the photosynthetic rate and in the optimal temperature for the respective rate (ranging from 3.8 µmol CO2m-2s-1 at
32.5ºC and 5.8 µmol CO2m-2s-1 at 22.0ºC for clone 136 (Murça) and 57
(Valpaços), respectively. These temperature values are in agreement with the values of ºD obtained from the mother trees suggesting that they conserve some thermal memory from different clones (long-term adaptability). Associating the thermo-sensitivity to water stress, we obtained changes in the metabolites contents of the leaves. The strategy adopted by the clones under highest stress was the increase of soluble sugars and the reduction of starch content. The photosynthetic pigments also reflected the resistance/sensitivity of the grafted plants as well as the content of phenolic compounds. High levels of total phenols detected in stressed plants, were considered the main response against abiotic stresses. In what concerns the fatty acid composition, the low unsaturation level (clones 24 and 67: Chaves and Alfândega da Fé) showed better thermo-tolerance in contrast to the high level found in clones 130, 136, (Bragança and Murça, respectively ).
In the last Chapter 9, Ca90 was chosen as the rootstock control for its known resistance to "the ink disease" caused by Phytophthora cinnamomi. To better understand the resistance mechanisms plants were inoculated as well as seedlings from C. sativa grafted with Judia variety. The pos-inoculation phase (9-16 days after inoculation) was followed by physiological and biochemical studies. All plants of Ca90 survived to the oomycete infection, while 87.5% of the Judia plantlets died after 30 days. After 16 days, the plantlets of Judia variety showed a reduction of 92.9% in their leaf water potential, contrasting with 35.8% of the Ca90 plants. This decrease in Judia plantlets was caused by a decrease in chlorophyll content (23.2%) and an
increase in chla/b (3.1 to 3.5). Also, there was a decrease of 58.3% in the photosynthesis rate of Judia variety plantlets. The significant increase in soluble sugars and phenols (50%) in Ca90 clones seem to be a response to the invasion and consequently to increased resistance.
Finally Chapter 10 refers to the conclusions and general discussion which is also referred the importance of this study and which proposals for future studies.
Keywords: Castanea sativa, Degree-days, Calibre, Foliar changes, Basic biochemistry, Phenolic compounds, Antioxidant activities, Clonal selection, Phytophthora cinnamomi, Cryphonectria parasitica.
O
BJECTIVOS
A elaboração desta dissertação prende-se com alguns objectivos principais: i) selecção de ecotipos da variedade Judia conjugando melhores características tecnológicas com maior tolerância ao calor e às principais doenças do castanheiro (Castanea sativa Mill.) e ii) estudo, em plantas resistentes e sensíveis, da resposta fisiológica e bioquímica pós-inoculação com Phytophthora cinnamomi Rands.
No entanto para a elaboração dos diferentes trabalhos científicos, foram definidos objectivos mais específicos:
1. Caracterização de frutos da variedade Judia provenientes de sete diferentes ecotipos (diferentes condições climáticas) durante 2006 e 2007 com o intuito de encontrar as condições óptimas de graus-dia (somatório da temperatura) que possibilitem uma melhor qualidade e calibre do fruto (Capítulo 2).
2. Caracterização (somatório da temperatura, ºD) morfológica e histológica de folhas da variedade Judia provenientes de sete ecotipos diferentes da região de Trás-os-Montes (Capítulo 3).
3. Influência da temperatura (ºD) nas diferentes fases fenológicas (2007-Capítulo 3), estudo da biologia floral da variedade Judia e do desenvolvimento do óvulo, fertilização e embriogénese (2008) (Capítulo 4).
4. Avaliação da influência das condições climáticas (essencialmente ºD) na composição química e em certas propriedades morfológicas e
tecnológicas de frutos da variedade Judia em sete ecotipos diferentes (2008) (Capítulo 5).
5. Influência das condições climáticas na actividade antioxidante de frutos provenientes de sete ecotipos diferentes na região de Trás-os-Montes (2008) (Capítulo 6).
6. Avaliação do nível de tolerância, dos ecotipos de Judia seleccionados, à infecção com Phytophthora cinnamomi (doença da tinta) e
Cryphonectria parasitica (cancro) (Capítulo 7).
7. Selecção clonal de plantas da variedade Judia, foto e termo-resistentes, para posterior produção (2009) (Capítulo 8).
8. Monitorização fisiológica e bioquímica de plantas da variedade Judia e de Ca90 inoculadas com Phytophthora cinnamomi Rands para melhor entender os mecanismos de resistência à “doença da Tinta” (2008) (Capítulo 9).
LISTA DE FIGURAS
Figura 1.1 - Distribuição de C. sativa na Europa. 5
Figura 1.2 - DOP existentes na região de Trás-os-Montes e Alto Douro. 6
Figura 1.3 - Folhas típicas de Castanea sativa Mill.. 10
Figura 1.4 - Flores femininas (esquerda) e amentilhos ou flores masculinas (direita). 11
Figura 1.5 - Invólucro protector do fruto (esquerda) e deiscência de um ouriço com
três frutos (direita).
12
Figura 1.6 - Evolução da área de castanheiro e respectiva produção e produtividade
entre 1989 e 2009.
20
Figura 1.7 - Aspecto de um souto localizado em Rio Bom (Valpaços) com elevado
grau de ataque de cancro cortical (C. parasitica) visível pela deformação das copas dos castanheiros.
21
Figura 1.8 – Curvas da temperatura média anual, da época de polinização
(Junho-Julho) e maturação do fruto (Agosto-Outubro) de Portugal continental.
23
Figura 1.9 – Curvas da precipitação média durante a época de polinização
(Junho-Julho) e maturação do fruto (Agosto-Outubro) de Portugal continental.
24
Figura 2.1 - Influence of total amount of precipitation occurred between May and
September in the total fruit size.
44
Figura 2.2 - Effect of temperature amount (MayOctober) displayed as degree-day
(ºD) in the total and corrected fruit size. Correlation was done with all local data from 2006 and 2007.
45
Figura 2.3 - Influences of the temperature amount (MayOctober) displayed as
degree-day (ºD) in the starch content.
49
Figura 2.4 - Dendrogram (A) built according the morphological and chemistry data
and dendrogram showing the genetic similarity among different ecotypes (B).
52
Figura 2.5 - Variability of the localities under study, in both years using the first two
principal components based on fruit biometry and chemical analysis data.
54
Figura 3.1 – Influence of temperature sum (ºD) in lamina thickness (A) and stomata
density (B) in 2007, during the vegetative cycle (MayOctober).
Figura 3.2 – Influence of temperature sum (ºD) in leaf area (A). Relationship
between leaf area and corrected fruit size in 2006 (white lozenges) and 2007 (black lozenges) (B).
78
Figura 3.3 - Dendrogram showing the genetic similarity among different ecotypes
(A) and dendrogram (B) built according the 6 principal components extracted from leaf morpho (Table 3.2) and histological (Table 3.4) parameters and fruit biometric parameters (Table 3.5).
82
Figura 4.1 - Mitotic activity (anaphase) in the buds on April 17th, 2008 (A) and
pistillate inflorescences at blooming showing stamen remnants, collected in the 4th week of june, 2008 (B).
101
Figura 4.2 - Phenogram with the dates of blooming of the 3 varietiess and the dates
of pollination (X).
102
Figura 4.3 - Ovule development from megaspore mother cell stage to fertilization:
developing ovules are located in the upper part of the ovary axis,; collected July 14th (A – 200x, tree A- Lada); longitudinal section of ovule showing the megaspore stage, collected July 17th (B – 800x – tree C-Lada); longitudinal section of mature ovule showing two integuments and a nucellus with a small embryo sac; collected July 24th (C – 200x, tree C-Negral); zygote, collected August 7th (D – 800x, tree C-Negral).
104
Figura 5.1 – Principal component loadings plot (A) and scores plot (B) for the two
first principal components for all parameters under study.
130
Figura 6.1 – Example chromatograms of phenolic compounds of chestnut fruit
extract: 1 = Gallic acid; 2 = Ellagic acid.
151
Figura 6.2 – Antioxidant activity of Castanea sativa (var. Judia) from different
ecotypes: scavenging activity on ABTS.+ radicals (RSAABTS) (%, A),
scavenging activity on DPPH• radicals (RSADPPH) (%, B), FRAP value
(C) and hemolysis inhibition (%, D). Each value is expressed as mean ± standard error (n = 3 per tree).
155
Figura 7.1 – Evolution of the necrosis caused by P. cinnamomi in three studied
clones (75, 83 - resistant and 136 - susceptible).
Figura 7.2 – Evolution of the necrosis in the clone Judia 67, seven days after
infection with P. cinnamomi (left) and C. parasitica (right).
169 171
Fig.ura 7.3 – Geographical position of the ecotypes under study and respectively
necrosis length provoked by C.parasitica infection.
Figura 7.4 – Geographical position of the ecotypes under study and respectively
necrosis length provoked by Pcinnamomi infection.
172 172
Figura 8.1 - Relationship between optimal temperature (T100) and radiation
(PPFD100) with origin’s mean of degree-days (ºD).
185
Figura 8.2 - Correlation between maximal photosynthesis (A100T) and stomatal
conductance (gsT) (A) and transpiration (ET) (B).
189
Figura 8.3 – Lipid composition of leaves for different Judia’s grafted clones: (A)
percentage of each fatty acid species; (B) analysis of saturated fatty acids (SFA), unsaturated fatty acids (UFA) and lipid unstauration index (UI).
200
Figura 8.4 – Correlation between photosynthetic photon flux density for maximal
photosynthesis (PPFD100) and unsaturated index (UI).
201
Figura 8.5 – Principal component loadings plot (A) and scores plot (B) for the two
first principal components for all parameters under study.
203
Figura 9.1 Infected shoots of chestnut plantlets, showing the typical chlorosis
developed 16 days after inoculation with Phytophthora cinnamomi oomycete in the sensitive genotype (Judia, A) and resistant genotype (Ca90, B).
221
Figura 9.2 Newly developed shoots on the base of the stem before death of the
sensitive plants (Judia) upon inoculation with Phytophthora
cinnamomi oomycete.
222
Figura 9.3 – Results of the first two principal component analysis projected on the
space for all parameters under study (A) and variability of the control and inoculated plants of the two clones under study using the same principal components (B). In panel B, JCI9 and JCI16 stands for Judia control on day 9 and 16, respectively, and JI9 and JI16 for Judia infected on day 9 and 16, respectively. In same way, Ca90C9 and Ca90C16 stands for Ca90 control on day 9 and 16, respectively, and Ca90I9 and Ca90I16 for Ca90 infected on day 9 and 16, respectively.
232
Figura 10.1 – Variabilidade genética existente em algumas árvores em estudo de
Figura 10.2 – Análise dos valores médios dos calibres e bioquímica básica nos três
anos em estudo.
252
Figura 10.3 – Clones enxertados em Ca90 (Setembro 2009). A enxertia foi feita por
borbulha durante o mês de Junho 2009. Este trabalho decorreu em Vila Pouca de Aguiar em terrenos dos Viveiros Grupo ServiRuri.
254
Figura 10.4 – Terreno destinado à instalação do germobanco durante a sua
preparação em Janeiro 2010 (esquerda) e planta no local definitivo (Julho 2010, direita). O germobanco localiza-se em Torgueda (840 m altitude), em terreno com 3 ha, cedido por contrato de comodato entre a UTAD e a Junta de Freguesia.
254
LISTA DE TABELAS
Tabela 1.1 - Países com maior produção de castanha na Europa e na Ásia. 3
Tabela 2.1 - Edaphoclimatic characterization of seven different locals. There are
displayed the geographic coordinates, orchard altitude, climatic parameters (A) and soil physical and chemistry properties (0-50 cm) of each provenience (B).
36
Tabela 2.2 - Effect of year, locality and respective interaction on chestnut fruit
biometry.
42
Tabela 2.3 – Effect of locality, year and respective interaction on basic chemical
composition (g/kg DW) of the chestnut fruits.
48
Tabela 3.1 – Edaphoclimatic characterization of seven different locals. There are
displayed the geographic coordinates, orchard altitude, mean climatic parameters, degree-day (ºD) (A) and soil physical and chemistry properties (050 cm) of each provenience (B).
72
Tabela 3.2 – Leaf morphological characterization (A) ((leaf area (A), length (L),
width (W), shape index (SI=L/W) and shape factor (f)) and external leaves morphology (B) ((distance to first tooth (DT), number of teeth (NT), length petiole (LP), fresh weight (FW), dry weight (DW) and leaf mass per unit area (LMA)) of chestnut tree from seven areas of Trás-os-Montes.
73
Tabela 3.3 – Variation of 2007’s values compared to 2006. 74
Tabela 3.4 – Leaf tissue thickness (total lamina (Lth), upper epidermis (UE),
palisade layer (Pl), spongy layer (Sl), palisade /spongy (Pl/Sl), lower epidermis (LE)), ostiole (O) and stomata density (Sd). Data were collected (n=15 per tree) in 2007 in seven distinct places from Trás-os-Montes region.
75
Tabela 3.5 – Study of the fruit size of the samples from seven locals with different
edaphoclimatic conditions (corrected fruit/kg (CF), total fruit/kg (TF) and ratio corrected/total (CF/TF)).
77
Tabela 3.6 – The six principal components (PC) which explain 100% of cumulative
variance determined with all the parameters showed in Tables 3.2, 3.4 and 3.5 with corresponding the first five absolute values of high-impact variables.
79
Tabela 3.7 – Dating of phenological phases. Data were collected during 2007, and
include bud break, female flowering phase, male pollination, length of catkins (n=6 per tree) and beginning of fruit fall.
80
Tabela 3.8 – Genetic distance, below the imaginary line, and genetic identity above
the imaginary line.
Tabela 4.1 - Climatic characterization of the sites of cultivation of ‘Judia’ in 2007
and 2008. There are displayed the geostrategic coordinates, orchard altitude and mean climatic parameters: average temperature and degree-days.
98
Tabela 4.2 – Phenological phases of ‘Judia’ observed in 2007 in plants grown in sites
with different climatic characteristics (see Table 4.1).
99
Tabela 5.1 – Climatic, geographic (A) and edaphic (B, 0 - 50 cm) characterization of
different localities under study.
112
Tabela 5.2 – Proximate composition and free sugars (g kg-1 DM) of different locals
under study.
119
Tabela 5.3 – Mineral content (g kg-1 DM) of Judia variety located in seven localities. 122
Tabela 5.4 – Total lipids (g kg-1 DM), total fatty acids and ƩSFA, ƩUSFA (ƩMUFA
and ƩPUFA) composition (%) of the seven locals under study.
124
Tabela 5.5 – Morphological fruits parameters of Judia variety evaluated in seven
localities from Trás-os-Montes region
127
Tabela 6.1 – Climatic characterization of different localities under study. There are
presented the geographic coordinates, orchard altitude and climatic parameters: average temperature, degree-days and precipitation from MayOctober (A), and soil physical and chemical properties (0 - 50 cm) of each provenience (B).
149
Tabela 6.2 – Contents of phenolics and flavonoids (mg/g) in the extracts of
chestnut from different ecotypes of the Trás-os-Montes region.
152
Tabela 6.3 – Correlations established between total phenolics and flavonoids with
antioxidant activity EC50 values.
152
Tabela 6.4 – EC50 values obtained in the antioxidant assays for chestnut extracts
from different ecotypes.
158
Tabela 6.5 – Correlations established among different antioxidant activity EC50
values.
158
Tabela 7.1 – Sensibility (length of necrosis) of different clones, under study, to
P.cinnamomi and C. parasitica.
170
Tabela 8.1 – Values of degree-day of origin’s localities between May-October,
T100T, ET, gsT and WUET formaximal photosynthesis net (A100T) obtained
by the temperature curve andPPFD100, EPPFD, gsPPFD and WUEPPFD for
maximal photosynthesis net (A100PPFD) obtained by the radiation curve of
different grafted clones of Judia variety.
Tabela 8.2 – Chlorophyll fluorescence parameters of different grafted clones of Judia
variety.
191
Tabela 8.3 – Leaf area, specific leaf area (SLA), density of the leaf tissue (D),
relative water content (RWC), succulence (S) and water saturation deficit (WSD) of different clones of Judia variety (n = 6).
194
Tabela 8.4 – Photosynthetic pigments concentration per dry mass (mg g-1) and per
unit leaf area (mg dm-2) of different grafted clones of Judia variety.
196
Tabela 8.5 – Leaf metabolites concentration per unit leaf area (mg dm-2) of different
grafted clones of Judia variety.
197
Tabela 9.1 - Weather characterization (average, minimum and maximum air
temperature, relative humidity, precipitation and soil temperature) of the days and the periods under study.
220
Tabela 9.2 – Infected area and the number of died plants of chestnut 9, 16 and 30
days after inoculation with Phytophthora cinnamomi.
221
Tabela 9.3 –Changes in physiological parameters (A, net CO2 assimilation; gs,
stomatal conductance; E, transpiration; A/gs, intrinsic water use
efficiency; WUE, water use efficiency; Ci/Ca, ratio internal CO2
concentration to ambient CO2; ψw, leaf potential) of chestnut Judia and
Ca90 plants upon inoculation with Phytophthora cinnamomi.
224
Tabela 9.4 – Changes in chlorophyll fluorescence parameters (Fo, minimum
fluorescence yield; Fm, maximum fluorescence yield; Fv, variable
fluorescence yield; Fv/Fm, photochemical efficiency of PSII of
dark-adapted leaves) after inoculation with Phytophthora cinnamomi to Judia and Ca90 plantlets
227
Tabela 9.5 – Analysis of photosynthetic pigments (Chla+b, total chlorophyll; Chla/b,
ratio of Chla to Chlb; Car, total carotenoids; Chl/Car, ratio of
chlorophylls to carotenoids) and SPAD-readings on infected and non-infected (control) chestnut plants of Ca90 and Judia clones after inoculation with Phytophthora cinnamomi.
228
Tabela 9.6 – Leaf metabolites concentration (SP, total soluble protein; SS, total
soluble sugars; St, starch; TP, total phenols) and antioxidant activity (AA, antioxidant activity) of the clones under study.
ÍNDICE GERAL
AGRADECIMENTOS V RESUMO xix ABSTRACT xvii OBJECTIVOS xxiii LISTA DE FIGURAS xxvLISTA DE TABELAS xxix
CAPÍTULO 1
Introdução Geral
1.1 Introdução 3
1.2 Classificação sistemática do género Castanea 7 1.3 Origem do castanheiro europeu (Castanea sativa Mill.) 8 1.4 Aspectos bioecológicos de Castanea sativa Mill. 9
1.5 Influência da temperatura 13
1.5.1 Influência da temperatura no ciclo vegetativo 14 1.5.2 Influência da temperatura na polinização 15 1.5.3 Influência da temperatura na maturação do fruto 16 1.5.4 Influência da temperatura na incidência da doença da tinta 18
1.6 Referências bibliográficas 24
CAPÍTULO 2
Study of Morphological and Chemical Diversity in Chestnut Trees (var. „Judia‟) as a Function of Temperature Sum
2.1 Introdução
33
2.2 Material and Methods 34
2.2.1 Plant material 34 2.2.2.Edaphoclimatic conditions 35 2.2.3 Fruit biometry 37 2.2.4 Chemical analysis 37 2.2.5 Molecular characterization 38 2.2.6 Data analysis 38
2.3 Results and Discussion 39
2.3.1 Edaphoclimatic conditions 39 2.3.2 Fruit biometry 40 2.3.3 Chemical analysis 46 2.3.4 Multivariate analysis 50 2.4 Conclusions 55 2.5 References 56
CAPÍTULO 3
Study of Morphological and Phenological Diversity in Chestnut Trees („Judia‟ variety) as a Function of Temperature Sum
3.1 Introduction
63
3.2 Material and Methods 65
3.2.1 Plant material and growth conditions 65
3.2.2 Leaf morphological characterization 66
3.2.3 Leaf histology 67 3.2.4 Fruit biometry 68 3.2.5 Phenology 68 3.2.6 Molecular characterization 69 3.2.7 Statistical analysis 69 3.3 Results 70 3.3.1 Edaphoclimatic conditions 70
3.3.2 Leaf morphological characterization 70
3.3.3 Leaf histology 74 3.3.4 Fruit biometry 76 3.3.5 Phenology 79 3.3.6 Molecular characterization 80 3.4 Discussion 82 3.5 References 89
CAPÍTULO 4
Phenology and Reproductive Biology in variety “Judia” (Castanea sativa Mill.)
4.1 Introduction 97
4.2 Material and Methods 98
4.3 Results and Discussion 100
4.4 References 105
CAPÍTULO 5
Chemical composition and technological properties of chestnut fruits (var. “Judia”) as a function of edaphoclimatic conditions
5.1 Introduction 109
5.2 Material and Methods 110
5.2.1 Plant material and growth conditions 110
5.2.2 Chemicals 110
5.2.3 Edaphoclimatic conditions 111
5.2.4 Proximate composition analysis and free sugars 112 5.2.5 Determination of total lipids and fatty acids composition 113
5.2.6 Mineral composition 113
5.2.7 Morphological and technological properties of fruits 114
5.3 Results and Discussion
116
5.3.1 Edaphoclimatic conditions 116
5.3.2 Proximate composition analysis and free sugars 116
5.3.3 Mineral composition 120
5.3.4 Total lipids and fatty acids composition 123 5.3.5 Morphological and technological properties of fruits 125 5.3.6 Classification by multivariate analysis 128
5.4 References 131
CAPÍTULO 6
Antioxidant activities of chestnut nut of Castanea sativa Mill. (variety Judia) from different ecotypes
6.1 Introduction 139
6.2 Material and Methods 141
6.2.1 Chemicals 141
6.2.2 Samples and samples preparation 141
6.2.3 Edaphoclimatic conditions 142
6.2.4 Determination of phenolic and flavonoid content 143 6.2.5 HPLC analysis of phenolic and flavonoid compounds 144
6.2.6 ABTS• +radical-scavenging activity 144
6.2.7 DPPH radical-scavenging activity 145
6.2.8 Ferric reducing activity (FRAP assay) 146 6.2.9 Inhibition of erythrocyte hemolysis mediated by peroxyl
free radicals
147
6.2.10 Statistical analysis 148
6.3 Results and Discussion 148
6.3.1 Edaphoclimatic conditions 148
6.3.2 Antioxidants contents 150
6.3.3 Antioxidant properties 153
6.4 References 160
CAPÍTULO 7
Study of Judia ecotypes susceptibility to ink (P. cinnamomi) and blight (C. parasitica) diseases
7.1 Introduction 167
7.2 Material and Methods 168
7.3 Results and Discussion 168
CAPÍTULO 8
Influence of long-term adaptations in the photosynthetic productivity of Castanea sativa (var. Judia)
8.1 Introduction 177
8.2 Material and Methods 180
8.2.1 Plant material and growth conditions 180
8.2.2 Gas exchange measurements, leaf potential and chlorophyll
fluorescence 180
8.2.3 Sclerophylly and leaf moisture parameters 182 8.2.4. Photosynthetic pigments and foliar metabolic assays 182
8.2.5 Fatty acids 183
8.2.6 Statistics 183
8.3 Results and Discussion 184
8.3.1 Gas exchange measurements and chlorophyll fluorescence 184 8.3.2 Sclerophylly and leaf moisture parameters 192 8.3.3 Photosynthetic pigments and foliar metabolic assays 195
8.3.4 Fatty acids 198
8.3.5 Multivariate analysis 202
CAPÍTULO 9
Comparison of the physiological and biochemical changes between the resistant and the sensitive chestnut (Castanea) plantlets after inoculation with Phytophthora cinnamomi
9.1 Introduction 213
9.2 Material and Methods 216
9.2.1 Plant material 216
9.2.2 Pathogen inoculation and injury area measurement 217 9.2.3 Gas exchange measurements, SPAD-readings and
fluorescence 217
9.2.4 Water potential 218
9.2.5 Photosynthetic pigments and metabolites essays 218
9.2.6 Statistical analysis 219
9.3 Results 220
9.3.1 Weather conditions 220
9.3.2 Morphologic changes 220
9.3.3 Gas exchange, leaf water potential and fluorescence 222 9.3.4 Photosynthetic pigments and SPAD-readings 226
9.3.5 Leaf metabolites 229
9.3.6 Multivariate analysis 230
9.4 Discussion 233
9.4.1 Gas exchange, leaf water potential and fluorescence 233
9.4.2 Photosynthetic pigments 237
9.4 References 241
CAPÍTULO 10
Conclusões Gerais
10.1 Conclusões gerais e discussão 249
10.2 A Importância deste estudo para o melhoramento da espécie
Castanea sativa 256 10.3 Referências 257 ANEXOS Fichas Clonais Judia 24 261 Judia 57 263 Judia 58 265 Judia 67 267 Judia 75 269 Judia 83 271 Judia 100 273 Judia 130 275 Judia 136 277
1.1 Introdução
O castanheiro (Castanea sativa Mill.) é considerado uma planta multiusos que se desenvolve principalmente no Hemisfério Norte dada a sua aptidão para produção de fruto e madeira. A produção de fruto está mais localizada em alguns países asiáticos e na Europa, onde Portugal assume um destaque importante (Tabela 1.1). É possível observar grandes oscilações na década 1998-2008, tanto na Europa como na Ásia. Assim, em Portugal a produção de castanha teve um incremento até 2003, com excepção de 2001, verificando-se uma forte redução nos últimos anos (excepção para 2006). Em Itália (maior produtor de castanha da Europa), nestes 10 anos, a produção foi de cerca de 50 mil toneladas, observando-se um aumento gradual a partir de 2004. A China, o maior produtor mundial, tem tendência semelhante, mas a um ritmo acentuado. Contrariamente, esteve a produção na Coreia do Norte e no Japão. Em Portugal, a produção tem-se mantido estável, dependendo das variações anuais que se observam das consequências climáticas do ano.
Tabela 1.1 – Países com maior produção de castanha na Europa e na Ásia (Fonte: FAO –
FAOSTAT Statistics Division, 2009 e 2010 [1]).
Ano
Espanha França Grécia Itália Portugal China Japão Coreia do Norte
1998 11852 11411 12820 78425 29314 450000 26200 109956 1999 10580 12563 15733 52158 30969 534631 30000 95768 2000 9230 13224 15303 50000 33317 598185 26700 92844 2001 9510 13032 14935 50000 26118 599077 29000 94130 2002 9362 11223 15200 50000 31385 701684 30100 72405 2003 16821 10118 16800 50000 33267 797168 25100 60017 2004 9510 12431 18712 50000 31051 922735 24000 71795 2005 8629 8144 20946 52000 22327 1031857 21800 76447 2006 10140 9670 17442 53000 30900 850000 23100 82450 2007 15000 8284 10200 55000 22000 925000 22100 77524 2008 15000 6258 9800 55000 22000 925000 22100 80000 Produção de Castanha (t)
O castanheiro distribui-se por regiões que apresentam uma pluviosidade anual entre 600 mm e 1000 mm. Tradicionalmente é aceite que deve ocorrer alguma precipitação no período Agosto/Setembro para que o crescimento do fruto seja beneficiado. Contudo resultados apresentados por [2] demonstram que em soutos adultos, mais importante que esta precipitação que possa ocorrer no período de Verão, é que durante o período de Inverno ocorra precipitação em quantidade suficiente para repor os níveis de água das camadas mais profundas do solo, de onde o sistema radicular das árvores absorve a água necessária ao crescimento durante o Verão. Provavelmente o benefício que é visto nas chuvas de Verão poderá estar mais ligado ao arrefecimento que consequentemente ocorre, dado que temperaturas acima dos 30-32ºC podem induzir reduções de crescimento (através da taxa de fotossíntese) acima de 50% da taxa máxima conforme a variedade [3]. Adapta-se bem a locais cujo clima sofre influência marítima, com temperaturas médias anuais entre os 8ºC e 15ºC e temperaturas mensais acima dos 10ºC, pelo menos durante seis meses [4]. Suporta bem invernos rigorosos no entanto, as alterações climáticas nomeadamente a subida da temperatura parecem começar a condicionar significativamente o seu desenvolvimento [5]. Na Europa, a área de castanheiro para produção de fruto está limitada a sul pelas Canárias (27ºN) e Madeira (32,5ºN) e a norte a produção só é possível até aos 53ºN, correspondendo a uma linha imaginária que passa pelo sul de Inglaterra e norte da Alemanha. Portugal localiza-se precisamente no centro desta grande faixa de Continente Europeu, localizando-se as principais áreas de castanheiro nos terrenos montanhosos de interior, entre os 500 e os 1100 m altitude correspondendo a uma latitude variando entre os 38,5ºN e os 41,3ºN [3] (Fig. 1.1).
Fig. 1.1 – Distribuição de C. sativa na Europa (Fonte: EUFORGEN, 2009).
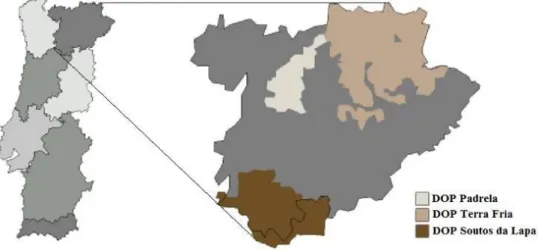
Em Portugal, cerca de 85% da área de souto localiza-se em Trás-os-Montes e Alto Douro, na sua parte mais montanhosa correspondente à Terra Fria, localizada sobretudo na parte norte da província. Aqui o clima tem mais semelhança com o clima de influência marítima acima referido como o mais favorável. No entanto, há algumas centenas de anos, antes da introdução das novas culturas, o castanheiro ocupou importantes áreas no Minho. Em Portugal, existem quatro áreas com Denominação de Origem Protegida (DOP) para a castanha: i) DOP da Castanha da Terra Fria; ii) DOP da Castanha dos Soutos da Lapa; iii) DOP da Castanha da Padrela; iv) DOP da Castanha do Marvão. Três destas DOP enquadram-se em Trás-os-Montes e Alto Douro (Fig. 1.2). Cerca de 70% a 80% da produção destina-se ao
mercado externo (em fresco e transformada), maioritariamente para países da União Europeia, em especial os da orla mediterrânica e Reino Unido e ainda para o continente americano (em especial o Brasil), bem como para o Japão.
Fig. 1.2 – DOP existentes na região de Trás-os-Montes e Alto Douro (Fonte: Borges et al.
[6]).
A área de produção da castanha diminuiu drasticamente na segunda metade do Séc. XX. Este decréscimo deveu-se em larga escala à doença da tinta provocada pelo oomiceta Phytophthora cinnamomi Rands e ocasionalmente Phytophthora cambivora Petri que ataca as raízes e colo das árvores [7] e mais recentemente pelo cancro do castanheiro originado pelo fungo Cryphonectria parasitica (Murrill) Barr [8]. Apesar dos inúmeros esforços para combater estas fraquezas na área da prevenção, muitos trabalhos de investigação foram dirigindo a sua atenção para a obtenção de plantas resistentes às referidas doenças como é o caso do COLUTAD [9] e Ca90 (comercialmente conhecido como Ferrosacri proveniente do INRA, França).
Percorrendo as várias áreas de Portugal existem imensas variedades [10-12] no entanto, em Trás-os-Montes, as mais conhecidas e vantajosas sob o ponto de vista frutícola e com melhor cotação a nível comercial são a Judia e a Longal [13]. A variabilidade genética destas variedades foi determinada primeiramente através de estudos isoenzimáticos [11, 14] e com marcadores moleculares tais como microssatélites[10] e RAPDs (Random Amplification of Polymorphic DNA) [15].
As origens mais importantes de diversidade de variedades de castanheiro Europeu, avaliadas com recurso a marcadores moleculares (microssatélites nucleares) localizam-se nas Astúrias, Galiza e na Península Ibérica Central (Trás os Montes, Salamanca, Cáceres e Ávila, apresentando a região de Trás-os-Montes o maior número de genótipos diferentes, indicando a importância desta região como centro de diversidade [16]. Estes autores explicam a diferenciação genética observada entre o Norte e Centro da Península Ibérica, como adaptações às condições climáticas, principalmente gradientes de temperatura e precipitação. Um estudo recente, realizado pelos mesmos autores, concluiu que existem 10 grupos principais de variedades em Portugal e Espanha, sendo a Longal a mais antiga e principal variedade, a partir da qual se originaram as variedades mais recentes, através de hibridação, mutação. (artigo em publicação, Rita Costa, comunicação pessoal).
1.2 Classificação sistemática do género Castanea
Pertencente a um dos nove géneros da Família Fagaceae e sub-família Castaneoideae, Castanea sativa é a espécie predominante em Portugal, bem
como no resto da Europa. Incluídos na mesma Família, Fagus e Quercus são, a par do Género Castanea, ecológica e economicamente importantes. Do Género Castanea fazem parte treze espécies entre as quais se destacam Castanea crenata Sieb. e Zucc. predominante no Japão, Castanea mollissima Bl. (China e Coreia), Castanea dendata Borkh. (América do Norte) e Castanea sativa Mill. predominante na Europa [17].
1.3 Origem do castanheiro europeu (Castanea sativa Mill.)
Estudos sobre a origem do castanheiro europeu (C. sativa) indicam que terá proveniência da Região do Cáucaso [18] há cerca de 90 milhões de anos, no Cretácio Médio, difundindo-se pela Europa com ajuda antropológica durante o período Cenozóico [8]. Por volta de 3000 a.C. o castanheiro europeu já estava bem presente na Grécia e no Sul de Itália. No milénio seguinte alcançou a Espanha e o sul de França [19]. O cultivo de C. sativa assume verdadeira expressão mais tarde, na Europa e nos países da margem sul do Mediterrâneo pela acção de difusão dos Romanos (Fig. 1.1).
Acabou por se estender por Portugal, Espanha, França, Itália, Eslovénia, Croácia, Grécia, Bulgária, Turquia, regiões junto ao Mar Cáspio e uma faixa estreita no norte de África que vai de Marrocos à Tunísia. Segundo Teixeira e Pais [20] em Portugal o castanheiro europeu está representado em depósitos da idade miocénica, há cerca de 15 milhões de anos nas proximidades de Canha. Estudos mais recentes referem a descoberta de vestígios de pólen fóssil de castanheiro na Serra da Estrela em formações do Paleolítico, com cerca de oito mil anos [21] o que permite afirmar que a ocupação do
castanheiro em Portugal possa ser anterior à ocupação da Península Ibérica pelos romanos, que no entanto deram um forte contributo para a sua dispersão pelo Império Romano e consequentemente pelos territórios que actualmente constituem Portugal Esta distribuição é descontínua e fruto de influências climáticas e antropogénicas [22].
Actualmente, fruto da introdução na Europa das espécies orientais, a fim de entrarem em programas de melhoramento do castanheiro, existem muitos exmplares dispersos, levando a que devido à polinização livre, muitas plantas resultantes de hibridações espontâneas estejam espalhadas por todo o lado.
1.4 Aspectos bioecológicos de
Castanea sativa Mill.
O castanheiro europeu é uma espécie caducifólia de grande porte (pode atingir 3035 m) e longevidade [23]. É constituído por folhas simples, caducas, pecioladas com o limbo oblongolanceolado, com recorte marginal crenato-serrado ou serrado, apresentando dentes cuspidado-aristados no prolongamento das nervuras secundárias, subcordado a subtruncado na base do limbo, com comprimento entre 10 a 25 cm e largura entre 5 a 8 cm (Fig. 1.3). De acordo com as variedades, a coloração das folhas varia, havendo sempre uma distinção entre a coloração (verde-escuro lustrosa) da página superior e a verde-claro da inferior. O pecíolo é amarelado ou vermelho com cerca de 2.5 cm de comprimento [24, 25].
Fig. 1.3 – Folhas típicas de Castanea sativa Mill.
Possui flores masculinas e femininas separadas mas na mesma planta (monóica) [26]. As flores estaminadas são amarelas na ântese e encontram-se reunidas em glomérulos de 5-6 flores (Fig. 1.4), dos quais emergem 10-20 estames com anteras ovóides, cor amarelo-esverdeada. As flores masculinas reúnem-se em amentilhos (Fig. 1.4), erectos ou sub-pendentes, com 15-30 cm de comprimento e 1 cm de diâmetro, que se inserem na parte terminal dos ramos do ano, ou próximo da base das folhas destes [25]. As inflorescências femininas surgem inseridas na base dos amentilhos androgínicos, menos numerosos, e são constituídas por 2-3 flores femininas, protegidas por um invólucro coriáceo protector, de brácteas verdes espinescentes (Fig. 1.5), que irá dar origem à cúpula [27]. O pólen é viscoso e a polinização pode ser entomófila (insectos) ou anemófila, em menor escala.
Fig. 1.4 – Flores femininas (esquerda) e amentilhos ou flores masculinas (direita).
O fruto é um pseudofruto, simples e seco. Além do embrião, cada semente tem também dois cotilédones, que constituem a parte comestível do fruto. A maturação anual dos frutos precisa, em média, de 75120 dias, desde a polinização à maturação ocorrendo deiscência a meados de Setembro, nas variedades mais precoces, e início de Novembro nas variedades mais serôdias [27]. Segundo Bergougnoux et al. [23] um ouriço possui 13 frutos (Fig. 1.5) mas, mais raramente, podem aparecer 78 frutos. O fruto é formado a partir do óvulo fertilizado contendo apenas uma semente fértil (monospérmico) ou se cada flor feminina tem vários óvulos poderão formar-se frutos polispérmicos. Assim, de acordo com o mesmo autor, no primeiro caso o fruto é designado de “marron” enquanto no segundo caso o fruto é designado de “castanha”.
Fig. 1.5 - Invólucro protector do fruto (esquerda) e deiscência de um ouriço com três frutos
(direita).
Como qualquer outra espécie frutícola, o castanheiro apresenta determinadas exigências edafoclimáticas que importa conhecer com o objectivo de o instalar em zonas edafoclimáticas adequadas e assim obter melhor rendimento. O castanheiro predomina em locais com diferentes tipos de clima: atlântico marítimo (invernos suaves e muito chuvosos), mediterrânico (verões quentes e secos) e continental (invernos rigorosos e verões quentes) [28]. A sua localização em Portugal sugere locais com características entre o continental e o mediterrânico com altitudes entre os 500 e os 1000m. Na Europa conhecem-se povoamentos para fruto a 1500m, como é o caso da Serra Nevada em Espanha e na Sicília [24, 29, 30] ou mesmo na Galiza onde podem ser encontrados castanheiros 1300m de altitude [31]. Relativamente à precipitação o castanheiro europeu necessita de 800 a 900 mm/ano, no entanto nos híbridos euro-japoneses esta necessidade aumenta para 12001300 mm/ano [32].
Em relação ao tipo de solo apropriado, algumas das limitações dos solos ao desenvolvimento de C. sativa são: i) reduzida espessura efectiva de solo, porque diminui o suporte radicular do castanheiro; ii) desequilíbrio textural (solos francos e franco-limosos), dá origem a uma estrutura com baixa estabilidade, o que propicia condições de arejamento pouco favoráveis ao castanheiro; iii) má drenagem particularmente em Invernos chuvosos. O excesso de água no solo e consequente falta de oxigénio têm sido associados à disseminação rápida da doença da tinta; iv) baixos teores de matéria orgânica (<20 g/kg), pois têm sido associados à maior incidência da tinta do castanheiro. O teor de matéria orgânica do solo além de promover uma maior estabilidade da estrutura e o arejamento do solo, deverá propiciar uma flora microbiana mais diversa; elevada acidez do solo e baixos teores de Ca e Mg de troca são limitantes ao desenvolvimento radicular do castanheiro e à nutrição das árvores [33, 34].
1.5 Influência da temperatura
O aquecimento global é cada vez mais um tema de debate e preocupação mundial. É esperado um aumento de 13ºC na temperatura média do ar no Hemisfério Norte [35]. Estas alterações climáticas provocam sérias limitações no crescimento das produções de castanha localizadas essencialmente nos locais mais quentes [36]. Uma vez que o castanheiro é uma espécie caducifólia (queda de folhas e paragem vegetativa no Inverno), poderá ser mais limitante, para o seu crescimento e desenvolvimento, o calor de verão que o frio do inverno. Em Portugal, os valores médios de temperatura máxima na região castanhícola, variam entre os 23-25ºC no
Minho Interior, 27ºC na Beira Alta e os 31ºC na Terra Fria Transmontana (maior produtor) e na zona de Marvão, onde se localiza um importante núcleo de castanheiro [3]. O número de dias quentes no ano, acima de 30ºC, constitui um indicador relevante do clima [37]. Na Terra Fria Transmontana, ocorrem, em média, quase 70 dias com temperaturas acima deste valor, enquanto na Beira Alta apenas ocorrem cerca de 30 dias. Esta limitação parece ser mais restritiva do que propriamente o stresse hídrico induzido por longos períodos de seca, nomeadamente quando nos referimos a árvores adultas com cerca de 3040 anos [38].
1.5.1 Influência da temperatura no ciclo vegetativo
O impacto das alterações climáticas na ecologia e fenologia de espécies vegetais já foi demonstrado [39]. Tal como em outras espécies, o ciclo vegetativo do castanheiro ocorre, normalmente, entre Março e Outubro. O inicio da actividade mitótica é a primeira fase deste ciclo e varia com as alterações de temperatura. Dinis et al., [40] refere num estudo, sobre a variedade Judia, que em locais cujos valores de somatório de temperatura (graus dia, ºD) são mais elevados, o inicio da actividade mitótica é mais precoce. Contrariamente, a deiscência do fruto é mais tardia em locais com valores mais baixos de ºD. Em concordância com estes resultados, os mesmos autores referem que em anos mais quentes a plena da floração feminina ocorre igualmente mais cedo que em anos mais frios. Assim, com o aumento da temperatura, a precocidade do ciclo vegetativo poderá ser prejudicado pelas geadas precoces de Outono que afectarão posteriormente a maturação do fruto e consequentemente a produção.
![Tabela 1.1 – Países com maior produção de castanha na Europa e na Ásia (Fonte: FAO – FAOSTAT Statistics Division, 2009 e 2010 [1])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15780520.1076932/39.748.104.652.682.887/tabela-países-produção-castanha-europa-faostat-statistics-division.webp)



![Fig. 1.6 – Evolução da área de castanheiro e respectiva produção e produtividade entre 1989 e 2009 (Fonte: INE, estatísticas agrícolas 2009 [70])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15780520.1076932/56.748.117.634.466.714/evolução-castanheiro-respectiva-produção-produtividade-fonte-estatísticas-agrícolas.webp)

![Fig. 1.8 – Curvas da temperatura média anual, da época de polinização (Junho-Julho) e maturação do fruto (Agosto-Outubro) de Portugal continental (Fonte: INM, 2009 [71])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15780520.1076932/59.748.121.622.463.770/curvas-temperatura-polinização-maturação-agosto-outubro-portugal-continental.webp)


