Parent selection for cocoa resistance to witches’‑broom
Texto
Imagem



Documentos relacionados
The chocolates from SR162 cultivar, which is resistant to “witches’ broom disease”, showed higher antioxidant activity as well as higher concentration of monomeric
The relative coefficient of variation presented median values for the number of plagiotropic branches and fresh fruit yield (0.81 and 0.92, respectively) showing that selection
which considers only the reaction to white mold, the gains for these traits (straw test and oxalic acid absorption) are greater when compared to those obtained from selection
The aim of this study was to obtain progenies of com- mon bean with carioca type grains resistant to anthracnose and to angular leaf spot arising from crosses between the
isolates (1051 and TVC), previously found to control witches’ broom of cocoa under field conditions (José Luiz Bezerra, personal communication), produced
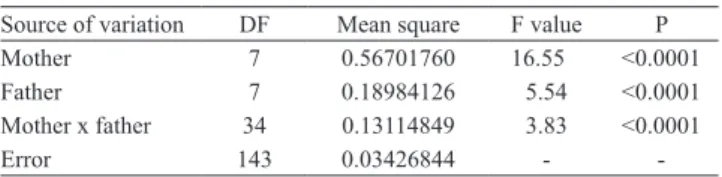
Heritability estimates and selective accuracy demonstrated a favorable situation to the selection of resistant progenies ,especially ‘STA-02’, ‘STA-04’, ‘STA-05’,
Considering two resistance classifications, and choosing as eligible for selection only clones at highly resistant/immune or very resistant classes according to Taylor (1967) and
