Cidália Raquel Moreira Borges
Study of sardine oil antioxidant
and
anti-inflammatory
properties for the development
of topical therapeutic and
cosmetic formulations
Master thesis
Master in Applied Biochemistry – Biomedicine
Work performed under the orientation of:
Prof. Dr. Andreia Gomes
Prof. Dr. Artur Cavaco-Paulo
Dr. Artur Ribeiro
January 2015
DECLARAÇÃO
Nome: Cidália Raquel Moreira Borges
Endreço electrónico: [email protected]
Número do Cartão de Cidadão: 13723651
Título da tese: Study of sardine oil antioxidant and anti-inflammatory properties for the development of topical therapeutic and cosmetic formulations
Orientadores: Professora Doutora Andreia Gomes
Professor Doutor Artur Cavaco-Paulo Doutor Artur Ribeiro
Ano de conclusão: 2015
Mestrado em Bioquímica Aplicada, Ramo de Biomedicina.
É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE COMPROMETE;
Universidade do Minho: 30 de Janeiro de 2015
Agradecimentos
Ao longo desta caminhada, foram muitas as pessoas que me ajudaram e apoiaram. A todas elas a expressão de um sincero agradecimento.
Começo primeiro pela Professora Doutora Andreia Gomes, orientadora deste projecto pela sua excelente orientação científica, disponibilidade,boa-disposição, acima de tudo pela oportunidade dada e confiança depositada ao longo deste projecto.
Ao Professor Doutor Artur Cavaco Paulo, co-orientador, pelo seu interesse, pelos ensinamentos,atenção e ajuda.
Ao Doutor Artur Ribeiro, co-orientador, pela inteira dedicação a este projecto, paciência, confiança, segurança, pelos valiosos ensinamentos e acima de tudo por me ter lançado nesta caminhada pelo mundo da ciência.
Ao longo desta experiência fizeste-me ver que: “A ciência têm as raízes amargas,
porém os frutos são doces." (Aristóteles). Obrigada por tudo!
À Doutora Marisa, pela amizade, pelo encorajamente constante, partilha de conhecimento e por toda a disponibilidade.
À Cristina, por todo apoio, partilha de conhecimento, ajuda e contribuição para o desenvolvimento deste trabalho.
Aos meus colegas do grupo de investigação, pela partilha, pelo conhecimento e boa-disposição.
Aos meus companheiros de laboratório, pelo bom ambiente laboratorial, pela partilha , ajuda e acima de tudo pela amizade e carinho.
Aos meus amigos, pela amizade, pela preocupação, pelo carinho, pela paciência em momentos mais complicados. Obrigada por me relembrarem que: “As dificuldades
devem ser usadas para crescer, não para desencorajar.”(William Channing)
À minha amiga Andreia, pela tua amizade, pela tua paciência nos momentos mais complicados, pelos teus conselhos e pelos grandes momentos partilhados.
Às minhas meninas, Kiki, Sophie e Maria, apesar de a vida nos levar a optar por caminhos diferentes, apesar da distância, a nossa amizade prevalece,as partilhas continuam e continuarão ao longo desta viagem e quiça um dia alugures encontramos-nos na mesma bancada de laboratório.
Nada disto seria possível sem o apoio incondicional dos meus pais e do meu irmão. Obrigada por acreditarem em mim , pela vossa presença constante e efectiva na minha caminhada. Obrigada por toda a compreensão e amor!
“Nas grandes batalhas da vida, o primeiro grande passo para a vitória é o desejo de vencer”
Study of sardine oil antioxidant and anti-inflammatory properties for the development of topical therapeutic and cosmetic formulations
Abstract
In Portugal sardine fishery is one of the oldest and the most important pelagic resource from Portuguese mainland, both economically and socially. The sardines have important nutritional characteristics, being an excellent source of vitamins, proteins, lipids and minerals with proven benefits for human health, being rich in ω-3-PUFAs, namely, EPA and DHA. Studies reported that there is a direct link between a diet enriched in ω-3-PUFAs and the prevention of many diseases such as cardiovascular disease, inflammatory conditions, mental disorders and prevention of various types of cancer.
The aim of this work was to characterize the antioxidant and anti-inflammatory role of sardine oil for the development of formulations for topical application.
To evaluate the antioxidant and anti-inflammatory effects of sardine oil in skin, human fibroblasts (BJ-5ta), human melanocytes (A375 and B), human keratinocytes (NCTC2544) and human monocytes (THP-1) were used.
The cytotoxicity studies showed that for higher concentrations of SO and SO+αT cell viability decreased. However the effect of SO+αT in keratinocytes was the opposite of melanocytes. Regarding PUFAs, concentrations higher than 0.025 mg/ml exhibited toxicity to both cell lines.
In proliferation assays, melanocytes suffer an inhibition of cell proliferation at concentrations above 4 mg/ml in SO. However, for the SO+αT cell proliferation increases with increasing concentration. In keratinocytes, the negative effect on cell proliferation when cells were incubated under same conditions was more marked. The cell proliferation profiles with PUFAs and PUFAs+αT were marked by inhibition in both cells lines, with more impact in melanocytes.
The lowest concentrations, 0.5 and 4 mg/ml, were tested to evaluate the protective role of sardine oil in the context of induced oxidative stress. Both concentrations were able to protect cells from damage with a more pronounced effect in keratinocytes. Moreover the incubation of cells with the sardine oil was able to induce expression of several cytoprotective enzymes and also the transcription factor Nuclear factor-erythroid-2-related factor 2 (Nrf2) – which plays a crucial role in the coordinated induction of many stress-responsive genes.
The lowest concentrations of sardine oil were tested on human fibroblasts and monocytes to evaluate their effect on inflammation and sardine oil showed to have anti-inflammatory potential as it can reduce production of pro-anti-inflammatory cytokine in cells stimulated with LPS.
The effect of sardine oil using three different assays (NaOH/DMSO, fluorescence and tyrosinase activity) on melanin synthesis was not clear. Only using the fluorescence based method for melanin assessment, the sardine oil showed to have some positive effect over melanin synthesis in human melanoma cells.
The results opened the opportunity to develop new therapeutic and cosmetic applications based on sardine-derived compounds. Their incorporation in topical creams may contribute to a better, more natural treatment of inflammation and in the prevention of skin aging.
Estudo das propriedades antioxidantes e anti-inflamatórias do óleo de sardinha para desenvolvimento de formulações tópicas com fins terapêuticos e cosméticos
Resumo
Em Portugal continental, a pesca da sardinha é um dos recursos pelágicos mais antigos e importantes, tanto a nível económico como social.
As sardinhas têm características nutricionais importantes, sendo uma excelente fonte de vitaminas, proteínas, lipídios e minerais com benefícios comprovados para a saúde humana. A sardinha é rica em ω-3-PUFAs, ou seja, EPA e DHA. Estudos relataram que há uma relação direta entre uma dieta enriquecida em ω-3-PUFAs e a prevenção de muitas doenças, tais como doenças cardiovasculares, doenças inflamatórias, distúrbios mentais e prevenção de vários tipos de cancro.
O objetivo principal deste trabalho foi caracterizar o papel antioxidante e anti- inflamatório do óleo de sardinha para futuro desenvolvimento de formulações com aplicação tópica.
Para avaliar os efeitos antioxidantes e anti-inflamatórios do óleo de sardinha na pele, foram usadas diversas linhas celulares de origem humana: fibroblastos (BJ-5ta), melanócitos (A375 e B) queratinócitos (NCTC2544) e monócitos (THP-1).
Os estudos de citotoxicidade mostraram que, para as concentrações maiores de SO e SO+αT a viabilidade celular diminui. No entanto, o efeito do SO+αT em queratinócitos foi contrário ao que aconteceu nos melanócitos. Em relação às PUFAs concentrações acima de 0,025 mg/ml induziram toxicidade em ambas as linhas celulares.
Em ensaios de proliferação, os melanócitos exibiram uma inibição da proliferação celular com óleo de sardinha em concentrações acima de 4mg/ml. Contudo, para SO+αT a proliferação aumentou com o aumento da concentração. Nos queratinócitos, o efeito negativo foi mais acentuado, quando as células foram incubadas sob as mesmas condições. Os perfis da proliferação celular em PUFAs e PUFAs+αT foram marcados pela inibição em ambas as linhas celulares, com mais impacto nos melanócitos.
As concentrações mais baixas, 0,5 e 4 mg/ml, foram testadas para avaliar o papel protetor do óleo de sardinha em situação de stress oxidativo. Estas duas concentrações foram capazes de proteger as células contra danos oxidativos, com um efeito mais evidente em queratinócitos. Além disso, a incubação de células com o óleo de sardinha
foi capaz de induzir a expressão de várias enzimas citoprotectoras e também o fator de transcrição Nrf2, que desempenha um papel crucial na indução coordenada de genes envolvidos em muitas respostas ao stress oxidativo.
As menores concentrações de óleo de sardinha foram testadas em fibroblastos e monócitos humanos em ensaios de inflamação. O óleo de sardinha mostrou ter um potencial efeito anti-inflamatório, uma vez que foi capaz de reduzir a inflamação em células estimuladas com LPS.
O efeito do óleo de sardinha estudado com três metodologias diferentes (NaOH/DMSO, fluorescência e atividade de tirosinase), sobre a síntese de melanina não foi claro. Somente usando uma abordagem baseada na fluorescência nos melanócitos humanos (B), o óleo de sardinha mostrou algum efeito positivo sobre a síntese da melanina.
Estes resultados criam a possibilidade de desenvolver novas aplicações terapêuticas e cosméticas à base de compostos derivados de sardinha. A sua incorporação em cremes tópicos podem contribuir para um melhor tratamento de inflamação e na prevenção do envelhecimento da pele.
Contents
Agradecimentos ... iii
Abstract ... v
Resumo ... vii
List of abbreviations ... xiii
List of tables ... xvii
List of figures... xix
I. INTRODUCTION ... 1
1.1 Sardine oil and human health ... 3
1.2 Omega-3 PolyUnsaturated Fatty Acids: the gold members of the fatty acids family. ... 4
1.3 Health benefits of ω-3 PUFAs ... 6
1.4 Role of ω-3 PUFAs in inflammation ... 8
1.5 Role of ω-3 PUFAs on Oxidative stress ... 10
1.6 Skin ... 14
1.7 Melanognesis ... 17
II. OBJECTIVES AND SCOPE OF THESIS ... 19
III. MATERIAL AND METHODS ... 23
3.1 Materials ... 25
3.1.1 Chemical Reagents ... 25
3.1.2 Buffers ... 27
3.1.3 Biological Materials ... 28
3.2 Cell culture maintenance... 29
3.3 Cell viability assessed by MTT assay ... 31
3.4 Cell proliferation assessed by Hoechst dye assay ... 32
3.5 Evaluation of oxidative stress by: ... 33
3.5.2 Catalase assay ... 34
3.5.3. Western Blotting assay ... 35
3.5.3.1 Analysis of protein levels profiles of HO-1, GCL, and NQO1 proteins 35 3.5.3.2 Analysis of protein levels profiles of Nrf2 transcription factor ... 36
3.6 Evaluation of the effects of anti-inflammatory assessed by: ... 37
3.6.1 Expression of IL-6 cytokine ... 37
3.6.2 Expression of TNF-α cytokine ... 38
3.6.2.1 Anti-inflammatory effect of sardine oil on previously stimulated cells . 38 3.6.2.2 Protective Anti-inflammatory effect of sardine oil... 38
3.7 Melanin quantifications assessed by: ... 39
3.7.1 NaOH/DMSO assay ... 39
3.7.2 Fluorescence assay ... 40
3.7.3 Tyrosinase assay ... 40
3.8 Statistical Analysis ... 40
IV. RESULTS AND DISCUSSION ... 41
4.1 Evaluation of SO and PUFAs cytotoxicity in melanocytes A375 and in keratinocytes NCTC2544 cell lines ... 43
4.1.1. Effect of SO and PUFAs in cell line A375 viability ... 43
4.1.2. Effect of SO and PUFAs in cell line NCTC2544 viability... 46
4.2 Effect of SO and PUFAs in cell proliferation of A375 melanocytes and NCTC2544 keratinocytes... 48
4.2.1 Effect of SO and PUFAS in proliferation of A375 cell line ... 48
4.2.2 Effect of SO and PUFAS in proliferation of NCTC2544 cell line ... 50
4.3 Protective effect of SO and PUFAs against induced oxidative stress ... 53
4.3.1 Evaluation of SO and PUFAs protective role by DCF- oxidative stress assay ... 53
4.3.2. Evaluation of SO and PUFAs protective role mediated by Catalase ... 56
4.4.1. Analysis of protein levels HO-1, GCL and NQO1 ... 60
4.4.2. Analysis of protein levels of Nrf2 ... 62
4.5 Anti -inflammatory effect of sardine oil ... 65
4.5.1 Evaluation of SO anti-inflammatory properties associated with levels of Interleukin- 6 in fibroblasts ... 65
4.5.2. Evaluation of SO anti-inflammatory properties by the levels of Tumor Necrosis Factor-α in monocytes ... 69
4.6 Study of the effect of sardine oil in melanogenesis ... 76
4.6.1 Effect of SO on melanogenesis in cell line A375 ... 76
4.6.2 Effect of SO on melanogenesis in cell line B ... 77
4.6.2.1 Evaluation of SO cytotoxicity in melanocytes cell line B ... 77
4.6.2.2 Effect of sardine oil on melanogenesis in cell line B ... 79
V. CONCLUSIONS AND FUTURE PERSPECTIVES ... 83
List of abbreviations
AA Arachodonic Acid
ALA α-Linolenic Acid
ARE Antioxidant Response Element
BSA Bovine Serum Albumin
COX Cyclo-OXygenase
DCFH2DA DiChlorodiHydroFlurescein DiAcetate
DMEM Dulbecco’s Modified Eagle’s Medium
DMSO DiMethylSulfOxide
DHA Docosahexanoic Acid
DHEA DocosaHexaenoylEthanolAmine
DHIA 5,6-DiHydroxyIndole Acid
DHICA 5,6-DiHydroxyIndole - 2- Carboxylic Acid
DTT DiThioThreitol
DPA DocosaPentanoic Acid
EDTA EthyleneDiamine Tetraacetic Acid
EPA EicosaPentanoic Acid
EPEA EicosaPentaenoylEthanolAmine
EpRE Electrophile Response Element
ERAD Endoplasmic Reticulum Associated Degradation factor
FBS Fetal Bovine Serum
GCL Glutamine Cysteine Ligase
GCLC Glutamine Cysteine Ligase Catalytic subunit
GCLM Glutamine Cysteine Ligase Modifier subunit
GPx Glutathione Peroxidase
GST Glutathione S-Transferase
15-HEPE 15-HydroxyEicosaPEntanoic acid
17-HODHE 17-HydrOxyDocosaHExanoic acid
HO-1 Heme-Oxygenase-1
ICAM InterCelular Adhesion Molecule-1
IL Interleukin
Keap-1 Kelch-like ECH associated protein-1
LA Linoleic Acid L-Dopa L-Dihydroxyphenylalanine LOX LipOXygenase LPS LiPopolySaccharides LTx LeuTrotrienes MTT MethylThiazolyldiphenylTetrazolium
NAEs N-Acyl Ethanolamines
NF-kB- Nuclear Factor kappa B
NQO-1 NADP(H) Quinone Oxireductase-1
Nrf2 Nuclear factor erythroid -2-related factor
PGs ProstaGlandins
PGE2 Prostaglandins E2
PPAR-α Peroxisome Proliferator Actived Receptor
PUFAs Polyunsaturated Fatty Acids
PUFAs+ αT Polyunsaturated Fatty Acids with α-Tocopherol
PVDF PolyVinlidene DiFluoride
ROS Reactive Oxygen Species
SCC Squamous Cell Carcinoma
SDS Sodium Dodecyl Sulfate
SO Sardine Oil
SO+ αT Sardine Oil with α-Tocopherol
SOD SuperOxide Dismutase
TEMED TetraMethylEthyleneDiamine
αT α-Tocopherol
TNF-α Tumor Necrosis Factor-α
TXs ThromboXanes
Tris Tris(hydroxymethyl)aminomethane
TYR TYRosinase
UV UltraViolet
List of tables
Table 1: EPA and DHA contents in some species of fishes. ... 4
Table 2: Reagents used in this work ... 25
Table 3: Antibodies employed and suppliers... 28
Table 4: Constitution of IL-6 and TNF-α kits ... 29
Table 5: Concentrations of SO, SO with Tocopherol, PUFAs, PUFAs with α-Tocopherol tested in cell lines A375, NCTC2544 and B ... 31
Table 6: Concentrations of SO, SO with Tocopherol, PUFAs, PUFAs with α-Tocopherol and α-α-Tocopherol tested in cell lines A375, NCTC2544 ... 34
List of figures
Figure 1. Sardines (Sardina pichardus) ... 3
Figure 2. The biosynthesis of ω-3 PUFAs ... 5
Figure 3. General scheme of the interacting mechanisms by which the ω-3 PUFAs might influence cell function ... 7
Figure 4. Scheme of anti-inflammatory mechanism of EPA as a competitive inhibitor of AA through the COX enzymes. EPA competitively inhibits PGE2 formation by COX-1 and COX-2 ... 9
Figure 5. Reactive oxygen species and their role in the development of human pathologies ... 11
Figure 6. The Keap-1 Nrf2 pathway ... 12
Figure 7. The NF-k B pathway ... 13
Figure 8. Schematic representation of the skin structure ... 15
Figure 9. Melanogenic pathway that occurs within melanosomes, leading to the production of eumelanin and/or pheomelanin ... 18
Figure 10. Representation of the reaction of enzymatic conversion of MTT to formazan ... 31
Figure 11. Cytotoxic effects of sardine oil (SO), sardine oil with α-Tocopherol (SO+αT) and α-Tocopherol (αT) in cell line A375 ... 44
Figure 12. Cytotoxicity of sardine oil (SO), sardine oil with α-Tocopherol (SO+αT) and α-Tocopherol (αT) in cell line NCTC2544. ... 47
Figure 13. Effects of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) in proliferation of A375 cells ... 49
Figure 14. Effects of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) on cell proliferation of cell line NCTC2544 ... 52
Figure 15. Protective effect of sardine oil (SO), sardine oil with α-Tocopherol (SO+αT) and α-Tocopherol (αT) measured by the DCF assay in cell lines A375 and NCTC2544 ... 54
Figure16. Effects of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) in the activity of catalase in cell lines BJ-5ta, A375 and NCTC2544 ... ..58 Figure 17. Effect of sardine oil (SO) on the levels of cytoprotective proteins in BJ-5ta, A375 and NCTC2544 cell lines... 61 Figure18. Effect of sardine oil (SO) on Nrf2/ARE signaling in BJ-5ta, A375 and NCTC2544 cell lines ... 64 Figure 19. Induction of inflammation of BJ-5ta cells by several concentrations of LPS at the end of 2, 8 and 24 hours of exposure ... 66 Figure 20. LPS induced inflammation of BJ-5ta cells along time ... 67 Figure 21. ELISA quantification of IL-6 produced by induced BJ-5ta cells after exposition to sardine oil (SO) (0.5 and 4 mg/ml), sardine oil with α-tocopherol (SO+αT) (0.5 and 4 mg/ml) and α-Tocopherol (αT) (0.5 and 4 mg/ml) ... 68 Figure 22. LPS induced inflammation of THP-1 cells along time ... 70 Figure 23. ELISA quantification of TNF- α produced by stimulated THP-1 cells after exposition to sardine oil (SO) (0.5 and 4 mg/ml), sardine oil with α-Tocopherol (SO+αT) (0.5 and 4 mg/ml) and α-Tocopherol (αT) (0.5 and 4 mg/ml) ... 71 Figure 24. LPS induced inflammation of THP-1 cells along time ... 73 Figure 25. ELISA quantification of TNF- α produced by stimulated THP-1 cell line previously exposed to sardine oil (SO) (0.5 and 4 mg/ml), sardine oil with α-Tocopherol (SO+αT) (0.5 and 4 mg/ml) and α- Tocopherol (αT) (0.5 and 4 mg/ml) ... 74 Figure 26. Effects of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) in melanogenesis evaluated by the NaOH/DMSO assay in cell line A375. ... 77 Figure 27. Cytotoxicity effect of sardine oil (SO), sardine oil with α-Tocopherol (SO+αT), and α –Tocopherol (αT) in cell line B ... 78 Figure 28. Effect of sardine oil (SO), sardine oil with α-Tocopherol (SO+αT) and α- Tocopherol (αT) in the expression of melanin evaluated by the NaOH/DMSO assay in cell line B ... 80
Figure 29. Effects of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) in the expression of melanin evaluated by the fluorescence assay in cell line B ... 81 Figure 30. Effect of sardine oil (SO), sardine oil with Tocopherol (SO+αT) and α-Tocopherol (αT) in tyrosinase activity in cell line B. ... 82
1.1 Sardine oil and human health
The sardine (Sardina pilchardus) is part of a small group of pelagic fish, which belongs to the family Clupeidae (Figure 1). This fish is abundant along the waters of continental shelf and of Iberian Atlantic. This species has a short longevityof 7-8 years and quickly reaches maturity in the first 2 years of life, with high fertility and a high fat content [1].
The Portuguese coast spawning of this species occurs in October and in May/June. Sardines feed of planktonic crustaceans and other larger planktonic animals and can reach up to 21 cm in length and 60 g in weight [2]. In Portugal sardine fishery is one of the oldest and most abundant, which makes sardines the most important pelagic resource from the Portuguese mainland, both economically and socially [3].
Sardines have important nutritional characteristics, being an excellent source of vitamins, proteins, lipids and minerals. They contain high levels of fat (approximately 13%) and are considered rich-oil fish with higher water content that in warm-blooded animals which makes it more digestible. Their proteic content have all essential amino acids and the lipids are constituted by several fatty acids with nutritional importance that are beneficial to health [2].
Furthermore sardines do not accumulate hazardous substances such as heavy metals and pesticides in their tissues because of their short life cycle and therefore are highly recommendable as a food source for humans [2].
Nowadays, fish is the most common source of ω-3 in human diet, and oily fishes belonging to the Scombridae, Clupeidae and Salmonidae families have the highest percentage of polyunsaturated fatty acids (PUFAs), particularly the eicosapentaenoic and docosahexaenoic acids (EPA, DHA, respectively) and the lowest percentage of saturated fatty acids (Table 1) [4].
Figure 1. Sardines (Sardina pichardus)
Table 1: EPA and DHA contents in some species of fishes(adpted from Rubio-Rodriguez et. al [4]).
Species Common name g/100g food portion
EPA DHA Scomber scombrus Mullus surmulets Sardina pilchardus Salmon salar Gadus morhua Merluccius merluccius Luvarus imperialis Mackerel Red mullet Sardine Salmon Cod Hake Sworfish 1.10 0.91 0.62 0.50 0.23 0.10 0.15 2.56 1.66 1.12 1.00 0.47 0.54 0.30
A diet rich in cold water fishes such as mackerel, salmon, sardines, halibut among others are associated with reduced incidence of coronary heart disease, inhibition of hepatic synthesis, secretion of triacylglycerol, inhibition of platelet aggregation, and prevention of cardiac arrhythmias [5].
1.2 Omega-3 PolyUnsaturated Fatty Acids: the gold members of
the fatty acids family
Fatty acids are organic compounds constituted by a hydrocarbonated chain with a carboxylic group at one end and methyl group at the other, the carboxyl group is reactive and readily forms ester links with the alcohol groups of glycerol or cholesterol, forming acylglycerols (triacylglycerols and phosphoslipids) and cholesterol esters. Fatty acid chain lengths vary from two to thirty or more and may contain double bonds, depending on the nature of the hydrocarbonated chain [6].
The fatty acids can be classified in saturated when have no double bonds between adjacent carbon atoms, monounsaturated when have one bond and polyunsaturated when have two or more double bonds along the chain. In almost all the unsaturated fatty acids, the double bonds are in the cis configuration and are typically positioned at the 3rd, 6th and 9th carbon atom from the terminal methyl group. The cis configuration means that the hydrogen atoms attached to the double bonds are on the same side. When the hydrogen atoms are on opposite sides the configuration is so-called trans [7].
The nomenclature of fatty acids is represented by the number of carbon atoms in the chain, the number of double bonds, and position of the first double bond relative to the methyl carbon (normally, the double bond is represented by ω or n). For example, arachidonic acid (AA) is shown as 20:4 ω-6, describing a molecule with 20 carbon atoms in the chain, four double bonds, with the first double bond at the 6th carbon from the methyl terminus. Thus, this nomenclature allows discrimination of different families of polyunsaturated fatty acids (PUFAs) based on the position of the first double bond in the chain, i.e. omega-3 PUFAS (ω-3 PUFAs) and omega-6 PUFAS (ω-6 PUFAs) [6,8].
The majority of the fatty acids can be synthesized by human body, but there is a group of PUFAs, the essential fatty acids, that the organism cannot produce: omega-3 and omega-6 fatty acids. The α-linolenic acid (ALA, 18: 3 ω-3) and the linoleic acid (LA, 18: 3 ω-6) are the most simple of the omega-3 and omega-6 fatty acids [9].
The α-linolenic acid is synthesized from the linoleic acid by desaturation, catalyzed by desaturase enzyme. The human organism do not possess the desaturase enzyme (delta-15 desaturase) so cannot synthetize α- linolenic acid, thus, these acids are considered as essential fatty acids [10].
Although, humans cannot synthesize α-linolenic acid can metabolize it in series of desaturation (addition of the bond double) and elongation (addition of two carbon atoms). The desaturation occurs at carbon atoms below carbon number 9 (counting from the carboxyl carbon) and mainly occurs in the liver. Figure 2 shows the pathway of conversion of α- linolenic fatty acid in others unsaturated fatty acids. The ALA can be converted to stearidonic acid (18:4 ω-3) by delta-6 desaturase and then elongated to eicosatetraenoic acid (20:4 ω-3) [11].
This fatty acid can be further desaturated by delta-5 desaturase to yield eicosapentaenoic acid (EPA, 20:5 ω-3). The pathway for conversion of EPA in
docosahexaenoic acid (DHA, 22:6 5 ω-3) involves addition of two carbons to form docosapentaenoic acid (DPA, 22:5 ω-3) followed by the addition of another two carbons to produce 24:5 ω-3. After desaturation to form 24:6 ω-3, translocation from the endoplasmic reticulum to peroxisomes and removal of two carbons by limited β-oxidation is formed DHA [12].
There is a higher rate of conversion of ALA to DHA, and some studies in healthy young women have revealed that 21 % of ALA is converted to EPA and 9 % is converted to DHA. Young men, on the other hand, show only an 8 % conversion to EPA and 0-4 % conversion to DHA. This increased efficacy of conversion in young women is attributed to the effects of estrogen. These data suggest that although ALA is considered an essential omega-3 fatty acid, humans can convert EPA and DHA in vivo under certain inducible conditions [13].
1.3 Health benefits of ω-3 PUFAs
Since the 1990s, there has been an increasing interest in the health benefits of ω-3- PUFAS, particularly in their role in the prevention of diseases. The importance of these fatty acids is evidenced by their effects on cardiovascular health which were observed in several studies in populations whose diet consisted mainly of fish. Over the last years, many studies reported the importance of these very long chain (ω-3) fatty acids in the prevention of cardiovascular diseases; in the development of dementia; in inflammatory diseases, in cancer prevention and possibly in the treatment of depression and bipolar disorder [13].
In 2005, was published a general review on the impact of ω-3 PUFAs on human health. This review concluded that, there is strong evidence of the clinical benefit this type of ω-3 PUFAs on cardiovascular diseases [14].
The regular consumption of ω-3- fatty acids reduces significantly the risk of sudden cardiac health. A study reported that individuals with the highest ω-3 PUFAs level in blood had a 90% reduction in sudden cardiac death compared to those with the lowest ω-3 PUFAs levels [15].
These fatty acids also reduce the total mortality and sudden death in patients following myocardial infarction [16]. The beneficial effects of ω-3fatty acids in reducing sudden cardiac death are associated to two principal factors. First, ω-3 PUFAs
reduce cardiac arrhythmias which are responsible for the large part of sudden cardiac deaths [17]. Second, increasing atherosclerotic plaque stability prevents plaque rupture and subsequent coronary events [18].
The increased cell and tissue ω-3 fatty acids content can affect an infinite of molecular pathways, including alteration of physical and chemical properties of cellular membranes, direct interaction with and modulation of membrane channels and proteins, regulation of gene expression via nuclear receptors and transcription factors, changes in eicosanoid profiles, and conversion of ω-3 PUFAs to bioactive metabolites [19].
In Figure 3 is shown a general scheme of the interacting mechanisms whereby ω-3 PUFAs could influence cell function.
Figure 3. General scheme of the interacting mechanisms by which the ω-3 PUFAs might influence cell function [10].
Increased ω-3 fatty acids
Altered membrane fatty acid composition Altered patterrn of eicosanoid synthesis Altered signal transduction patways Altered membrane
sctructure and fluidity
Altered pattern of gene expression
1.4 Role of ω-3 PUFAs in inflammation
Inflammation is a protective response of the body immediately after infection or injury, which is characterized by redness, pain, swelling and heat. Inflammation is characterized by an acute stage that involves a large number of molecular, cellular and physiological changes. This phase is characterized by increased blood flow; increased permeability through the blood vessels, allowing large molecules such as cytokines and antibodies to leave the bloodstream and penetrate the endothelial wall; and increased circulating leukocyte in the blood stream into the surrounding tissue [20,21].
The movement of cells into the inflammatory or infected site is induced by the up-regulation of adhesion molecules such as intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-(ICAM-1), and E-selectin on the surface of endothelial cells. The earliest cells to appear at inflamed sites are granulocytes, with monocytes, macrophages, and lymphocytes appearing later. Granulocytes, monocytes, and macrophages are involved in clearing up cellular and tissue debris, and in tissue repair [22].
The influence of ω-3 PUFAs on the functional responses of the various cell types involved in inflammation and on the production of the range of chemical mediators produced has been studied for many years and a number of effects have been reported. The results observed are highly suggestive that the ω-3 PUFAs act in an anti-inflammatory manner, moreover recent studies suggest that they may be involved in the resolution of inflammation [22,23].
The anti-inflammatory effects of ω-3 fatty acids appear to be mediated by eicosanoids-dependent processes. Increasing intake, in particular of EPA and DHA, will lead to an increase of these acids in cells with a simultaneous reduction of the ω-6 fatty acids[24].
The eicosanoid family of lipid mediators includes derivatives of 20-carbon PUFAs, mainly arachidonic acid (AA). The precursor fatty acid is released from the membrane phospholipids of inflammatory cells. The prostaglandins (PGs) thromboxanes (TXs)and leukotrienes (LTs) are eicosanoids however the most common in the membrane phospholipids of macrophages, neutrophils and lymphocytes is the ω-6 PUFAs arachidonic acid, a usual eicosanoid precursor. The AA is released from the phospholipids by action of phospholipase A2 enzyme, which are activated through inflammatory stimuli [25].
The free AA acts as a substrate for cyclo-oxygenase (COX), lipoxygenase (LOX) or cytochrome P450 enzymes. The COX enzymes, direct to the PGs and TXs, LOX enzymes direct to the LTs and cytochrome P450 enzymes to the hydroxyeicosatetraenoic and epoxyeicosatrienoic acids. Eicosanoids are long recognized as key mediators and regulators of inflammation acting via specific receptors, usually G protein-coupled receptors, and their synthesis and action are targets for a range of non-specific and specific pharmaceuticals [10,22].
The main anti-inflammatory mechanism of EPA is as a competitive inhibitor for pro-inflammatory AA on COX, which produces pro-inflammatory eicosanoids. In figure 4 is represented the anti-inflammatory mechanism of EPA as a competitive inhibitor of AA through the COX enzyme[26]
.
When EPA is subjected to oxygenation by COX, the less inflammatory prostaglandin E3 (PGE3) is generated, while AA generates highly pro-inflammatory prostaglandin E2 (PGE2). As EPA is structurally similar to AA, it can compete with AA to incorporate into the cell membrane phospholipids and become the preferential substrate for COX activity. Therewith, oxygenation of AA by COX can be inhibited, through ω-3 PUFAs and PGE3 is generated instead of PGE2. Therefore, ω-3 PUFAs act as nonsteroidal anti-inflammatory drugs to decrease COX enzyme activities and thereafter decrease inflammation [27,28].
Figure 4. Scheme of anti-inflammatory mechanism of EPA as a competitive inhibitor of AA through the COX enzymes. EPA competitively inhibits PGE2 formation by COX-1 and COX-2.
The levels of PGE2 can be manipulated by increasing the consumption of EPA. Therefore, when COX-associated biochemical reactions are intensively associated with
Less inflammatory Highly inflammatory Araquidonic Acid (AA) Eicosapentaenoic Acid (EPA) PGE2 PGE3 COX COX
EPA competes with AA for COX
EPA due to increased EPA levels and decreased pro-inflammatory AA levels, less harmful and significantly reduced inflammatory status can be expected. Thus, increased ω-3 PUFAs levels compared to ω-6 PUFAs can impart significant beneficial health effects [29].
The ω-3 PUFAs are related with lower levels of inflammation and insulin resistance in both animals and humans. Both EPA and DHA are recognized to have anti-inflammatory activities that decrease the production of pro-inflammatory mediators such as interleukin 1 (IL-1), interleukin 6 (IL-6), interleukin 8 (IL-8) and tumor necrosis factor –α (TNF-α) [30].
The ω-3 PUFAs namely, EPA and DHA are also well known to beneficially alter several metabolic processes including carbohydrate and the lipid metabolism. The ω-3 increases the β-oxidation of fatty acids and inhibit hepatic and lipogenic gene transcription [31,32]. Thus, due to increased fatty acid oxidation, there is induction of
peroxisome proliferator activated receptor (PPAR-α) in the liver, which in turn induces the proliferation of peroxisomes in the liver. DHA and EPA bind to this receptor and act as regulators of gene transcription involved in β –oxidation [33].
It is widely recognized that 80-90% of human cancers can be attributed to environmental and life style factors [34]. Epidemiological studies had shown a lower cancer incidence in association with fish consumption, and fishes rich in ω-3 PUFAs have been a special target due to their chemopreventive effects on carcinogenesis. Human populations with high dietary intake of ω-3 PUFAs enclose lower frequency of breast, prostate and colon cancers, than those who consume low amount of less ω-3 PUFAs consume [34,35].
1.5 Role of ω-3 PUFAs on Oxidative stress
The toxic effects of O2 are directly linked to its partially reduced forms, the reactive oxygen species (ROS), acting on cellular components [36].These reactive species are responsible for the most part of cellular damage and play a key role in the aging process [37].
The production of free radicals and other reactive species derived from the use of O2, in normal physiological conditions are maintained in an equilibrium state by this antioxidant defense system. However, when a
disequilibrium occurs between oxidants and antioxidants in favor of the oxidants, we face a situation defined as oxidative stress.
This disequilibrium can result from two different factors [38]:
- a decrease in antioxidants, due to mutations affecting antioxidant enzymes or depletion of antioxidants along with other essential diet constituents;
- excessive production of O2 and ROS, for example, by exposure to high levels of O2 or by the presence of toxins that are metabolized to produce ROS, among others.
Oxidative stress is able to cause damage to all the biomolecules and ultimately lead to cell death, being implicated in several pathologies.
The ROS can be produced from endogenous sources, such as mitochondria, peroxissomes, and cytocrome P450 and exogenous sources, including environmental agents, pharmaceuticals and industrial chemicals. This oxidative stress can cause DNA, protein, lipid damage conducting to changes in chromosome instability, genetic mutation and modulation of cell growth [39].
Figure 5 illustrates the potential outcomes of reactive species when not counterbalanced by antioxidant defenses of the cell.
Figure 5. Reactive oxygen species and their role in the development of human pathologies [39].
Damage to DNA, RNA protein and lipids
Chromossomal instability, gene mutations ROS Oxidative Stress Exogenous sources - UV lights - Inflammation cytokines - Pathogens Endogenous sources - Mithocondria - Peroxissomes - Cytocrome P450
Altered gene expression
Antioxidants
- Vitamin E - Selenium - Geen tea
PATHOLOGIES
Aging; Cancer; Chronic Inflammation and Autoimmune diseases, Neurological disorders, Fibrotic disease; Infections disease
The excess of ROS can be eliminated by endogenous and exogenous antioxidant molecules. An antioxidant is a substance that when present at low concentrations compared to those of an oxidizable substrate significantly delays or prevents oxidation of that substrate. This substance reductant, therefore, can be classified as a compound capable of preventing the pro-oxidation process, or biological oxidative damage [40].
The nuclear factor erythroid 2-related factor (Nrf2) is the master regulator of gene expression of antioxidant enzymes (Figure 6). Nrf2 regulates cytoprotective responses to endogenous and exogenous stress caused by ROS [41]. This nuclear transcription factor is fundamental for the coordinated induction of those genes encoding many stress-responsive and related proteins, as: NADPH quinone, oxidoreductase-1 (NQO-1), superoxide dismutase (SOD), glutathione S-transferase (GST), glutathione peroxidase (GPx), heme-oxygenase (HO-1), glutamate cysteine ligase (GCL), and catalase [42].
Figure 6. The Keap-1 Nrf2 pathway [43].
The Kelch-like ECH-associated protein (Keap1) is essential for the regulation of activity of Nrf2. In normal conditions, the Nrf2 is constantly degraded via the
ubiquitin-proteossome pathway in a Keap1 dependent manner [43]. Under oxidative and electrophilic stresses, Nrf2 degradation ceases. Stabilized Nrf2 accumulates in nuclei, heterodimerizes with small MAf proteins and activates target genes for cytoprotection and activates target genes involved in the glutathione synthesis, elimination of ROS, xenobiotic metabolism, and drug transport for cytoprotection through antioxidant response element (ARE)/electrophile response element (EpRE). Thus, the levels of Nrf2 protein are regulated by degradation processes, and inducible stabilization of Nrf2 is the essence of cellular response to oxidative and electrophilic stresses [44].
The damage caused by reactive oxygen species at the cellular level, which can trigger cell death by apoptosis and necrosis, depends on the severity of oxidative stress. The expression of target genes of Nuclear Factor-Kappa B (NF-kB) has the ability to promote cell survival (Figure 7) [45].
The NF-kB transcription factor is crucial in a system that allows cells to adapt and respond to environmental changes, a process essential for cell survival. A large number of diverse external stimuli lead to activation of NF-kB and the genes whose expression is regulated by NF-kB, playing important and conserved roles in immune and stress responses; and processes such as apoptosis, proliferation, differentiation, and development [46].
Figure 7. The NF-k B pathway [47].
In its inactive state, NF-kB is in the form of a trimer present in the cytosol. The activation of this transcription factor requires that its inhibitory subunit, IkB, is
phosphorylated. This leads to dissociation of IkB from the NF-kB trimer. The IkB is then degraded, while the remaining NF-kB dimer translocates to the nucleus.
EPA and DHA can inhibit the production of a range of inflammatory proteins, including COX-2, TNF-α, IL-1, IL-6, and IL-8. These inhibitory effects of ω -3 PUFAs seem to involve decreased IkB phosphorylation and decreased activation of NF-kB, effects that appear to involve a reduction in the activation of key early signaling proteins such as mitogen-activated protein kinases. In agreement with the in vitro studies, fish oil feeding to laboratory animals also decreases NF-kB activation and production of inflammatory cytokines [8].
Some studies in healthy human volunteers involving fish oil supplementation showed decreased production of TNF-α, IL-1β, IL-6, and various growth factors by monocytes or mononuclear cells that were stimulated with bacterial endotoxin. These findings indicate that very long-chain ω-3 PUFAs act on inflammatory processes through inhibiting the activation of NF-kB that occurs in response to exogenous inflammatory stimuli [48] .
1.6 Skin
The skin is the primary barrier which protects the body from dehydratation, mechanical trauma and microbial insults, consisting of an outer epidermis and appendages separated from the underlying dermis by a basement membrane [49].
Skin is a complex organ, composed by several tissues and a variety of accessory structures. The primary function of this organ is to prevent excessive loss of water while serving as a permeability barrier. Furthermore, skin is the first organ of the body’s defense mechanisms against external agents helping in the protection against mechanical, chemical, thermal and sunlight injuries in addition to infection by microorganisms. It also displays thermoregulatory sensory and metabolic functions [49]. To repair damaged tissue and replace old cells, skin depends on stem cell populations residing in the adult hair follicle, sebaceous gland, dermis and epidermis[50].
The skin is divided in three functional layers: epidermis, dermis and hypodermis, presented in Figure 8.
Figure 8. Schematic representation of the skin structure [51].
The epidermis (thickness varying from 0.06 to 1 mm) is the most superficial layer comprising a keratinized stratified squamous epithelium. There are four types of cells in the epidermis: keratinocytes, melanocytes, Langerhans cells and Merkel cells, being keratinocytes the most abundant. According to the degree of maturation of keratinocytes, the epidermis can be divided into five layers: basal, spinous, granular, lucid and stratum corneum [51].
The basal layer is the deepest, lying on the basement membrane separating the epidermis from the dermis. Is in this layer and in the deeper part of the stratum spinosum, where occurs the formation of keratinocytes. They continue their maturation along the following layers until the stratum corneum where are coreless and dead cells with the cytoplasm filled with keratin [49].
Below epidermis is the dermis. The dermis (1-2mm deep) is composed of connective tissue, which supports the epidermis and is responsible for the union between the skin and the subcutaneous tissue (hypodermis). There are two distinct layers within the dermis: the papillary and reticular layer. The papillary layer is the most superficial and presents an irregular organization, with overhangs, the dermal papillae. The primary function of the dermal papillae is to increase the surface area of contact between the dermis and the epidermis by reinforcing its connection. Below the
papillary layer is located the reticular layer. Both layers are rich in elastic fibers responsible for skin elasticity. Various structures present on the skin are derived from dermis, such as hair follicles and sebaceous and salivary glands [52]
The hypodermis (1-2mm) is fundamental in uniting the dermis to the adjacent organs and is responsible for the slip on the skin structures as it relies. Another function of the hypodermis is to serve as the power reserve and protection against cold, as a result of adipose tissue richness [51].
Skin condition and functioning are affected by environmental factors, such as ultraviolet (UV) irradiation, free radicals, toxic and allergic compounds, and mechanical damage, and by endogenous factors, such as genetic predisposition, immune and hormone status, and stress. Consequently, skin undergoes alterations resulting in photoaging, inflammation, reduced immune function, imbalanced epidermal homeostasis, and other skin disorders. Skin functioning and skin attractiveness are dependent on nutrition. This is evidenced by the development of skin lesions in response to nutritional deficiencies. Dietary supplementation with the deficient vitamins, minerals, or ω-3-essential fatty acids improves skin conditions in these situations [53].
It is known that when a feed is supplemented with ω -3 fatty acids, they will partially replace ω-6 fatty acids by competing with the same substrate (cyclooxygenase). Thus, there is a reduction in the synthesis of metabolites of ω-6. Either EPA or DHA are present in normal epidermis, because both are metabolized in the epidermis into monohydroxylated metabolites. EPA is metabolized into 15-hydroxyeicosapentanoic acid (15-HEPE) and DHA 17-hydroxydocosahexanoic acid (17-HODHE), which accumulate in the epidermis [54].
Several studies have shown that supplementation with fish oil, containing ω-3 fatty acids, has beneficial effects on skin inflammation. Some of these studies have focused on psoriasis patients, which presented significant improvement in the lesions after supplementation. This improvement is related to the partial substitution of ω-6 fatty acids and their metabolites which induce inflammation, by ω-3 fatty acids, phospholipids at epidermis level [53].
Other studies ω-3 fatty acids, as photoprotective agents, showed a reduction in concentration of PGE2 in irradiated skin. In irradiated skin, supplementation with ω-3
and consequent reduction of PGE2 will be involved, at least in part, in reducing the formation of erythema and partial reduction of immunosuppression [53].
It was also demonstrated in a clinical test that 3 months of fish oil intake lead to a reduction of burns and inflammation from UV radiation. A feed with a higher proportion of ω-3 fatty acids relative to ω-6 fatty acids is also associated with a lower risk of squamous cell carcinoma (SCC) development [55].
1.7 Melanognesis
The melanin production, or melanogenesis, is mainly responsible for skin and hair colour, and plays an important defensive role against the harmful effects of UV radiation of sunlight. However, overproduction and accumulation of melanin results in various skin dermatological disorders including melasma, freckles, age spots and sites of actinic damage or other hyperpigmentations. Melanin is produced normally by melanocytes and, to a greater extent, by melanoma cells. In mammals, melanin is synthesized in the melanosomes of melanocytes located in the basal layer of the epidermis controlled by tyrosinase, which plays a key role in melanogenesis [56].
The tyrosinase (TYR), a copper-containing binuclear enzyme, catalyzes two rate-limiting steps of melanin biosynthesis, in which the L-tyrosinase is hydroxylated of L-tyrosine to L-DOPA (o-diphenol product), and L-DOPA is further oxidized into the corresponding o-quinone [57]. In Figure 9 is shown the melanogenic pathway that occurs within melanosomes.
Melanin is synthesised as dark coloured brown–black insoluble eumelanin or light coloured red–yellow, alkali soluble, sulphur containing phaeomelanin. In both cases the rate-limiting step is the oxidation of tyrosine by tyrosinase in a series of reactions (Figure 9). Eumelanogenesis leads to the formation of indole derivatives. These derivatives include 5,6-dihydroxyindole (DHI) and 5,6-dihydroxyindole-2-carboxylic acid (DHICA), formed from the oxidation of the 1,4 addition product of tyrosinase generated dopaquinone. Intermediates in phaeomelanogenesis include cysteinyldopas [58] .
Figure 9. Melanogenic pathway that occurs within melanosomes, leading to the production of eumelanin and/or pheomelanin [59].
The polyunsaturated fatty acids, especially the ω-3 and ω-6 fatty acids have been extensively evaluated for both their nutritional and cosmetic benefits. PUFAs have been found to be involved in many biological functions in the skin such as prevention of transepidermal water loss, maintenance of the stratum corneum epidermal barrier, and disruption of melanogenesis in epidermal melanocytes, the pigment producing cells in the epidermal-junction of the skin [60].
They have also been reported to regulate protein trafficking in melanocytes. The fatty acids such as linoleic acid and α-linolenic acid and their metabolites have been used in the treatment of various skin disorders [61]. In a series of articles by Ando et al., it has been shown that linoleic and α-linolenic acids accelerate tyrosinase protein degradation through the endoplasmic reticulum-associated degradation (ERAD) pathway in melanocytes thereby inhibiting melanogenesis. Linoleic and α-linolenic acids have also been shown to inhibit hyperpigmentation in in vivo guinea pig skin models using both topical and oral applications [62].
The focus of this master thesis is to potentiate the use of a Portuguese natural resource in the development of future topical applications for cosmetic and therapeutics. Sardine oil is obtained from sardines, a well known fish in the Portuguese culture. Sardine is one of the most common fish of the Portuguese coast with important nutritional features. Sardine oil is also a natural source of nutrients with proven benefits for human health, being rich in ω-3-PUFAs fatty acids, particularly EPA and DHA. Several studies have shown that there is a direct link between a diet enriched in ω-3-PUFAs and the prevention of diseases such as cardiovascular disease, inflammatory conditions, mental disorders and prevention of various types of cancer.
Despite the described benefic effects of PUFAs we want to focus on the anti-oxidant and anti-inflammatory properties of the sardine oil for the future development of topical cosmetic and therapeutic formulations. The extraction of sardine oil from sardines is based on a mechanical process without the need of organic solvents. While for the extraction of PUFAs the process is not as friendly as it involves the use of solvents that could compromise the full benefits of the compounds. During the project was made a comparison between the effect SO and PUFAs in order to assess if SO was so effective as PUFAs.
Briefly, using sardine oil, sardine oil with α-tocopherol, polyunsaturated fatty acids, polyunsaturated fatty acids with α- tocopherol and α-tocopherol we intend to evaluate the effect of these compounds in cells representative of populations of cells from the skin. The interest was first to study the effect of various concentrations of these compounds over a certain period of incubation in cell viability assays and cellular proliferation in order to determine the limiting dose to be used. After selecting the best concentrations of the compounds, it was intended to evaluate their antioxidant properties by DCF and catalase assays as well as the hypothetic involvement in the induction of antioxidant enzymes - HO-1, GCL and NQO1 that result from activation of a key transcription factor - Nrf2 - in the oxidative stress related events.
Another paramount interest of the project was to evaluate the anti-inflammatory potential of sardine oil, sardine oil α-tocopherol and α-tocopherol in the behavior of fibroblasts and monocytes human in response to inflammatory processes.
Ultimately, it was also intended to study the effect of sardine oil in melanogenesis of pigmented cell lines.
3.1 Materials
3.1.1 Chemical Reagents
All reagents used in this work are listed in the following table:
Table 2: Reagents used in this work
REACTIVE AND ABBREVIATION BRAND
Acrylamide/Bis-Acrylamide Sigma Aldrich
Antibiotic Antimycotic Solution (100×) Sigma Aldrich
Bromophenol Blue Sigma Aldrich
Bovine Serum Albumin (BSA) Sigma Aldrich
Bradford Reagent Sigma Aldrich
Celecoxib LC Laboratories
Clarity Western ECL Substrate Bio-Rad
Complete Protease Inhibitor Cocktail Roche
DC Protein Assay Bio-Rad
Dichlorodihydroflurescein Diacetate (DCFH2DA)
Invitrogen
Dimethylsulfoxide (DMSO) Fischer Scientific
Disodium Phosphate (Na2HPO4.12H2O)
Panreac
Dithiothreitol (DTT) Fisher Scientific Dulbecco’s Modified Eagle’s Medium (DMEM) Sigma Aldrich
Dulbecco’s Modified Eagle’s Medium (without phenol red) Biochrom
Ethanol (EtOH) Fisher Scientific
Ethylenediamine Tetraacetic Acid (EDTA) Fluka
Fetal Bovine Serum (FBS) Biochrom
Glycerol Sigma Aldrich
Glycine Fischer Scientific
HEPES Sigma Aldrich
Hoechst Life Technological
Hybond-P polyvinlidene difluoride (PVDF) GE, Healthcare Hydrochloric Acid (HCL) Fisher Scientific
Hydrogen Peroxide (H2O2) Sigma Aldrich
Hygromycin B Sigma Aldrich
Kit Elisa for Human IL-6 Mabtech
Kit Elisa for Human TNF-α Mabtech
Kit Western Blot Detection Merck
L-Dihydroxyphenylalanine (L-Dopa) Sigma Aldrich
L- Glutamine Sigma Aldrich
Lipopolysaccharides (LPS) Sigma Aldrich
Medium 199 Sigma Aldrich
Methanol Sigma Aldrich
Methylthiazolyldiphenyltetrazolium (MTT) Sigma Aldrich
Piroxicam Sigma Aldrich
Polyunsaturated Fatty Acids (PUFAs) kindly supplied by IPMA
Potassium Chloride (KCl) Panreac
Potassium Dihydrogen Phosphate (KH2PO4)
Potassium Hydrogen Phosphate (K2HPO4)
Merck Merck
RPMI-1640 medium (without phenol red) Sigma Aldrich
RPMI 1640 medium Biochrom
Sardine Oil (SO) kindly supplied by IPMA
Sodium Bicarbonate Sigma Aldrich
Sodium Chloride (NaCl) Panreac
Sodium dodecyl sulfate (SDS) Merck
Sodium Orthovanadate (Na3VO4) Sigma Aldrich
Tetramethylethylenediamine (TEMED) Sigma Aldrich Tris(hydroxymethyl)aminomethane (Tris) Fisher Scientific
Trypsin Biochrom
Tocopherol Sigma Aldrich
Tween 20 Sigma Aldrich
3.1.2 Buffers
The buffers solutions utilized in this work are listed:
Phosphate-Buffered Saline (PBS 1x, pH 7.4): 137mM NaCl, 2.7mM KCl, 8.5mM Na2HPO4, 1.5mM KH2PO4
Potassium Phosphate Buffer (PPB 1x, pH 7.0): 31mM K2HPO43H2O, 19.3mM
KH2PO4
RIPA Buffer: 1% NP-40 in 150mM NaCl, 50mM Tris-HCL pH7.5, 2mM EDTA
Towbin Buffer(5×): 25mM Tris, 192mM Glycine
Running Buffer: Towbin Buffer (5×), 0.1%SDS
Transfer Buffer: (5×): 48mM Tris, 39mM Glycine, 0.0375%SDS
Transfer Buffer: Transfer Buffer (5×), absolute methanol
TPBS: PBS 1x, 0.05% Tween 20
Dithiothreitol Buffer (DTT): 1000mM DTT
Incubation Buffer (IB): PBS, 0.05% Tween 20, 0.1% BSA
Laemmeli Sample Buffer (5×): 250mM Tris-HCl (pH 6.8), 50% Glycerol, 10% SDS, 0.1% Bromophenol Blue
3.1.3 Biological Materials
3.1.3.1 Cell lines
In this project were used several cell lines as representative populations of skin cells:
Human skin fibroblasts (BJ-5ta)
Human skin melanoma (A375)
Human skin melanoma (B)
Human skin keratinocytes (NCTC2544) And one cell line from human peripheral blood:
Human monocytes (THP-1)
All this cell lines were obtained from American Type Culture Collection (ATCC), except the B cell line that was kindly supplied by Doctor Francisco Real from the Epithelial Carcinogenesis Group, Centro Nacional de Investigaciones Oncológicas, Madrid (Spain) and NCTC2544 cell line that was supplied by Instituto Zooprofilattico Sperimentale Della Lombardia e Dell’Emilia-Romagna, Pavia (Italy).
3.1.3.2 Antibodies
The antibodies used on this work are listed in following table:
Table 3: Antibodies employed and suppliers PRIMARY ANTIBODY SECONDARY ANTIBODY (IgG) BRAND
β- Actin Mouse Sigma Aldrich
Pro-caspase-3 Rabbit EMD Milipore Corporation
GCLC Rabbit Santa Cruz Biotecnhogical
GCLM Mouse
Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University
Histone-H1 Mouse Santa Cruz Biotechnology
HO-1 Mouse Enzo Life Sciences
NQO1 Mouse Santa Cruz Biotechnology
Nrf2 Rabbit Novus Biologicals
3.1.3.3 ELISA kits
Two ELISA kits were used in this study for the quantification of Interlueukin-6 (IL-6) and for the quantification of Tumor Necrosis Factor- alpha (TNF-α). The ELISA kits were composed of four antibodies:
Table 4: Constitution of IL-6 and TNF-α kits
Elisa for Human IL-6 Elisa for Human TNF-α
Monoclonal antibody (1 mg/ml) Monoclonal antibody (0.5 mg/ml)
Biotinylated monoclonal antibody (1 mg/ml)
Biotinylated monoclonal antibody (0.5 mg/ml)
Streptavidin-Horseradish Peroxidase Streptavidin-Horseradish Peroxidase
Recombinant human IL-6 standard Recombinant human TNF-α standard
3.2 Cell culture maintenance
BJ-5ta cell line (normal human skin fibroblasts immortalized by overexpression of telomerase), was cultured in four parts of Dulbecco’s modified Eagle’s medium (DMEM) (powder), containing L-glutamine, D-glucose, sodium bicarbonate and 1 part of medium 199, supplemented with 10% FBS, 1% (v/v) penicillin/streptomycin solution and 10 µg/mL hygromycin.
A375 cell line (human skin melanoma), was cultured in DMEM (liquid and without phenol red), containing 3.7% NaHCO3, 4.5% D-Glucose,
supplemented with 10% FBS, 1% (v/v) antibiotic/antimycotic solution and L-glutamine.
NCTC2544 cell line (human skin keratinocytes), was cultured in DMEM containing L-glutamine, D-Glucose and sodium bicarbonate, and supplemented with 7% FBS and 1% (v/v) antibiotic/antimycotic solution.
B cell line (human skin melanoma), was cultured in RPMI 1640 medium (powder), containing L-glutamine without phenol red and sodium bicarbonate, supplemented with sodium bicarbonate, HEPES, 10% FBS not inactivated and 1% (v/v) antibiotic/antimycotic solution.
THP-1 cell line (human peripheral blood monocytes), was cultured in RPMI-1640 medium (liquid), containing 2.0% NaHCO3 without L- glutamine, supplemented with 10% FBS, L-glutamine and 1% (v/v) antibiotic/antimycotic solution.
Cellular subcultures were performed when confluence reached values close to 80-90% while for the cell line suspension THP-1 when confluence reached values close to 8×105 cells/ml. Adherents cells (BJ-5ta, A375, NCTC2544 and B) were maintained in 25cm2 tissue culture flasks and the suspension cells THP-1 were maintained in plate dishes 100mm. The cell lines were maintained in an incubator at 37°C in a humidified atmosphere with 5% CO2. The cell culture medium was renewed two times per week. For subcultures and plating, the adherent cells were detached with a trypsin solution 0.05%, and fresh medium was added in order to neutralize the trypsin. The cell suspension was centrifuged 5 min at 160g. The supernatant was discard and fresh medium was added to obtain a new cell suspension. The cell suspension was loaded in a
Neubaeur chamber and the concentration of cells present in the suspension was
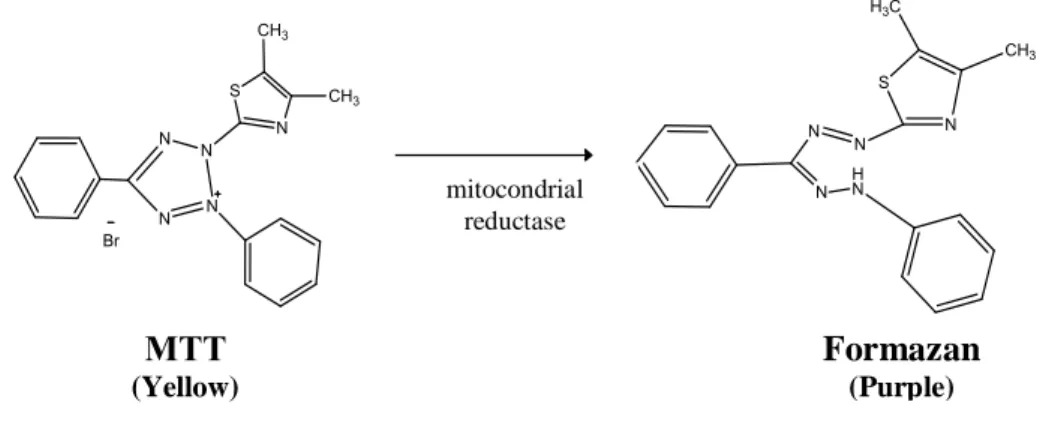
mitocondrial reductase Formazan (Purple) MTT (Yellow)
3.3 Cell viability assessed by MTT assay
The MTT assay is a colorimetric method used to measure the toxicity of different substances in cells. The principle of this assay is that in viable cells, mitochondrial activity is constant and therefore an increase or decrease in the number of viable cells is linearly related to mitochondrial activity. The mitochondrial activity of the cells is reflected by the conversion of the tetrazolium salt (MTT) into formazan crystals. The amount of formazan reflects the reductive potential of the cytoplasm and therefore cell viability [63].
Figure 10. Representation of the reaction of enzymatic conversion of MTT to formazan.
Cells were seeded at a density of 10000 cells/well for cell lines A375, and B and 7500 for NCTC2544, on a 96-well tissue culture plate, the day before the experiments. The cells were exposed to different concentrations of SO, SO with α-tocopherol, PUFAs, PUFAs with α-tocopherol and α-tocopherol.
For A375 and NCTC2544 cell lines, four concentrations of SO, SO with α-tocopherol, PUFAs, PUFAs with α-tocopherol and α-tocopherol were tested. In B cell line the PUFAs were not tested. The concentrations are presented in the table 4.
Table 5: Concentrations of SO, SO with α-Tocopherol, PUFAs, PUFAs with α-Tocopherol tested in cell lines A375, NCTC2544 and B
SO SO with α-Tocopherol PUFAs PUFAs with α-Tocopherol 0.5 mg/ml 0.5 mg/ml 0.025 mg/ml 0.025 mg/ml 4 mg/ml 4 mg/ml 0.1 mg/ml 0.1 mg/ml
8 mg/ml 8 mg/ml 0.5 mg/ml 0.5 mg/ml
20 mg/ml 20 mg/ml 1 mg/ml 1 mg/ml
The concentrations of α-Tocopherol correspond to the amount added to SO and PUFAs. For a better comprehension the concentrations of α-Tocopherol are expressed in the corresponding SO and PUFAs concentrations. Two concentrations of SO and two concentrations PUFAs (0.025 mg/ml of PUFAs correspond 0.025 mg/ml α-Tocopherol; 0.5 mg/ml PUFAs correspond 0.5 mg/ml α-Tocopherol; 4 mg/ml SO correspond 4 mg/ml α-Tocopherol and 20 mg/ml correspond 20 mg/ml α-Tocopherol).
For the B cell line the concentrations of α-Tocopherol are expressed in the corresponding in the SO concentrations (0.5 mg/ml SO correspond 0.5 mg/ml α-Tocopherol; 4 mg/ml SO correspond 4 mg/ml α-α-Tocopherol; 8 mg/ml SO correspond 8 mg/ml α-Tocopherol and 20 mg/ml SO correspond 20 mg/ml α-Tocopherol).
Cells incubated with DMSO (30% of the total volume) and cells without the addition of the compounds were used as controls; control of death and control of life respectively. Cells were incubated at 37°C in a humidified atmosphere with 5% CO2. At the end of 24 and 48 hours of contact, cell metabolic activity was assessed by MTT viability assay.
After incubation with samples, 110 µl of a MTT solution (5 mg/ml diluted in medium) was added to each well, and cells were incubated at 37°C for 2 hours. The MTT solution was carefully decanted, and formazan crystals were dissolved in 110 µl of a DMSO/EtOH (1:1(v/v)) mixture. Colour was measured with 96-well plate reader at 570 nm in a microplate reader SpectraMax Plus (Molecular Devices).
3.4 Cell proliferation assessed by Hoechst dye assay
The Hoechst 334342 (2’-[4-ethoxyphenyl]-5-[4-methyl-1-piperazinyl]-2,5’-bi-1Hbenzimidazoletrihydrochloride trihydrate) is a cell-permeable DNA stain that binds preferentially to adenine-thymine (A-T) regions of DNA, binding to the minor groove of DNA and is excited by ultraviolet light, emitting blue fluorescence at 460-490 nm [64].

![Figure 3. General scheme of the interacting mechanisms by which the ω-3 PUFAs might influence cell function [10]](https://thumb-eu.123doks.com/thumbv2/123dok_br/17678148.826212/27.893.148.754.501.948/figure-general-scheme-interacting-mechanisms-pufas-influence-function.webp)


![Figure 6. The Keap-1 Nrf2 pathway [43].](https://thumb-eu.123doks.com/thumbv2/123dok_br/17678148.826212/32.893.131.763.534.1019/figure-the-keap-nrf-pathway.webp)
![Figure 7. The NF-k B pathway [47].](https://thumb-eu.123doks.com/thumbv2/123dok_br/17678148.826212/33.893.238.695.674.1009/figure-the-nf-k-b-pathway.webp)
![Figure 8. Schematic representation of the skin structure [51].](https://thumb-eu.123doks.com/thumbv2/123dok_br/17678148.826212/35.893.204.703.113.495/figure-schematic-representation-skin-structure.webp)
![Figure 9. Melanogenic pathway that occurs within melanosomes, leading to the production of eumelanin and/or pheomelanin [59].](https://thumb-eu.123doks.com/thumbv2/123dok_br/17678148.826212/38.893.229.688.148.453/figure-melanogenic-pathway-melanosomes-leading-production-eumelanin-pheomelanin.webp)