DAMAGES AND ECONOMIC IMPACT OF GRAPEVINE V IRUS DISEASES
Texto
Imagem
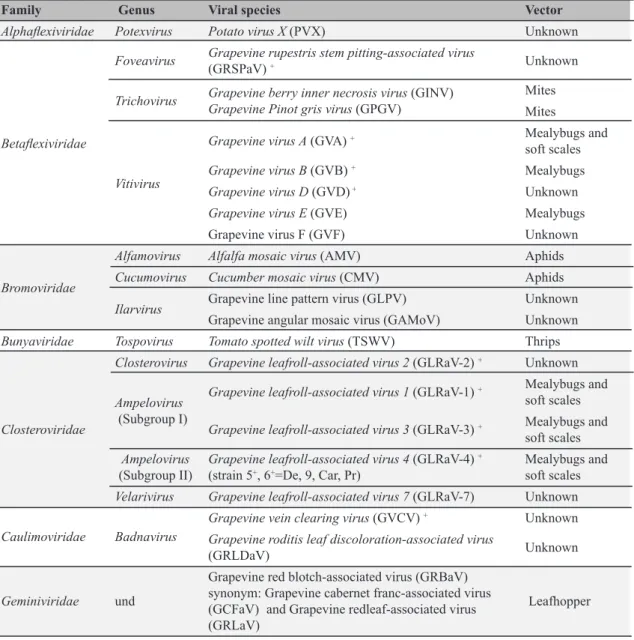
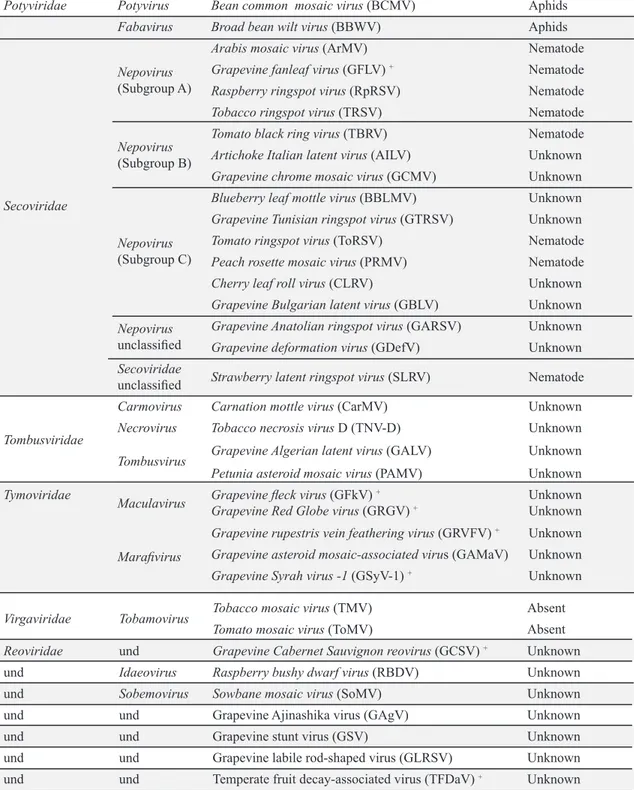
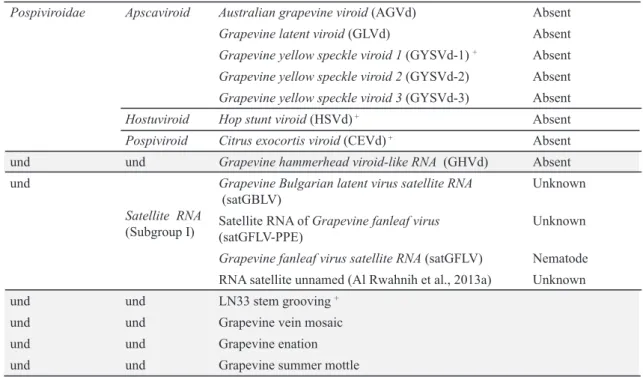

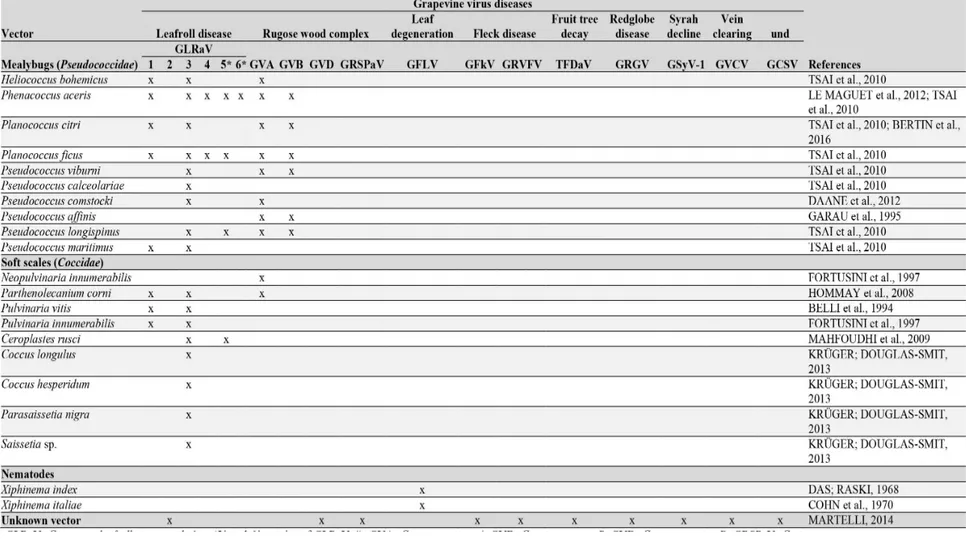
Documentos relacionados
João IV, a quem se prognostica o prometido império, que o orador traduz pela perífrase de "infinitas felicidades, muito maiores ainda, do q foraõ nossas desgraças tegora"
The process of building the Faultload database is executed offline, and must pre- cede the actual testing phase, as a set of test values must be created for each unique parameter
The nurses agreed with the universal access to technology, which should also consider the results of their use and the professional opinion based on what is best for the
Expression of Grapevine leafroll-associated virus 3 coat protein gene in Escherichia coli and production of polyclonal
The present study describes the coat protein (CP) gene sequences of four GLRaV-3 isolates sampled from grapevines grown in the São Francisco River basin,
c antisera against Brazilian isolates of Grapevine leafroll-associated virus 2 (GLRaV-2) and Grapevine virus B (GVB), developed from expressed coat proteins (CPs) in
This analysis, using the NJ method, included a total of 15 CP sequences of local isolates of GRSPaV and 12 reference isolates (sequence variants) retrieved from the
Real-time RT-PCR (TaqMan) assays for the detection of Grapevine leafroll associated viruses 1-5 and Figure 1 - Quantification of Grapevine leafroll-associated virus 4 (GLRaV-4)
