PASTOS:
FUENTE NATURAL DE ENERGiA
Alfredo Calleja Suarez
Ricardo Garcia Navarro
Angel Ruiz Mantec6n
Rodrigo Pelaez Suarez
U N IV E R S ID A D D E L E O N A r e a d e p u b lic a c io n e s
R e u n i6 n Iber ic a d e P a s to s y F o r r a je s ( 4a. 2 0 1 0. Z a m o r a, M ir a n d a d o D o u r o)
P a s to s : fu e n te n a tu r a l d e e n erg ia : 4a R e u n i6 n Iber ica d e P a s to s y F orr a je s, 3 - 6 m a y o 2 010, Z a m o r a - Mir a n d a d o D o u r o = [4a] R e u n ia o Ib e r ic a d e P a s ta g e n s e fo r r a g e n s, 3 - 4 m a io 2 0 1 0, [Z a m o r a - M ir a n d a d o D o u r o ] IA lfr e d o C a lle ja S u a r e z ... le t. a l.] ( c o o r d ..). - [L e 6 n ] : U n iv e r s id a d d e L e 6 n, A r e a d e P u b lic a c io n e s ; [E s p a n a ] : S o c ie d a d E s p a n o la p a r a e l E s tu d io d e lo s P a s to s, 2 0 1 0
5 4 8 p. :g r a f., ta b /a s ; 28 e m.
in d ic e p o r a u to r e s. - B ib lio g r a f. A I fin a l d e c a d a c a p. - T exto s e n ca s te lla n o, p o r tu g u e s e in g le s
IS B N 9 7 8 - 8 4 - 9 7 7 3 - 5 02- 5
1. P a s to s - Exp lo ta c i6 n-C o n g r e s o s. I. C alle ja S u a r e z, A lfr e d o . II. U n iv e r s id a d d e L e 6 n. A r e a d e P u b lic a c io n e s . III. S o c ie d a d E s p a n o la p a r a e l E s tu d io d e lo s P a s to s . IV. R e u n ia o Ib e r ic a d e P a s ta g e n s e F o r r a g e n s ( 4a. 2 0 1 0 . Z a m o r a )
6 3 3.2 ( 0 6 3 )
© U n iv e r s id a d d e L e 6 n
A r e a d e P u b lic a cio n e s © L o s a u to r e s
E d ita : S o c ie d a d E s p a n o la p a r a e l E s tu d io d e lo s P a s to s
E d ic i6 n c o o r d in a d a p o r A lfr e d o C a lle ja S u a r e z, R ic a r d o G a r c ia N a v a r r o,
A n g e l R u iz M a n te c 6 n, R o d r ig o P e la e z S u a r e z
IS B N : 9 7 8 - 8 4 - 9 7 7 3 - 5 0 2 - 5
D e p 6 s ito le g a l: L E - 6 6 7 - 2 0 1 0
PAST AGENS PERMANENTES
SEMEADAS BIODIVERSAS
RICAS EM
LEGUMINOSAS:
COMPOSI<;AO
FLORISTICA
AO LONGO DE UM
GRADIENTE
MESOTOPOGR.AFICO
C.AGUIAR, 1. PIRES, M.ANGELO RODIGUES & Ma
ESTER FERNANDEZ-NllNEZ
Centro deInvestigayao de Montanha (CIMO), ESA - Instituto Politecnico de Braganya, Apartado
1172, 5301-855 Braganya, Portugal - e-mail: cfaguiar@ipb.pt mailto:
Admite-se que as Pastagens Permanentes Semeadas Biodiversas Ricas em Leguminosas
(PPSBRL) sejam superiores as misturas simples a perseguir e a ajustar-se a heterogeneidade espacial
caracteristica dos solos mediterranicos. Testaram-se duas hip6teses que emergem directamente desta
proposiyao: 1) as especies das misturas de PPSBRL acomodam-se as caracteristicas estacionais
controladas por gradientes mesotopognificos. 2) Esta acomodayao ocorre escala da especie e da
cu!tivar (cv.).
Numa PPSBRL estabilizada avaliou-se 0 efeito de urn gradiente mesotopografico na
aglomerayao de especies pratenses indigenas ou melhoradas, e de morfotipos T . s u bte r r ane u m,
definidos apartir das cv. da mistura original de sementes. A "posiyao fisiografica" exerceu urn efeito
significativo naestrutura dos dados floristicos. A percentagem de cobertura relativa das especies edos
morfotipos das cv. precoces de T. su b ter r aneum foi superior no cimo de encosta. As especies mais
exigentes em humidade e osmOlfotipos das cv. deflorayao mais tardia exibiram coberturas maximas a
meia ou no fundo-de-encosta. Os resultados, embora contingentes, suportam ambas hip6teses. A
segregayao espacial dos gen6tipos melhorados tern, certamente, urn papel fundamental na
superioridade agron6mica das misturas complexas desementes pratenses.
Os trevos que nao enterram as sementes e as gramineas melhorados tern dificuldade em
persistir em pastagens sujeitas aurn pastoreio intensivo e continuo.
Palavras chave: gramineas, leguminosas, T .su b ter r a n e u m, ecologia depastagens
INTRODUC<;::Ao E OBJECTlVOS
A evidencia empirica acumulada nas u!timas decadas (e.g. Clark, 2001, Sebastia et a l ., 2004 e
Tilman, 2006) indicia que as misturas complexas de sementes pratenses sao mais produtivas, menos
permeaveis a especies aut6ctones de baixo valor alimentar e mais resistentes a flutuayoes c1imaticas
extremas do que as misturas simples. Embora a interacyao diversidade-funyao seja urn terna classico
da ecologia das comunidades, em pastagens, como em outros tipos de ecossistemas, grande parte dos
trabalhos publicados foram realizados a pequenas escalas espaciais e em condiyoes experimentais
controladas (Symstad et a l ., 20 0 3).
David Crespo, de algum modo inspirado na estrutura floristica dos prados naturais,
desenvolveu com sucesso nos (l!timos 30 anos urn sistema inovador de pastagens: Pastagens
Pennanentes Semeadas Biodiversas Ricas em Leguminosas (PPSBRL), caracterizado pelo uso de
misturas complexas de sementes, por vezes recorrendo a mais de duas dezenas de especies/gen6tipos
de gramineas e1eguminosas. Tendo em considerayao a diversidade de especies/gen6tipos e expectavel
que em condiyoes de campo, asPPSBRL sejam superiores as misturas simples de sementes pratenses a
mediterranicos. Com base num dispositivo experimental simples, procurou-se testar duas hip6teses
que emergem directamente da proposiyao anterionnente enunciada: 1) as especies das misturas de
PPSBRL acomodam-se as caracteristicas estacionais controladas por um gradiente mesotopogrMico. 2) Esta acomodayao ocon'e a escala da especie e da cultivar (cv.).
MA T E R I A L E M E T O D O S
A experiencia decorreu na Quinta da Franya (Covilhil, Portugal), na regiao da Cova daBeira, a
leste da Serra da Estrela (400
16' N; 70
30' W), numa PPSBRL semeada em 2001, ja estabilizada, anualmente fertilizada com ca. 27 kgP20s/ha eintensamente pastoreada par bovinos e ovinos durante todo 0 cicIo produtivo. A pastagem esta situada numa encosta granitica de declive suave (2-3%), na
qual se distinguem tres posiyoes fisiograficas marcantes, ao longo de urn gradiente mesotopografico
( s e n s u Billing, 1973): cimo-de-encosta, meia-encosta e fundo-de-encosta. Este gradiente esta,
naturalmente, correlacionado com a espessura e a disponibilidade de agua no solo, crescentes no senti do cimo-de-encosta - fundo-de-encosta. Em cada uma das tres posiyoes fisiograficas colocara
m-se quatro quadrados pennanentes, protegidos par caixas de rede quinze dias antes da amostragem
floristica, perfazendo urn total de 12 quadrados (areas de amostragem). As plantas vasculares
detectadas na amostragem foram identificadas ao nivel da especie ou do morfotipo, no caso do T.
subter r a n eu m. Em Maio de 2008 0 grau de cobertura relativo de cad aespecie/morfotipo foj estimado
pelo metodo do ponto quadrado. 0 grau de cobertura relativo foi calculado dividindo 0 nlunero de
toques da especie/morfotipo pelo numero total de toques em plantas vasculares realizados noquadrado (Mueller-Dombois & Ellenberg, 1974).
o
T. su b ter ra n e u m e uma especie c1eistogamica, sendo as suas populayoes naturaisconstituidas por linhas puras (Katznelson, 1974). A cleistogamia obrigat6ria pr6pria da especie
implica que as caracteristicas geneticas das cultivares comerciais se mantem ap6s a sementeira. Pela
mesma razao, cultivares (cv.) morfologicamente consistentes podem ser reconhecidas no campo muitos anos ap6s a instalayao das pastagens. De modo a caracterizar morfologicamente as cv. de T.
s u bter r a n e u m presentes na Quinta da Franya, no Outono de 2007 semeou-se em estufa, na Quinta de Sf Apol6nia (Braganya, Portugal), uma colecyao de sementes extremes cedida pela empresa
FERTIPRADO. Para emular a perturbayao pela herbivaria, os T. s u bterra neu m semeados foram
sujeitos a um corte manual mal ultrapassavam os 5 cm de altura. As plantas foram colhidas e
prensadas no final do Inverno, inicio da Primavera de 2008. Com base na observayao das
caracteristicas morfol6gicas externas foram definidos quatro morfotipos e construida uma chave
dicot6mica de identificayao (Tabelal). Foi possivel discriminar as cv. 'Losa', 'Gosse' e 'Denmark'. As cv. 'Campeda' e 'Woogenellup' foram agregadas num unico morfotipo porque alem de serem morfologicamente semelhantes tem uma precocidade similar. 0 T. s ub te r r ane um 'Gosse' e uma selecyao da subespecie o x a l o i d e s (= subsp. yan n i n i c u m ); as restantes cv. pertencem a subsp. s u bter r a n e um . A morfologia externa e insuficiente para uma identificayao absoluta das cv. de T . s u b ter r a neu m em condiyoes de campo porque os Tr i f o l i u m sao por natureza morfologicamente muito
Tabela 1. Chave dicotomica para as cv. de T . s u b ter r a n e u m cultivadas na Quinta da Fran~a
I.Caules hirsutos 'Losa"
-Caules glabros a esparsamente vilosos 2
_.Peciolos glabros; estipulas glabras; marca d'agua geralmente pouco evidente 'Denmark' -Peciolos vilosos a esparsamente vilosos; estipulas glabras ou vilosas; marca d'agua evidente 3
~. Peciolos esparsamente vilosos; estipulas glabras; foliolos geralmente mgosos, com amarca d'agua
.uase horizontal atingindo a margem 'Gosse'
- Peciolos vilosos; estipulas hirsutas; foliolos geralmente nao mgosos emarca d'agua concentrada no
-entro dos foliolos 'Campeda' + 'Woogenellup'
Os dados floristicos foram explorados com metodos multivariados de ordenayao,
'sponibilizados pelo pacote estatistico CANOCO (Tel' Braak & Smilauer, 2002). A aplicayao aos
dados totais de uma DCA ("Detrended Correspondence Analysis") demonstrou que 0 "turnover" de
-pecies (diversidade beta) era baixo, e que seria adequado 0 usa de metodos lineares de ordenayao,
.::oncretamente deuma RDA (analise de redundancia) (Tel'Braak & Smilauer, 2002). Antes decorrer 0
CA OCO foram construidas duas matrizes de dados: "amostras (quadrados permanentes) x
;-~rcentagem de cobertura relativa das especies/morfotipos" e "amostras x variaveis explanat6rias".
::oi considerada uma unica variavel explanat6ria semi-quantitativa: "posiyao fisiogrMica", com tres
2veis fundo-de-encosta, meia-encosta e cimo-de-encosta, cuja significancia naestruturayao dos dados
:-oristicos foi testada com permutayoes de Monte Carlo. Os dados da matriz original "amostras x
:::<:'rcentagem de cobertura das especies/ morfotipos" resultam de contagens e pOI' essa razao foram
'{}garitmizados (Leps & Smilauer, 2003).
RE S UL TAD O S E D Is c u s sAo
Numa primeira RDA (figura I) constatou-se que 0 primeiro elXO da ordenayao (EX1) :::xplicava uma parte muito significativa da variancia das especies/morfotipos: 22% (Valor Pr6prio
_-r]=0,22), urn valor superior it variancia explicada pelo E X2 (19%, VP=O,19). Uma vez que se _ :lsiderou uma unica variavel explanat6ria - a"posiyao fisiografica" - 0 EX1e0 unico eixo can6nico
.: modelo multivariado explicitado na figura 2; os demais eixos sao "livres" e herdam a variancia
:=-s·dual. A significancia do E X1 e, implicitamente, da "posiyao fisiografica" no controlo da
~omerayao ("assembling") das especies/morfotipos, foi confirmada com permutayoes de Monte
=-2.1'10(p<0,006). 0 bipJot "especies/morfotipos -variaveis explanat6rias" mostra que os vectores das
::''Ontas indigenas menos exigentes em agua (e.g. P l a n t a g o c o r o n o p u s e On o n is s p i n o s a ) foram
~pados no lado esquerdo da figura, enquanto os vectores das plantas meso-higr6filas, muitas das
'_ is perenes, se concentraram no lado direito (e.g G a u d in i a f rag il is, Holcus lanatu s, T r if oli um ~ biu m, P lant a g o l an ceo l a t a e T. c e rnu um). 0 diagrama de ordenayao (figura I) patenteia que 0
::::meiro grllpo de especies emais abundante no cimo-de-encosta, enqllanto as coberturas relativas do
cgundo sao maximas nos solos hUmidos de fundo-de-encosta. 0 fundo-de-encosta esta sujeito a
:harcamentos prolongados na epoca das chuvas, assim 0 indica 0vector do T. c e rnu um.
Urn padrao analogo ocorre com asplantas semeadas (figura 2). 0 O r n i t h o p us co m p res s u s e os
-rlotipos caracteristicos das cv. de T.su b ter r a n eum mais precoces ('Losa' e 'Gosse') evidenciam-se
metade direita da figura 2, enquanto os vectores das especies pratenses melhoradas mais exigentes
cv. de T. su b te rraneum de cicio fenol6gico intennedio ou tardio ('Denmark' e tipo 'Camped2
'Woogenellup'), apontam para 0 lado esquerdo da figura. Por conseguinte, a cobertura relativa
morfotipos das cv. precoces de T. su b ter r a neu m foi maxima no cimo de encosta; as especies m _
higr6filas e osmorfotipos das cv. mais ser6dias de T. su bter r aneum dominaram a meia encosta ou ~
fundo de encosta (Tabela 2).
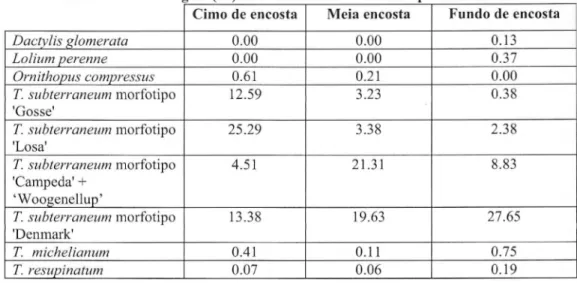
Tabela 2. Percentagem (%) de cobertura relativa das especies semeadas
Cimo de encosta Meia encosta Fu n d o de encosta
D actylis [ <lomerat a 0.00 0.00 0.13
Lol ium peren ne 0.00 0.00 0.37
Orn ith ovu s c omvres sus 0.61 0.21 0.00
T .s u bter r aneu m morfotipo 12.59 3.23 0.38
'Gosse'
T.s ub ter ra neum morfotipo 25.29 3.38 2.38
'Losa'
T.s u b t e r r a neum morfotipo 4.51 21.31 8.83
'Campeda' +
'Woogenellup'
T.s u b t e r r a neum morfotipo 13.38 19.63 27.65
'Denmark'
T. m i che li an u m OAI 0.11 0.75
T.re supin a t u m 0.07 0.06 0.19
o morfotipo das cv. de precocidade intennedia 'Campeda' e 'Woogenellup' apresentou urn
maximo na meia-encosta (Tabela 2). As estatisticas da RDA das plantas pratenses semeadas foram
tambem analogas as da RDA com todas as especies. Mais uma vez 0 EX1 explicou mais variancia do
dados das especies/morfotipos do que 0 EX2, respectivamente 28,1% e 27,6%. A "posiyao
fisiografica" exerceu um control0 significativo sobre a disposiyao espacial das especies/ morfotipo
pratenses melhorados (p<0,004, pennutayoes de Monte Carlo).
No Tabela 2 constata-se ainda que na PPSBRL da Quinta da Franya os trevos que nao
enterram os frutos eas gramineas melhorados apresentam coberturas relativas muito baixas, sobretudo
nos solos mais secos deposiyoes fisiograficas mais convexas(cimo-de-encosta).
CONCLUSOES
Na Quinta da Franya a pOSlyaO fisiografica a pequena escala controlou a aglomerayao
("assembling") espacial das plantas pratenses melhoradas, ou nao. As especies melhoradas mais
exigentes em humidade (D. glomer ata, 1 . p e ren ne , T. re su p inatum eT . m ich eli anu m) evitaram 0 cimo
de encosta e concentraram-se no fundo-de-encosta. A segregayao espacial dos morfotipos de T.
su b ter r a n eu m esta, aparentemente, correlacionada com a precocidade da florayao e da produyao de
fruto: os morfotipos das cv. precoces ('Losa' e 'Gosse') foram mais abundantes no cimo de encosta,
em solos mais secos, enquanto os morfotipos das cv. mais tardias ('Denmark' e tipo 'Campeda' +
'Woogenellup') encontraram um 6ptimo ecol6gico local ameia-encosta ou no fundo-de-encosta.
As especies/cv. melhoradas das misturas de PPSBRL sao distribuidas de fonna unifonne
aquando da sementeira; com 0 tempo segregam-se espacialmente, em funyao das caracteristicas do
solo. Os resultados da experimentayao, embora contingentes, sustentam as duas hip6teses fonnuladas
na introduyao. Assim sendo, 1) 0 nicho realizado (vd. Keddy, 2007) nao e comum a todas
':esempenhani urnpapel determinante na estruturayao espacial da flora das PPSBRL, tanto ao nivel da
~:pecie como da cultivar. A segregayao espacial especies/cv. pratenses melhoradas tern, certamente,
~ papel fundamental nas caracteristicas agron6micas superiores das misturas complexas de sementes
..=atenses.
A flora melhorada das PPSBRL enfrenta POl'em duas importantes contrariedades: os trevos
_._"nao enterram as sementes e as gramineas tern dificuldade em persistir em pastagens sujeitas a urn
_ -, oreio intensivo e continuo.
_~GRADECIMENTOS
-~alho financiado pela FCT-Fundayao Para a Ciencia e Tecnologia, projecto PTDC/AG
R---.....;'.1.169637/2006 "BioPast- Uma Abordagem Integrada as Pastagens Pennanentes Biodiversas Ricas
Leguminosas".
-~FERENCIAS BIBLIOGRAFICAS
-
=
:::
.
1
-
GS, W. D., 1973. Arctic and alpine vegetations: similarities, differences, and susceptibility to:mbance. Bi oSci e nc e 23, 697-704.
--=--\RK, E.A., 2001. Diversity and stability in humid temperate pastures. Em: Com pet i t ion a n d S uc
-_ -ion i n Pa st ur e s, 103-118. Ed.P. TOW, A. LAZENBY. CABI Publishing.
'=::L'\-WELL,W.K.; SCHW ILK, D.W.; ACKERLY, D.D., 2006. A trait-based test for habitat filt
er-_. -onvex hull volume. Ec olo g y 87, 1465-1471.
_-~~LSON 1. 1974. Biological flora ofIsrael. 5. The subterranean clovers of Trifol i um subsect.
- ~ sJmOl ph u m Katzn. Tr!foli um s ub te rraneum L. (sensu 1ato).Isr . J Bot . 23, 69-108.
i{::::::J:JY,P.A., 2007. P lan ts and Ve g et at ion Origi ns , Pr o ces s e s , C onseq uenc e s. Cambridge University
-':=-LER-DOMBOIS, D.; ELLEMBERG, H., 1974.Ai m s an d M e tho ds o f V e g et at i o n Eco l o gy . John
.Sons.
~. TIA.,M . T. et al.,2004. Higher yield and fewer weeds in grass/legume mixtures than in mono
-~-- 12 sites of COST action 852. Em: L a n d U se Sy stems in Grasslan d Dom ina ted Re gi ons 48 3
-~. . LUESCHER et al. Book ofAbstracts.
-~AD, A. 1., et a!., 2003. Long-term and large-scale perspectives on the relationship between
,_ ityand ecosystem functioning. Bi o S cience , 53, 89-98.
__ --....'\D;.., 2006. Biodiversity and ecosystem stability ina decade-long grassland experiment. Na
-_-.1. 629-632.
- "~BIODIVERSE PERM ANENT PASTURES RICH IN LEGUM ES: FLORISTIC
COMPOSTION ALONG A M ESOTOPOGHRAPHYCAL GRADIENT
· l-\RY
-:1le environmental tuning of Sown Biodiverse Permanent Pastures Rich in Legumes
) to Mediterranean soils heterogeneity is in general considered superior to simple mixtures.
_?ltheses that emerge from this proposition were tested: I) the SBPPRL species segregate sp
a-cording to the soil characteristics, along a mesotopographic gradient; 2) this spatial accommo
-- ,:urs atspecies and cultivar (cv.) levels.
- a stabilized SBPPRL was evaluated the effect of a mesotopographic gradient in the assem
the cv. present in the original seed mixture. The explanatory variable "physiographic position" ex
-erted asignificant effect inthe florist data structure. The species more exigent insoil humidity and the
morphotypes of the late flowering T . su bte r r a n e u m cv. avoided hill tops and had a relative cover
maximum in bottom flat areas. The early flowering T.su bte r r aneu m cv. had ahigher relative cover in
the drier hilltop soils. Although contingent, the gathered data supports both formulated hypothesis.
Improved pasture plants spatial segregation has, certainly, a fundamental role in the agronomic
superi-ority ofSBPPRL.
The improved clover species that don't burry seeds and grasses have clear persisting
difficul-ties inpastures subjected to intense and continuous grazing.
Keywords: grasses, legumes, T rif ol i um su bte r r a n eum, pasture ecology
Fi g u ra 1 . "B i p l o t" especies/cv.-variaveis e xp l a n a t o r i as de u ma R DA rea l i z a da c o m dad o s
f l o r i s t i c o s e a m b i e n t a is r e c o l h i d o s n u m a Pa st a g e m P e r m a n e n t e Sem e a da B i o d i ver s a R i ca e m
Le gu m i n osa s ( PPS B R L) n a Q u i n t a da Fran~a (C o v i l ha , P o r t u g a l ) .
Legenda: Pos fisi -variavel explanat6ria "Posi9ao Fisiognifica" (tres niveis: cimo-de-encosta
a
esquerda, fundo-de-encosta
a
direita). A i r cary - Ai r a ca ryo p hyl l e a , B r a b a r r - B r as s i ca b ar r el i e r i, Br o h o r d - B r o m u s h o r deaceu s, C a p b u r s - C a ps el l a b u rsa - p a s t o r i s , Cer b r a e - Cer a s t i u m b rac hypetal u m, Ch a m i x t - C h a m a e melu m m i x t um, Ch r s eg e - C h rys an t hem um seget um, D ac-D act yli s g l o m e r a t a, Er o cicu - E r o d i u m ci c uta r i u m , G a u d jrag - G a u d ini a j r agi lis, H o l l ana - H o l c us
l a n a t u s, Lol p e ren - L o l ium p e ren ne, O no n a t r - O n o n is s p i n osa, O r n c om p - O r n ithop u s c o m pre ss us,
Pia co r a - P l a n ta go c o ron o pus , P i a la ne - Pl a n t ago lanc eo l a t a, R u m ac et - R u m e x ac et o s a, S is o j j i
-S i.< ,ym b r i u m o f f ici n a l e, Tric er n - T r i f o l i u m c e r n u u m , T r i d u b i - T . d u b i u m, Trigl o m - T .g l o mer atu m,
T r i mic h - T.m i c hel i a n u m, T s C p+w o - T.s u bter r aneu m morfotipo 'Campeda' + 'Woogenellup', T s
goss - T . s ub te r r a n eu m morfotipo 'Gosse', Ts L o sa - T. subter r aneum morfotipo 'Losa', TsDen m
Figura 2. "Biplot" especies/cv. melhoradas - variliveis explanat6rias de uma RDA realizada com
dados floristicos e ambientais recolhidos numa PPSBRL na Quinta da Fran«;a (Covilha,