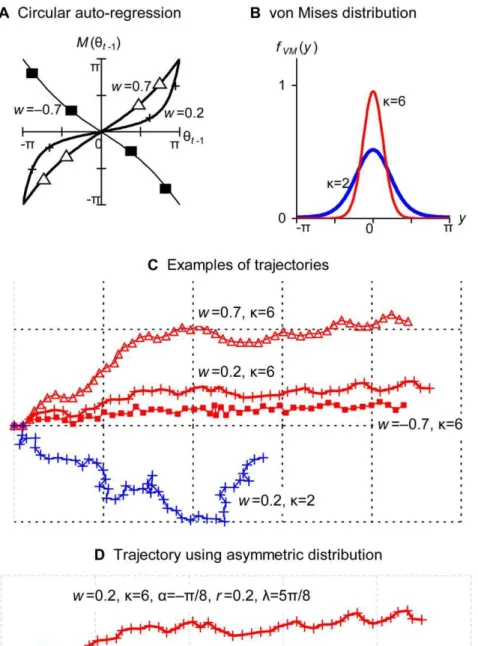
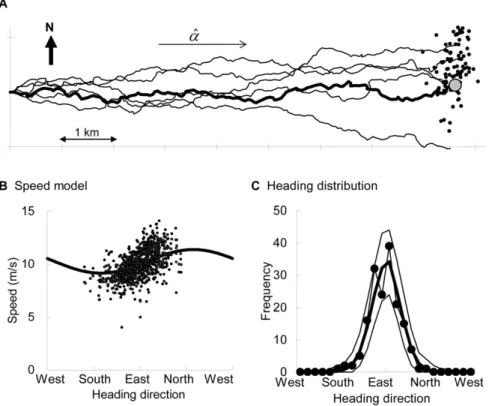
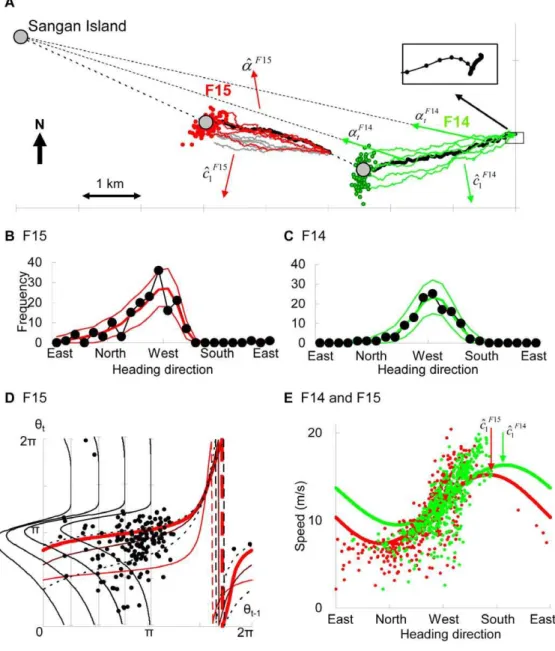
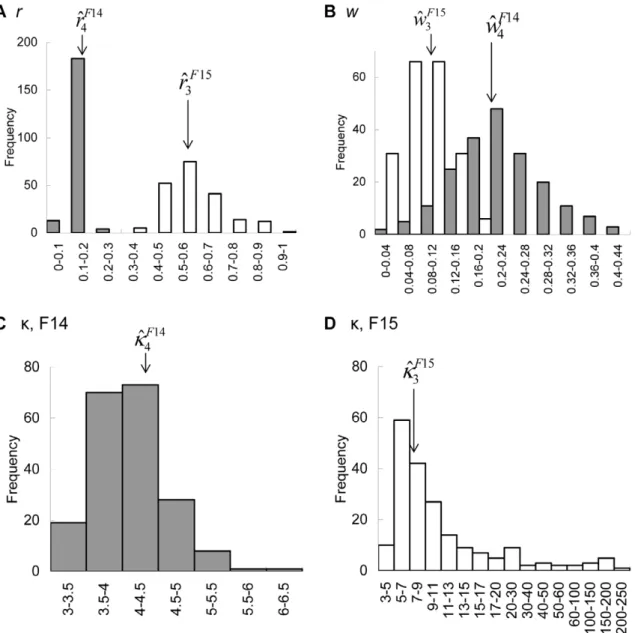
Toward the quantification of a conceptual framework for movement ecology using circular statistical modeling.
Texto
Imagem


Documentos relacionados
4.2 What kinds of environmental studies appear most likely to enable health officials to predict the nature and degree of future stresses?. 4.3 What are your present
How much knowledge do the national health authorities have about the sources of revenue for the sector and for its component institutions; of the purpose for which
After eleven sessions of intensive analysis of Official Document No, 85, the Executive Committee approved Resolutions III, IV, V, VI and XXI (Annexes I through V of this
That the Director shall be authorized to transfer credits between parts of the budget, provided that. such transfers of credits between parts as are made do not exceed 10 per cent
To instruct the Director to present to the X Meeting of the Directing Council the Proposed Program and Budget of the Pan American Sanitary Organization for the year 1958, as
To transmit to the Directing Council, as the Pro- posed Program and Budget of the Executive Committee, pur- suant to Chapter IV, Article 12-C, of the Constitution of the
Recently, Hassani-Gangaraj 7 proposed a inite element simulation to predict the kinetics of layer growth and nitrogen distribution during nitriding according the analogy
As for the methodological recommendations that the main representatives of the Latin American movement of history of ideas proposed as norms for the historiographical task
