Universidade do Minho
Universidade do Minho
Universidade do Minho
Universidade do Minho
E s c o l a d e C i ê n c i a s
Ricardo Filipe Duarte da Cruz
Indução
Indução
Indução
Indução e Recuperação do S
e Recuperação do S
e Recuperação do S
e Recuperação do Stresse
tresse
tresse
tresse Hídrico
Hídrico
Hídrico
Hídrico
em
em
em
em
Variedades Portuguesas de
Variedades Portuguesas de
Variedades Portuguesas de
Variedades Portuguesas de Milho
Milho
Milho
Milho
Setembro de 2006
Universidade do Minho
Universidade do Minho
Universidade do Minho
Universidade do Minho
E s c o l a d e C i ê n c i a s
Ricardo Filipe Duarte da Cruz
Indução
Indução
Indução
Indução e Recuperação do Stresse
e Recuperação do Stresse
e Recuperação do Stresse
e Recuperação do Stresse Hídrico
Hídrico
Hídrico
Hídrico
em
em
em
em
Variedades Portuguesas de Milho
Variedades Portuguesas de Milho
Variedades Portuguesas de Milho
Variedades Portuguesas de Milho
Tese de Mestrado
Biologia do Stresse em Plantas
Trabalho efectuado sob a orientação de
Professor Doutor Jorge Marques da Silva
Professor Doutor Jorge Marques da Silva
Professor Doutor Jorge Marques da Silva
Professor Doutor Jorge Marques da Silva
Prof
Prof
Prof
Professora Doutora Ana Cunha
essora Doutora Ana Cunha
essora Doutora Ana Cunha
essora Doutora Ana Cunha
Setembro de 2006
É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE COMPROMETE.
COMPROMETE. COMPROMETE. COMPROMETE.
“Have courage for the great sorrows of life and patience for the small ones; and when you have laboriously accomplished your daily task, go to sleep in peace. God is awake.” Victor Hugo (1802 Victor Hugo (1802 Victor Hugo (1802 Victor Hugo (1802 ---- 1885) 1885) 1885) 1885)
Agradecimentos
Agradecimentos
Agradecimentos
Agradecimentos
Agradecimentos
Ao Professor Jorge Marques Silva por me ter sugerido o tema deste trabalho, e me ter acolhido prontamente na Faculdade de Ciências da Universidade de Lisboa. Agradeço-lhe também a liberdade que me permitiu ao longo do trabalho e a discussão dos problemas que foram surgindo. Agradeço-lhe ainda a leitura cuidadosa e com espírito crítico do presente manuscrito.
À Professora Ana Cunha pela disponibilidade e pelo acompanhamento ao longo de todo o trabalho, em especial o realizado na Universidade do Minho. Agradeço-lhe também pela leitura cuidadosa e crítica do presente manuscrito.
Ao Professor Pedro Fevereiro e ao Engenheiro Silas Pego pelo apoio ao projecto e o auxílio na obtenção das sementes.
À Professora Anabela Bernardes da Silva pelo auxílio prestado na determinação das actividades da PEPC e EM-NADP.
À Professora Celeste Arrabaça pelo auxílio prestado na determinação das actividades da RuBisCO.
Ao Banco Português de Germoplasma Vegetal, Braga, pelas sementes das diferentes variedades portuguesas de milho.
À Ana Elisabete, à Ana Sofia, à Elsa Elias e ao André Costa pela ajuda imprescindível que me ofereceram ao longo de todo o trabalho, quer na utilização dos aparelhos, quer na metodologia aplicada. Também quero agradecer-lhes a amizade e o apoio ao longo do trabalho.
À Alexandra Marques pela discussão dos problemas que foram surgindo, pela amizade e apoio que demonstrou.
À Cátia Nunes pela ajuda preciosa na quantificação dos aminoácidos livres e prolina, na Universidade do Minho.
À funcionária Manuela Lucas pela ajuda preciosa, pela amizade e apoio que sempre demonstrou.
A todos os docentes da Secção de Fisiologia e Bioquímica Vegetais pelo ambiente e apoio que demonstraram ao longo do trabalho.
Agradeço à minha família, à Leonor e a todos os meus amigos pelo apoio nos momentos de maior dificuldade.
Resumo
Resumo
Resumo
Resumo
Resumo
Este trabalho teve como objectivo geral, na primeira fase, caracterizar as estratégias de resposta ao stresse hídrico, com ênfase no metabolismo fotossintético, em seis variedades portuguesas de milho (Zea mays L.) e, na segunda fase, aprofundar o conhecimento sobre o comportamento fisiológico das duas variedades com respostas mais contrastantes. Utilizaram-se dois sistemas de indução de stress, um rápido e um lento, e recorreu-Utilizaram-se, na primeira faUtilizaram-se, à medição das trocas gasosas através da análise de gases por radiação infra-vermelha e de fluorescência modulada da clorofila a. Nas duas variedades seleccionadas para a segunda fase, estudou-se adicionalmente a resposta do aparelho fotossintético ao stress hídrico sob diferentes regimes de PPFD e diferentes concentrações de CO2. Analisaram-se também as
actividades das principais enzimas do metabolismo fotossintético (RuBisCO, PEPC, EM-NADP) e a variação do teor em prolina.
Nos ensaios de indução lenta do stresse ocorreu uma diminuição de todos os parâmetros de trocas gasosas (A, E, gs, WUE) nas seis variedades. Destacaram-se as
variedades PB260 e PB269, como aquelas onde os parâmetros sofreram uma diminuição progressiva e mais lenta, e a variedade PB369, onde os efeitos negativos do stresse hídrico se fizeram sentir de forma mais abrupta, com uma diminuição rápida dos valores. As variedades recuperaram os valores iniciais da maioria dos parâmetros após irrigação, exceptuando-se a variedade PB369 cuja recuperação se deu de forma mais lenta. Nos ensaios de fluorescência da clorofila a, verificou-se uma diminuição de vários parâmetros (Fv/Fm, ΦPSII, qP, ETR) ao
longo do stresse lento e do stresse rápido. Por outro lado, qN aumentou em todas as variedades, à excepção de PB269, na qual não variou. Comparando os ensaios dos dois tipos de stresse, verificou-se que os parâmetros fisiológicos que diminuíram o fizeram de forma mais marcante no stresse rápido, em todas as variedades. As variedades PB269 e PB369 foram seleccionadas como sendo a mais e a menos tolerante ao stresse hídrico, respectivamente.
A partir das determinações dos potenciais hídricos constatou-se que a variedade PB269 conseguiu manter um potencial hídrico mais elevado do que a variedade PB369 em condições de stresse hídrico semelhantes e que possui uma parede celular mais elástica, permitindo a manutenção da turgescência até THR mais baixos apresentando-se esta como uma boa estratégia de tolerância ao stresse hídrico.
Através da decomposição do parâmetro de fluorescência qN, verificou-se que houve um aumento de qE com a intensificação do stresse hídrico, na variedade PB369. Por outro lado, na variedade PB269 houve a manutenção deste parâmetro até THR de cerca de 50%, indicando uma maior capacidade de manter activos os processos fotossintéticos sem recorrer a processos de amortecimento não-fotoquímicos.
Nos ensaios enzimáticos, observou-se que em ambas as variedades houve diminuição das actividades enzimáticas da RUBISCO, PEPC e EM-NADP com a diminuição do THR, nos ensaios de stresse lento. Contudo, nos ensaios de stresse rápido tal diminuição só aconteceu na RUBISCO, mantendo-se as actividades das outras enzimas constantes.
Os dados parecem indicar que ambas as variedades possuem uma reserva de prolina que poderá ter sido rapidamente mobilizada no período de recuperação do stresse, principalmente na variedade PB269.
No seu conjunto, os resultados obtidos parecem mostrar que existe um espectro alargado de respostas fisiológicas face ao stresse hídrico que, a ter uma base genética de elevada hereditabilidade e estabilidade, poderão constituir uma base de selecção em futuros programas de melhoramento.
Abstract
Induction and Recovery of Water Stress in
Induction and Recovery of Water Stress in
Induction and Recovery of Water Stress in
Induction and Recovery of Water Stress in
Portuguese Maize Cultivars
Portuguese Maize Cultivars
Portuguese Maize Cultivars
Portuguese Maize Cultivars
Abstract
Abstract
Abstract
Abstract
This work aims for the identification of several aspects of water stress physiological response strategies in different Portuguese maize cultivars (Zea mays L.), in a first stage, and expand the knowledge of the most tolerant and less tolerant cultivars physiological responses to water stress. The responses were studied under two water stresse regimes – slow stress and rapid stress – relevant in field conditions, and in the recovery process after irrigation. In the first part of the work, a preliminary characterization of six cultivars was made measuring gas exchange with an infrared gas analyser and chlorophyll a fluorescence. In the second part of the work, a deeper characterization of those two cultivars was made through the study of water relations and photosynthetic metabolism under different PPFD and external CO2
conditions, the enzymatic activities (RuBisCO, PEPC, NADP-ME) and proline content variation.
In slow stress, all the parameters of gas exchange (A, E, gs, WUE) suffered a reduction
in all six cultivars. In PB260 and PB269, the reduction was made in a slower progressive way, and, in contrast, in PB369 the parameters declined abruptly. In general, the parameters recovered upon irrigation, with the exception of PB369, in which the recovery was slower. In rapid stress, the parameters responded in a similar but very fast way. PB369 was the only cultivar with a slower response. Chlorophyll fluorescence parameters (Fv/Fm, ΦPSII, qP, ETR)
suffered a decrease, in slow and rapid stress. On the other hand, qN increased in all cultivars, with the exception of PB269, in which it maintained its values. Comparing slow and rapid stress, the parameters decreased faster in rapid stress, in all cultivars. PB269 and PB369 were selected as the cultivars more and less tolerant to water stress.
PB269 is able to maintain higher leaf water potential under water stress and as a more elastic cell wall that allows it to maintain turgescence at lower RWC, resulting in a good tolerance strategy to water stress.
The decomposition of the qN fluorescence parameter, showed a rise of the qE component, in PB369, with increased water stress. PB269 maintained this parameter to a RWC of 50%, indicating a suitable photosynthesis without activating major non-photochemical quenching processes.
On the enzymatic assays, there was a decrease in the activities of RUBISCO, PEPC and NADP-ME, with RWC decrease, in slow stress. However, in rapid stress this decrease only occurred in RUBISCO.
Both cultivars appear to have a proline reserve that could be easily mobilised during recovery, especially in PB269.
The data presented in this work appears to indicate the existence of a wide spectrum of physiological responses to water stress that, having a highly hereditable and stable genetic background, can establish the basis of future plant breeding programs.
Índice
Índice
Índice
Índice
Índice
Agradecimentos ……….. I Resumo ……….……... III Abstract ……….…….... V Índice ………...………... VII Símbolos e Abreviaturas ……….... IX p1. Objectivos e Abordagem do Problema ……… 1
p 2. Introdução Geral ... 3
2.1. Descrição Botânica do Milho .………..………...….……….…. 3
2.2. O Milho na Economia Mundial e em Portugal ………....…..……….……….…… 4
2.3. Relações Hídricas em Plantas ……….. 4
2.4. Stresse Hídrico nas Plantas ……….. 6
2.4.1. Aspectos Gerais ………...……….. 6
2.4.2. Respostas Estomáticas ao Stresse Hídrico ……….... 8
2.4.3. Metabolismo do Carbono e Efeitos do Stresse Hídrico na Fotossíntese ………...… 9
2.5. Estratégias de Sobrevivência à Seca ……….. 14
2.5.1. Aspectos Gerais ……...……….... 14
2.5.2. Modificações Anatómicas, Morfológicas e Metabólicas na Resposta ao Stresse Hídrico ……….…. 15
2.5.3. Solutos Compatíveis e Ajustamento Osmótico ………... 19
2.6. Técnicas de Melhoramento e Selecção para Tolerância ao Stresse Hídrico ………...……...………… 24
p 3. Metodologia Geral ………..……. 29
3.1. Material Vegetal ………. 29
3.2. Indução do Stresse Hídrico ………..……….. 30
3.3. Relações Hídricas ………...… 30
3.3.1. Determinação do Teor Hídrico Relativo da Folha ……….………… 30
3.3.2. Determinação do Teor Hídrico do Solo ……….. 31
3.4. Medição das Trocas Gasosas por Análise de Gases por Radiação Infra-Vermelha (IRGA) …...………….. 31
3.5. Medição da Fluorescência Modulada da Clorofila a …………...………...………..…………. 33
3.6. Medição da Libertação de O2 ………. 36
3.7. Medição de Actividades Enzimáticas ……….... 37
3.8. Análise Estatística ……….. 37
p 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho ………. 39
4.1. Introdução ………... 39
4.2. Metodologia ………... 39
4.2.1. Material Vegetal ………. 39
4.2.2. Relações Hídricas ……… 39
4.2.3. Medição das Trocas Gasosas ……….. 39
4.2.4. Medição da Fluorescência Modulada da Clorofila a ………..… 41
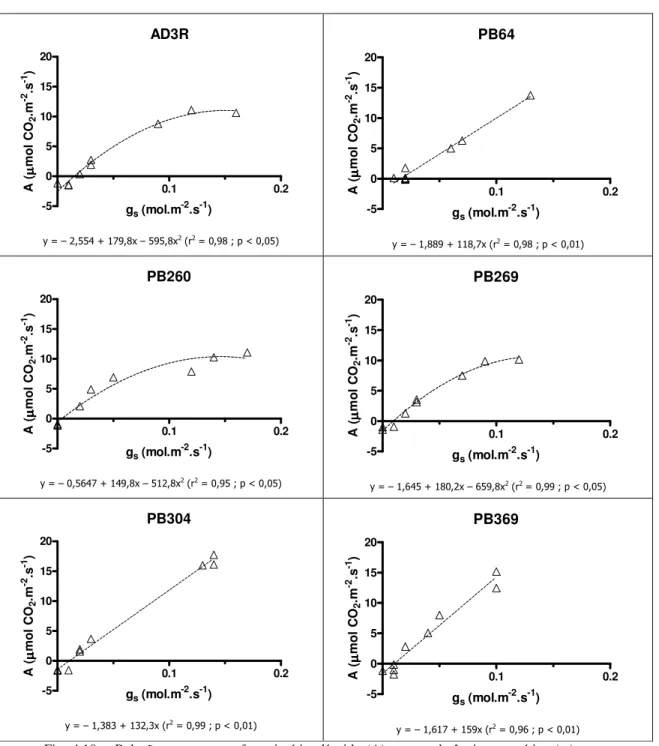
4.3. Resultados e Discussão ………..… 42
4.3.1. Relações Hídricas ……….... 42
4.3.2. Trocas Gasosas ………...…. 47
4.3.2.1. Stresse Lento ……….... 48
4.3.2.2. Stresse Rápido ……….. 59
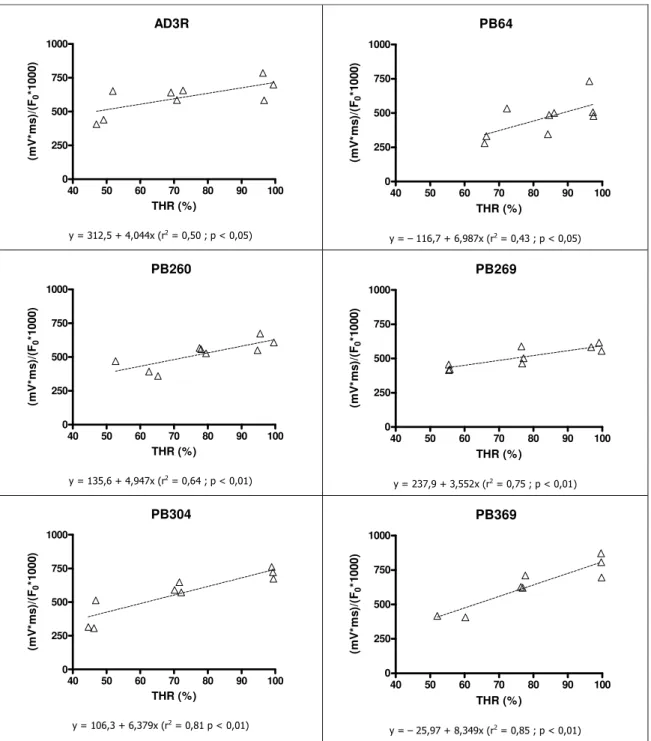
4.3.3. Fluorescência Modulada da Clorofila a ……….. 68
4.3.3.1. Stresse Lento ……….... 68
4.3.3.2. Stresse Rápido ……….…. 75
4.4. Conclusões ………...……….. 83
4.4.1. Conclusões das Trocas Gasosas ………...………... 83
4.4.2. Conclusões da Fluorescência Modulada da Clorofila a ………...………... 83
4.4.3. Conclusões da Fase I ……….……….. 84
Índice
5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico …... 85
5.1. Introdução ………..…. 85
5.2. Metodologia ………... 85
5.2.1. Material Vegetal ……….. 85
5.2.2. Relações Hídricas ……….... 85
5.2.2.1. Determinação do Teor Hídrico Relativo da Folha ……….……….. 85
5.2.2.2. Determinação do Potencial Hídrico das Folhas ………... 86
5.2.2.3. Estabelecimento de Curvas Pressão/Volume ………... 86
5.2.3. Medição das Trocas Gasosas ………...… 87
5.2.3.1. Curvas A/PPFD ……… 87
5.2.3.2. Curvas A/CO2a ……….. 87
5.2.3.3. Determinação da Libertação de O2 ………...………… 88
5.2.4. Medição da Fluorescência Modulada da Clorofila a ………..… 88
5.2.5. Determinação de Clorofilas e Carotenóides ……… 89
5.2.6. Determinação de Actividades Enzimáticas ………. 89
5.2.6.1. Extracção ……….. 90
5.2.6.2. Ribulose-1,5-Bisfosfato Carboxilase/Oxigenase (RuBisCO) ………..… 90
5.2.6.3. Fosfoenolpiruvato Carboxilase (PEPC) ………...… 91
5.2.6.4. Enzima Málica Dependente de NADP (EM-NADP) …...……….... 91
5.2.7. Quantificação de Proteína Solúvel ……….…. 91
5.2.8. Quantificação de Solutos Compatíveis ………... 92
5.2.8.1. Extracção ……….. 92
5.2.8.2. Quantificação de Aminoácidos Livres e Prolina ……….. 92
5.3. Resultados e Discussão ……….. 93 5.3.1. Relações Hídricas ……….... 93 5.3.2. Trocas Gasosas ………...…. 96 5.3.2.1. Curvas A/PPFD ……… 96 5.3.2.2. Curvas A/CO2a ………. 105 5.3.2.3. Libertação de O2 ………. 112
5.3.3. Fluorescência Modulada da Clorofila a ……….... 114
5.3.4. Clorofilas Totais e Carotenóides ………... 125
5.3.5. Metabolismo do Carbono ……… 128
5.3.5.1. Ribulose-1,5-Bisfosfato Carboxilase/Oxigenase (RuBisCO) ……… 129
5.3.5.2. Fosfoenolpiruvato Carboxilase (PEPC) ………. 133
5.3.5.3. Enzima Málica Dependente de NADP (EM-NADP) ……...……….. 136
5.3.6. Solutos Compatíveis ………..… 137 5.3.6.1. Aminoácidos Livres ……… 138 5.3.6.2. Prolina ………. 140 5.3.7. Conclusões da Fase II ……….... 142 p 6. Conclusões Gerais ………. 145 p 7. Referências Bibliográficas ……….... 147 p
Símbolos e Abreviaturas
Símbolos e Abreviaturas
Símbolos e Abreviaturas
Símbolos e Abreviaturas
Símbolos e Abreviaturas
A taxa fotossintéticaA/CO2a taxa fotossintética em função da concentração externa de CO2
A/PPFD taxa fotossintética em função da densidade de fluxo fotónico fotossintético
ABA ácido abcísico
Apot potencial fotossintético
ATP adenosina trisfosfato
ATPase ATP sintase
C3 metabolismo fotossintético cujo primeiro composto estável formado possui 3
átomos de carbono
C4 metabolismo fotossintético cujo primeiro composto estável formado possui 4
átomos de carbono
Ca clorofila a
Ca2+ ião cálcio
Cb clorofila b
Ca+b clorofila total
CA1P 2-carboxiarabinitol-1-fosfato Ci concentração interna de CO2 CO2 dióxido de carbono CO2a concentração externa de CO2 Cx+c carotenóides totais DMSO dimetilsulfóxido DTT ditiotritol E taxa transpiratória
EDTA ácido etilenodiamina tetracético
EM-NADP enzima málica dependente de NADP
Eq. equação
ETR taxa de transporte de electrões no PSII
Fm fluorescência máxima
Fm’ fluorescência máxima medida à luz
Fo fluorescência basal
Fo’ fluorescência basal medida após iluminação
Símbolos e Abreviaturas Fig. figura gs condutância estomática H2O água H2O2 peróxido de hidrogénio HCl ácido clorídrico
HEPES ácido N-(2-hidroxietil) piperazina N’-2-etanosulfónico
IRGA infra-red gas analyser (analisador de gases por radiação infra-vermelha)
K+ ião potássio
Kcat constante de catálise
KCl cloreto de potássio
KOH hidróxido de potássio
LHCII complexo antena do PSII
M molar
MDH malato desidrogenase
MPa mega Pascal
Mg2+ ião magnésio
MgCl2 cloreto de magnésio
mV milivolt
N2 azoto molecular
NADH nicotinamida adenina dinucleótido reduzida
NADP fosfato de nicotinamida adenina dinucleótido
NADP+ fosfato de nicotinamida adenina dinucleótido oxidado
NADPH fosfato de nicotinamida adenina dinucleótido reduzido
NaHCO3 hidrogenocarbonato de sódio
NaH14CO
3 hidrogenocarbonato de sódio marcado radioactivamente
NPQ amortecimento não-fotoquímico
O2 oxigénio molecular
p probabilidade
P5CS ∆1-pirrolina-5-carboxilato sintetase
PAM pulse amplitude modulated (amplitude de pulso modulada) PDH prolina desidrogenase
PEG polietilenoglicol
PEP fosfoenolpiruvato
Símbolos e Abreviaturas
Pf peso fresco
pH logaritmo do inverso da concentração hidrogeniónica
PPFD densidade de fluxo fotónico fotossintético
ppm partes por milhão
PRK fosforibulocinase Ps peso seco PSII fotossistema II Pt peso túrgido PVP polivinilpirrolidona QA quinona A QB quinona B
QAC composto quaternário de amónio
qE coeficiente de amortecimento do estado de alta energia
qI coeficiente de amortecimento fotoinibitório
qN coeficiente de amortecimento não-fotoquímico
qP coeficiente de amortecimento fotoquímico
qT coeficiente de amortecimento de transição de estado
QTL quantitative trait loci (loci de características quantitativas) r2 coeficiente de determinação
rbcS subunidade menor da RUBISCO
ROS reactive oxygen species (espécies reactivas de oxigénio)
RuBisCO ribulose-1,5-bisfosfato carboxilase/oxigenase
RuBP ribulose-1,5-bisfosfato
SLA área foliar específica
SnCl2.H2O cloreto de estanho hidratado
THR teor hídrico relativo
THS teor hídrico do solo
V volt
WUE eficiência do uso da água Φ
Φ Φ
ΦPSII rendimento quântico efectivo do PSII
Ψ Ψ Ψ Ψg potencial gravítico Ψ Ψ Ψ Ψp potencial de pressão Ψ Ψ Ψ Ψs potencial osmótico Ψ Ψ Ψ Ψw potencial hídrico
1. Objectivos e Abordagem do Problema
1. Objectivos e Abordagem do Problema
1. Objectivos e Abordagem do Problema
1. Objectivos e Abordagem do Problema
1. Objectivos e Abordagem do Problema
O milho (Zea mays L.) é originário da América Central, tendo sido rapidamente distribuído pelos quatro cantos do Mundo através das viagens marítimas do século XVI. Ao longo de gerações, a agricultura produziu numerosas variedades locais de milho. Actualmente, estas variedades voltam a adquirir elevada importância, mas num contexto de valorização dos produtos regionais. A diversidade genética das variedades portuguesas de milho constitui um recurso valioso para o melhoramento da espécie e, além disso, a comparação de variedades com diferentes capacidades de resistência ao stresse pode fornecer informações importantes sobre os mecanismos fisiológicos que a conferem.
O stresse hídrico é a mais importante limitação à produtividade vegetal. O desenvolvimento de variedades resistentes à seca assume particular importância no panorama de alterações climáticas, em que o aumento da temperatura prolonga o período de restrições hídricas das plantas (Bruce et al., 2002; Taiz e Zeiger, 2002; Chaves et al., 2002, 2003). Para contrariar a influência destas alterações na produtividade, deverá ser desenvolvido um programa de selecção para uma maior tolerância ao stresse hídrico, mantendo uma boa eficiência do uso da água e da capacidade fotossintética, de modo a garantir um aumento de produção nestas condições.
Para melhor compreender as limitações fisiológicas e bioquímicas da fotossíntese, torna-se necessário elucidar aspectos fundamentais do metabolismo fotossintético e da gestão da água ao nível celular e da planta inteira e, assim, avançar no conhecimento dos mecanismos de resposta ao stresse.
A tolerância é o resultado duma resposta integrada, determinada por múltiplos genes. Deste modo, a identificação de um elevado número de alterações de valor adaptativo reveste-se de enorme importância para o planeamento de estratégias de melhoramento da resistência à seca de espécies de interesse agrossilvícola.
Existem ainda lacunas de conhecimento sobre os mecanismos que afectam a fisiologia e o metabolismo em condições de stresse hídrico, nomeadamente sobre a contribuição relativa dos processos estomáticos e não-estomáticos (ou metabólicos) na redução da fotossíntese (Lawlor e Cornic, 2002). Sob stresse moderado parecem predominar as limitações estomáticas, mas, em certas circunstâncias, à medida que o stresse se intensifica, as limitações não-estomáticas (como a alteração da actividade e estrutura das enzimas do metabolismo fotossintético do carbono) ganham peso (Marques da Silva e Arrabaça, 2004a). A extensão de cada variação, contudo, é desconhecida. Foi sugerido ser possível incluir diferentes espécies
1. Objectivos e Abordagem do Problema em dois grupos com respostas ao stresse distintas: nas plantas do tipo I, as limitações ao nível do mesófilo aumentam linearmente com a intensidade do stresse, enquanto que, nas plantas do tipo II, não há limitações do mesófilo até um determinado nível de stresse hídrico, mas, uma vez atingido o limiar, observa-se um rápido aumento dessas limitações (Lawlor e Cornic, 2002). Não se conhecem as razões porque uma espécie responde da forma I ou II e se comporta de forma distinta, em diferentes circunstâncias, admitindo-se, no entanto, que a velocidade de imposição do stresse poderá ser relevante. Deste modo, também a forma como cada variedade responde ao stresse é determinante para seleccionar a estratégia de melhoramento.
Uma resposta quase universal ao défice hídrico é a acumulação de solutos osmoticamente compatíveis com o metabolismo celular (Bohnert e Shen, 1999). Esta resposta pode conduzir a um ajustamento osmótico e à manutenção de um estado hídrico favorável para o metabolismo e, neste caso, os solutos denominam-se osmorreguladores. Noutros casos, a síntese de certos solutos compatíveis pode ter um papel fundamental na protecção de proteínas (nomeadamente enzimas), na estabilização de membranas celulares e na sequestração de espécies reactivas de oxigénio (ROS, do inglês reactive oxygen species) (Bohnert e Shen, 1999), cuja produção aumenta em condições de défice hídrico (Iturbe-Ormaetxe et al., 1998). Neste caso, designam-se os solutos de osmoprotectores. Vários estudos revelam que a produção de solutos compatíveis permitiu uma maior actividade de enzimas do ciclo de Calvin em condições de défice hídrico (Shen et al., 1997b; Heuer, 2003).
Dada a importância de conhecer a influência do stresse hídrico no desenvolvimento, crescimento e produtividade das plantas agrícolas, tornou-se necessário o desenvolvimento e aplicação de técnicas que forneçam parâmetros com os quais seja possível avaliar os efeitos do stresse ambiental. Algumas dessas metodologias serão aqui abordadas, nomeadamente a medição da fluorescência da clorofila a e das trocas gasosas por análise de gases na banda do infra-vermelho (IRGA).
Neste trabalho, proceder-se-á à análise de seis variedades portuguesas de milho através destas técnicas, isolando as duas que mais se distinguem, positiva e negativamente, em termos de tolerância ao stresse hídrico. Ulteriormente, a investigação seguirá novas abordagens dessas duas variedades, de modo a identificar características que permitam explicar algumas diferenças entre a variedade mais tolerante e a variedade mais susceptível ao stresse hídrico, nomeadamente actividades de enzimas-chave do metabolismo do carbono e quantificação de solutos compatíveis, como a prolina.
2. Introdução Geral
2
2
2
2.... Introdução Geral
Introdução Geral
Introdução Geral
Introdução Geral
2.1 2.1 2.1
2.1.... Descrição Botânica Descrição Botânica Descrição Botânica do Milho Descrição Botânica do Milho do Milho do Milho
Durante séculos os investigadores interrogaram-se sobre a origem geográfica do milho. Actualmente, todos estão de acordo que a região de origem desta espécie é o Novo Mundo, seja no México, na América Central ou na América do Sul.
A espécie de milho que hoje utilizamos é incapaz de sobreviver sem a intervenção do Homem, uma vez que não possui mecanismos de disseminação, nem órgãos de resistência. É uma espécie relativamente recente, com alguns milhares de anos, tendo surgido provavelmente a partir do período em que o Homem iniciou o desenvolvimento da agricultura.
O milho pertence à Divisão Magnoliophyta, Classe Liliopsida, Ordem Poales, e Família das Poaceae (também conhecida como Gramineae).
É uma espécie muito polimórfica, apresentando populações muito precoces, em que a maturação é atingida em 60-70 dias, e populações muito tardias, que exigem 10 a 11 meses. Também o número de folhas por planta pode variar entre 8 e 48, a altura do caule entre 60 centímetros e 6 metros e a dimensão da espiga entre 2,5 e 30 centímetros (Floyd et al., 2002).
Embora existam grandes diferenças morfológicas entre as populações de milho, todas as variedades cultivadas têm características morfológicas comuns, nomeadamente os órgãos vegetativos (raízes, caule, folhas) e os órgãos reprodutores (panícula e espigas).
No milho, existem dois sistemas radiculares: um seminal ou provisório e um coronário ou definitivo. As raízes seminais são as primeiras a formar-se permitindo à planta desenvolver-se até formar as raízes coronárias, as quais permitem também fixar a planta ao solo (Floyd et al., 2002).
O caule do milho é formado por nós e entrenós, variando o número de nós entre 8 e 48. Ao nível de cada nó estão inseridos uma folha e um gomo axilar. Quando os gomos axilares da base do caule se desenvolvem dão origem a rebentos que, tal como o pé principal, também são formados de caule, folhas e inflorescências (Floyd et al., 2002).
Num pé de milho as folhas apresentam dimensões diferentes. As folhas da base são pequenas. O engrossamento do caule provoca a ruptura dos vasos que as alimentam fazendo com que elas deixem de ser funcionais. As folhas inseridas na zona mediana do caule (ao nível da inserção das espigas) são as mais longas e mais largas. No terço superior do caule, as
2. Introdução Geral folhas vão diminuindo de dimensão à medida que a sua inserção se aproxima da panícula (Floyd et al., 2002).
2.2. 2.2. 2.2.
2.2. O Milho na Economia MO Milho na Economia MO Milho na Economia MO Milho na Economia Mundial e em Portugalundial e em Portugalundial e em Portugal undial e em Portugal
Actualmente, o milho é cultivado em diversos pontos do planeta, desde países como Canadá e Rússia até ao Brasil e países africanos; desde locais situados abaixo do nível do mar junto do Mar Cáspio até aos 3600 metros de altitude nos Andes peruanos; desde zonas semi-áridas onde a precipitação anual não ultrapassa 250 milímetros até às regiões onde ela ultrapassa os 1000 milímetros; em regiões com Verão muito curto (Canadá) e nas de Verão permanente (Equador). Nenhuma outra cultura tem uma distribuição geográfica tão vasta (CIMMYT, 2001).
Depois do trigo e do arroz, o milho é o cereal mais cultivado, com uma produção de mais de 400 milhões de toneladas por ano, sendo os EUA o maior produtor mundial, com cerca de 40% da produção (CIMMYT, 2001).
Em Portugal, a área cultivada é de cerca de 180.000 hectares. Os milhos mais cultivados são os híbridos, calculando-se que ocupem cerca de 71,4 % da área global da cultura. A cultura do milho decorre no período da Primavera-Verão, quando as temperaturas são mais elevadas, sendo a temperatura óptima para o crescimento do milho de 24 a 30ºC (Syngenta, 2004). Este facto levanta sérios problemas ao cultivo de milho, dado que estas temperaturas, associadas ao aumento da evapotranspiração e à diminuição dos recursos hídricos no solo, limitam a água disponível para a planta durante o seu desenvolvimento o que é determinante para garantir uma boa produtividade.
2.3 2.3 2.3
2.3. Relações Hídricas em P. Relações Hídricas em P. Relações Hídricas em P. Relações Hídricas em Plantaslantaslantas lantas
A água é o líquido mais abundante à face da Terra, tendo uma função muito importante nas plantas como responsável pela dissolução e transporte de iões e moléculas e influenciando também a estrutura de proteínas, ácidos nucleicos, polissacáridos e outros constituintes da membrana celular. A água forma o ambiente no qual ocorre a maioria das reacções celulares, e participa directamente em muitas reacções bioquímicas (Taiz e Zeiger, 2002).
As células vegetais, quando completamente desenvolvidas, possuem um enorme vacúolo constituído de uma solução aquosa de natureza diversa, restando apenas 5 a 10% do
2. Introdução Geral volume celular para o citoplasma. A água constitui cerca de 80 a 95% da massa dos tecidos vegetais em crescimento. Por outro lado, estruturas como a madeira, constituída essencialmente por células mortas, contém quantidades baixas de água. As sementes possuem 5 a 15% de água, estando entre os tecidos vegetais com menor quantidade de água, embora na germinação necessitem de absorver enormes quantidades (Taiz e Zeiger, 2002).
As plantas estão continuamente a absorver e a perder água, grande parte dela perdida por transpiração foliar, através dos estomas, à medida que o CO2, necessário para a
fotossíntese, é absorvido da atmosfera.
O fluxo de água também é um meio importante de adquirir os minerais existentes no solo através da absorção nas raízes. O movimento de água é conduzido por difusão simples ou fluxo de massa, ou através de uma combinação destes dois mecanismos de transporte, em resposta a uma diferença de potencial hídrico (Steudle e Peterson, 1998; Steudle, 2000). A difusão acontece devido ao constante movimento térmico das moléculas, que ocorre na presença de um gradiente de concentrações. Por outro lado, existe o movimento em fluxo de massa, ao longo do contínuo solo-planta-atmosfera (Wei et al., 1999; Lawlor, 2002). Actualmente, a manutenção deste fluxo é explicada por dois modelos complementares, o modelo da pressão radicular, segundo o qual a água ascende na planta devido a uma pressão provocada pela entrada de água nas raízes, e o modelo de tensão-coesão-adesão, segundo o qual a coluna de água ascende devido às propriedades específicas das moléculas de água, referidas no nome do modelo (Kramer e Boyer, 1995).
A água é absorvida pelas raízes e o seu movimento pode ocorrer através de duas vias possíveis: apoplástica (ao longo dos espaços intercelulares) e simplástica (atravessando paredes celulares e membranas plasmáticas e depois difundindo-se pelos plasmodesmos) (Steudle e Peterson, 1998). No caso das células, designa-se por osmose a difusão de água através de membranas, dependendo de um gradiente da energia livre da água através da membrana, normalmente medido como diferença de potencial hídrico (Taiz e Zeiger, 2002).
A concentração de solutos no citosol e vacúolo e a pressão hidrostática exercida pela membrana plasmática na parede celular são os dois factores principais que afectam o potencial hídrico, embora, à medida que a distância vertical aumenta, a gravidade também possa desempenhar um papel importante. Os componentes do potencial hídrico (Ψw) podem
ser resumidos na expressão Ψw = Ψs + Ψp + Ψg, onde Ψs corresponde ao potencial osmótico
(efeito dos solutos na energia livre da água), Ψp corresponde à pressão hidrostática ou de
2. Introdução Geral maiores dimensões aumenta cerca de 0,1 MPa / 10 m) (Taiz e Zeiger, 2002). O potencial hídrico é normalmente um bom indicador do estado hídrico da planta.
Ao contrário das células animais, as células vegetais podem suportar uma elevada pressão intracelular, denominada pressão de turgescência, resultante da presença da parede celular. Esta pressão é necessária para diversos processos fisiológicos, nomeadamente a elongação celular, as trocas gasosas nas folhas, o transporte floémico, e diversos processos de transporte membranar. A pressão de turgescência também contribui para a manutenção da rigidez e da estabilidade mecânica em tecidos vegetais não-lenhificados.
2.4 2.4 2.4
2.4. Stresse . Stresse . Stresse . Stresse HHHHídrico nídrico nídrico nídrico nas Pas Pas Pas Plantaslantaslantaslantas
2.4 2.4 2.4
2.4....1. 1. 1. 1. Aspectos GeraisAspectos GeraisAspectos GeraisAspectos Gerais
As plantas estão frequentemente expostas a stresses ambientais que podem resultar em défice hídrico, toxicidade por cálcio e outros metais, deficiência em iões e fotoinibição, entre outras.
A reduzida disponibilidade de água é um dos factores ambientais mais importantes na regulação do crescimento e desenvolvimento das plantas e uma das principais causas para a redução do rendimento das colheitas que afectam a maioria das zonas cultivadas por todo o mundo. À medida que os recursos hídricos para utilização agronómica se vão tornando cada vez mais limitantes, o desenvolvimento de linhas de plantas tolerantes à seca tem surgido como um objectivo importante a atingir (Stoop et al., 1996; Hare et al., 1999; Jiang e Zhang, 2002; Bruce et al., 2002).
Durante o ciclo de vida de uma planta ocorrem frequentemente períodos de défice hídrico nos solos e/ou na atmosfera, mesmo fora de regiões áridas/semi-áridas, como foi documentado nas florestas decíduas temperadas (Burghardt e Riederer, 2003) ou nas florestas tropicais (Asner et al., 2004).
Na natureza, as plantas podem estar sujeitas a dois tipos de stresse hídrico de acordo com a sua duração, podendo ser de longo prazo (de dias a semanas ou meses) ou curto prazo (de horas a dias) (Chaves et al., 2003). As respostas das plantas à escassez de água são complexas, envolvendo alterações adaptativas e/ou efeitos deletérios. Em condições naturais, estas respostas podem ser sinergistica ou antagonisticamente modificadas por sobreposição de outros stresses, como é o caso do stresse salino (Xiong et al., 2002).
2. Introdução Geral As plantas terrestres também podem estar sujeitas à desidratação durante a exposição a temperaturas baixas (Hare et al., 1999), uma vez que a água pode não estar disponível na forma líquida para ser absorvida pelas raízes e, também, devido à diminuição da absorção radicular, nestas condições, mesmo quando está disponível.
Algumas das diferenças entre espécies, no que respeita ao crescimento e sobrevivência, podem ser atribuídas a diferentes capacidades para a aquisição de água, transporte e acumulação, e não a diferenças drásticas do metabolismo num determinado estado hídrico. No entanto, a regulação do metabolismo fotossintético também está dependente de factores como as limitações da difusão de CO2 na folha, repartição na alocação
de carbono para órgãos não-fotossintéticos e moléculas de defesa, ou mudanças na bioquímica da folha, que podem ser afectados pelo stresse hídrico (Chaves et al., 2002).
Em condições de défice hídrico, também ocorre o aumento da produção de radicais livres de oxigénio nas plantas, e a acumulação de solutos compatíveis poderá ter algum efeito protector contra danos oxidativos nas proteínas, interagindo com os grupos que as constituem (Iturbe-Ormaetxe et al., 1998).
As respostas iniciais ao stresse hídrico permitem a sobrevivência imediata. No entanto, para que a planta seja capaz de suportar o stresse imposto é necessária uma aclimatação, a qual produz novas capacidades metabólicas e/ou estruturais mediadas pela alteração da expressão génica (Sharp et al., 2004).
As respostas fisiológicas ao stresse ambiental têm que ser activadas antes de ocorrerem danos significativos nos tecidos afectados (Hare et al., 1999). Contudo, é a resposta integrada ao nível da planta inteira, incluindo a assimilação de carbono e a alocação de fotoassimilados para diferentes partes da planta e a capacidade reprodutora, que vai determinar a sobrevivência e persistência em condições de stresse ambiental (Chaves e Oliveira, 2004).
Para providenciar uma base sólida às estratégias de melhoramento da produção vegetal é necessário uma melhor caracterização dos receptores moleculares envolvidos na percepção do stresse e dos eventos moleculares que especificam a expressão da tolerância ao stresse (Hare et al., 1999).
Os grandes avanços na área da genómica permitiram um enorme progresso na caracterização de genes individuais e das suas respostas específicas ao stresse, de modo a determinar a cascata de eventos que estarão associados a vias de sinais inter- e intracelulares do défice hídrico, como é o caso do ácido abcísico (ABA) (Griffiths e Parry, 2002). Contudo, como a maioria dos esforços são dirigidos para a transformação genética das plantas (Casas et
2. Introdução Geral al., 1993), as abordagens fisiológicas tradicionais são negligenciadas. Até agora, foram desenvolvidas poucas variedades com maior tolerância ao stresse ambiental. Contudo, no mesmo período de tempo, o conhecimento das respostas fisiológicas aumentou substancialmente (Heuer, 2003).
2 2 2
2.4.4.4.4.2.2.2.2. Respostas . Respostas Estomáticas. Respostas . Respostas EstomáticasEstomáticasEstomáticas ao Stresse H ao Stresse H ao Stresse Hídrico ao Stresse Hídricoídricoídrico
Os estomas ocupam um papel central nas trocas gasosas, funcionando como o controlo a curto prazo da perda de água por transpiração e da assimilação do CO2 pelas plantas (Jones,
1998).
Foram identificadas duas respostas gerais (Tipo I e Tipo II) da fotossíntese ao stresse hídrico, que se distinguem pelo conteúdo relativo de água foliar abaixo do qual surgem os efeitos não-estomáticos (metabólicos) (Lawlor, 2002). Contudo, durante a imposição gradual de défices hídricos moderados (normalmente até –1,8 MPa durante um período de 15 dias ou mais), as respostas baseiam-se quase exclusivamente na limitação da condutividade estomática (gs) (Cornic e Fresneau, 2002; Yordanov et al., 2003).
Quando os défices hídricos se desenvolvem, um dos primeiros acontecimentos a tomar lugar nas plantas é o fecho estomático, em resposta a um declínio da turgescência da folha e/ou do seu potencial hídrico (Yordanov et al., 2003), ou ainda em resposta ao aumento da concentração de ABA no fluxo transpiratório (Sharp e LeNoble, 2002; Chaves et al., 2003). Este encerramento permite evitar a desidratação da planta e fenómenos de cavitação, os quais poderiam comprometer a sobrevivência das plantas. A diminuição do CO2 intracelular, após o
fecho estomático, e a consequente baixa eficiência do uso da luz pode induzir, a longo prazo, uma regulação da maquinaria fotossintética, no sentido de se adequar ao carbono disponível (Chaves et al., 2002).
À medida que a seca progride, nas plantas cultivadas no campo, o fecho estomático ocorre por períodos cada vez mais longos, durante o dia, tendo início a meio da manhã (Tallman, 2004). Esta diminuição da condutância estomática tem como efeito simultâneo a redução da assimilação de carbono e das perdas de água no período de maior défice de pressão de vapor entre a folha e a atmosfera, e leva a uma optimização da assimilação de carbono, em relação à disponibilidade de água (Jones, 1998).
Foi encontrada claramente uma dependência do ciclo diurno no que respeita à resposta estomática, tendo em conta a humidade do ar e ao estado hídrico da folha numa dada hora
2. Introdução Geral (Tallman, 2004), sugerindo que algumas das alterações diurnas na função estomática podem resultar de processos metabólicos com um ritmo circadiano.
2.4 2.4 2.4
2.4....3333. . . . Metabolismo do Carbono e Metabolismo do Carbono e Metabolismo do Carbono e Efeitos do Stresse Hídrico na Metabolismo do Carbono e Efeitos do Stresse Hídrico na Efeitos do Stresse Hídrico na Efeitos do Stresse Hídrico na F
F F
Fotossínteseotossínteseotossíntese otossíntese
As plantas terrestres apresentam uma enorme diversidade de formas e estratégias adaptativas aos desafios impostos pelo meio ambiente. Também ao nível dos processos fotossintéticos demonstram esta variabilidade. As principais vias fotossintéticas denominam-se C3 e C4, devido ao facto de se ter observado que o primeiro produto estável da fixação de
CO2 era uma molécula com três e quatro átomos de carbono, respectivamente (Ehleringer e
Cerling, 2002). Existe uma terceira via fotossintética denominada CAM (de crassulacean acid metabolism) em que há uma fixação do carbono durante o período nocturno, altura em que os estomas estão abertos, ao contrário do que acontece com as outras duas vias fotossintéticas (Lüttge, 2004).
A fotossíntese C3 é um processo com várias etapas nas quais o CO2 atmosférico é
fixado em moléculas orgânicas estáveis, ocorrendo praticamente em todas as células do mesófilo das folhas. No primeiro passo, a ribulose-1,5-bisfosfato carboxilase/oxigenase (RuBisCO) cataliza a reacção de carboxilação de uma molécula de ribulose-1,5-bisfosfato (RuBP) com uma molécula de CO2 para formar duas moléculas de fosfoglicerato. Contudo, a
RuBisCO também apresenta afinidade para o O2, iniciando uma via metabólica – conhecida
por fotorrespiração – levando à perda de carbono e energia (Taiz e Zeiger, 2002; Ehleringer e Cerling, 2002).
A fotossíntese C4 representa uma modificação bioquímica da fotossíntese C3, de modo
a reduzir a actividade oxigenásica da RuBisCO, aumentando a concentração de CO2 junto
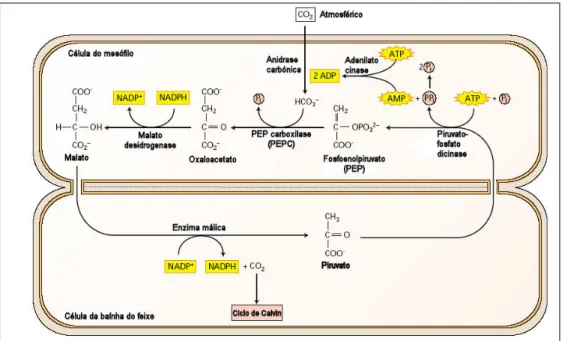
desta enzima. Nas plantas C4, a fotossíntese C3 encontra-se restrita às células da bainha dos
feixes vasculares (Fig. 2.1). É nas células do mesófilo nas quais se encontra a enzima fosfoenolpiruvato carboxilase (PEPC), a qual fixa o HCO3- (a partir do CO2 numa reacção
levada a cabo pela anidrase carbónica) e PEP em oxaloacetato, um ácido C4 que se difunde
para as células da bainha do feixe vascular, onde é descarboxilado (Ehleringer e Cerling, 2002).
A descarboxilação deste composto e a sua introdução na via C3 normal é realizado de
três formas, consoante o tipo de planta C4: através da enzima málica dependente de NADP
2. Introdução Geral NAD (EM-NAD) (como é o caso de Flaveria trinervia); e através da enzima fosfoenolpiruvato carboxicinase (PEPCK) (como é o caso de Panicum maximum) (Edwards et al., 2001; Walker et al., 2002). No entanto, esta divisão tem sido vista como uma simplificação do que realmente acontece, dado que o milho também possui uma actividade descarboxilativa considerável pela enzima PEPCK (Wingler et al., 1999) e na espécie Panicum maxicum a descarboxilação também é realizada pela enzima EM-NAD (Walker et al., 2002).
Independentemente da forma como é descarboxilado o composto C4, a actividade
elevada da PEPC conduz a uma elevada concentração de CO2 junto da RuBisCO, limitando a
actividade fotorrespiratória (Ehleringer e Cerling, 2002).
Fig. 2.1 – Esquema da via fotossintética nas plantas C4, do subtipo EM-NADP, como é o caso do milho (adaptado de Taiz e Zeiger, 2002).
As plantas C4, como o milho, têm sido documentadas como melhor adaptadas a
condições de défice hídrico devido a uma resistência estomática mais elevada, apresentando, uma melhor eficiência do uso de água e a capacidade de manter a actividade fotossintética em condições de baixo potencial hídrico (Bruce et al., 2002). O milho é capaz de tolerar um nível mais elevado de stresse e manter o crescimento das suas raízes e partes aéreas a níveis de stresse que são inibitórios para plantas C3, como o trigo (Triticum aestivum) (Siddique et al.,
1999; Nayyar, 2003).
Quando o stresse hídrico é imposto lentamente, como acontece normalmente em condições naturais, pode ocorrer uma redução na capacidade bioquímica de assimilação e
2. Introdução Geral utilização de carbono, juntamente com restrições na difusão gasosa (Chaves et al., 2002). Diferenças na resposta da fotossíntese à concentração interna de CO2 (Ci) indicam que a
necessidade bioquímica de CO2 é regulada em função da diminuição da disponibilidade de
CO2, associada ao fecho dos estomas induzido pela seca (Tallman, 2004). Existem dados
experimentais que demonstram que a diminuição das taxas de assimilação de CO2
encontradas em folhas sujeitas a stresse hídrico não pode ser revertida simplesmente aumentando o fornecimento externo de CO2, indicando que o stresse hídrico também deve
afectar o metabolismo do mesófilo (Lawlor, 2002; Parry et al., 2002; Tang et al., 2002; Cornic e Fresneau, 2002). Esta resposta do mesófilo torna-se progressivamente mais importante com o aumento da deficiência em água (Parry et al., 2002).
Numa experiência de campo com videiras, a assimilação de CO2 foi significativamente
limitada devido ao fecho dos estomas, à medida que a seca se instalava durante o período de Verão, mas também houve uma redução na actividade de diversas enzimas do ciclo de Calvin (Maroco et al., 2002).
Foi demonstrado que os efeitos do défice hídrico na capacidade fotossintética do tremoceiro dependiam da temperatura da folha e da luz incidente (Chaves et al., 1992 in Chaves et al., 2002). Foram obtidos dados semelhantes em diversas solanáceas, mostrando que a fotossíntese era significativamente menos inibida por temperaturas acima dos 38-40ºC em plantas desidratadas comparadas com plantas bem regadas (Havaux, 1992).
Numa experiência com duas espécies de Quercus não se encontraram diferenças significativas nas taxas de assimilação de carbono quando estas se comparavam entre plantas em situação de humidade suficiente no solo e plantas sujeitas a um stresse hídrico moderado no início de Julho. Contudo, no fim do Verão quente e seco (Setembro), as trocas gasosas, a meio do dia, na azinheira (Quercus ilex L.) eram menos reduzidas que as observadas no sobreiro (Quercus suber L.) (Faria et al., 1998 in Chaves et al., 2002). Isto sugere um efeito menos severo do stresse hídrico na primeira relativamente ao último. De facto, foram observados valores significativamente mais elevados de potencial hídrico nas folhas no fim do Verão de 1999 na azinheira (-1,52 MPa) quando comparado com os -2,38 MPa medidos no sobreiro. Uma das hipóteses é a de que as raízes da azinheira eram capazes de retirar água de camadas de solo mais profundas, permitindo a esta espécie manter o influxo de água e taxas de assimilação de carbono mais elevadas nas folhas, por um período de tempo mais longo, do que as do sobreiro (Chaves et al., 2002).
No final da época seca (Setembro), foi observada uma diminuição da fotossíntese na azinheira e no sobreiro, comparativamente com o início do Verão (Julho). Durante o período
2. Introdução Geral mais quente do dia, ocorreu uma diminuição da fotossíntese, associada a uma diminuição do rendimento quântico do fotossistema II. Neste período do dia, a assimilação de carbono estava limitada pela diminuição da condutância estomática, podendo aquela ser vista como um importante mecanismo protector contra a foto-oxidação e, portanto, contra a seca nestas árvores. Esta regulação da fotossíntese resulta de uma dissipação térmica do excesso de energia de excitação nos cloroplastos, como foi mostrado pelo aumento do amortecimento não-fotoquímico (NPQ) em Setembro quando comparado com Julho (Chaves et al., 2002).
Noutras espécies, sabe-se que a seca rigorosa diminui a quantidade da proteína RuBisCO (Vu et al., 1987). A quantidade de RuBisCO nas folhas é controlada pela sua taxa de síntese e degradação. Mesmo sob stresse hídrico, a holoenzima da RuBisCO é relativamente estável, com um tempo de semi-vida de vários dias (Webber et al., 1994 in Parry et al., 2002). Contudo, o stresse hídrico no tomate (Bartholomew et al., 1991), em Arabidopsis (Williams et al., 1994 in Parry et al., 2002) e no arroz (Vu et al., 1999 in Parry et al., 2002) leva a uma rápida diminuição na abundância de transcritos da subunidade menor da RuBisCO (rbcS), o que pode indicar uma diminuição na sua síntese (Marques da Silva e Arrabaça, 2004b).
Em alguns estudos, o potencial fotossintético (Apot), medido em condições saturantes
de CO2, não é afectado por uma pequena diminuição do teor hídrico relativo (THR) mas
torna-se progressivamente mais inibido, e menos estimulado por elevadas concentrações de CO2, abaixo de um determinado valor de THR (resposta Tipo I). Noutros estudos, Apot e a
estimulação da taxa fotossintética por elevadas concentrações de CO2 diminui
progressivamente à medida que THR diminui (resposta Tipo II) (Lawlor, 2002).
Diversos estudos sugeriram que a diminuição da capacidade fotossintética resulta da insuficiente regeneração de ribulose-1,5-bisfosfato (RuBP) (Giménez et al., 1992). Se isto é provocado ou não pela diminuição da síntese de ATP na fase fotoquímica continua a ser discutido (Gunasekera e Berkowitz, 1993; Tezara et al., 1999; Lawlor, 2002).
Embora a actividade da RuBisCO possa não ser a principal limitação do metabolismo cloroplastidial, os efeitos do stresse hídrico na quantidade e actividade da RuBisCO não podem ser ignorados. No estudo de Tezara et al. (1999), a disponibilidade de ATP diminuiu, tendo sido identificada como um factor chave na tolerância ao stresse hídrico. No entanto, as alterações nos níveis de ATP foram inferiores às verificadas para a actividade da RuBisCO, em potenciais hídricos baixos, quando a assimilação de CO2 estava muito reduzida.
A actividade da RuBisCO é regulada para ajustar à capacidade da folha em regenerar RuBP, sendo modulada in vivo quer pela reacção com CO2 e Mg2+ para carbamilar o resíduo
2. Introdução Geral de lisina no local catalítico, quer pela ligação de inibidores no local catalítico (Parry et al., 2002). A ligação dos inibidores à RuBisCO carbamilada previne a catálise, enquanto que a ligação do substrato RuBP na enzima não-carbamilada previne a carbamilação do resíduo de lisina que é essencial para a actividade. A libertação de tais compostos tão fortemente ligados necessita da participação da RuBisCO activase e da hidrólise de ATP (Zhang e Portis, 1999). Enquanto que a medição da actividade da RuBisCO imediatamente após a extracção (actividade inicial) reflecte a actividade in vivo, a carbamilação potencial (actividade total) pode ser determinada incubando os extractos com elevadas concentrações de CO2 e Mg2+
antes do ensaio. Contudo, a carboxilação máxima potencial (actividade máxima) apenas é revelada se forem dados passos para, em primeiro lugar, remover quaisquer inibidores que estejam ligados aos locais activos (Parry et al., 2002). A maioria dos inibidores pode ser removido in vitro através de altas concentrações de sulfato (Parry et al., 1997).
No tabaco (Nicotiana tabacum L.), a seca diminuiu a actividade inicial e total da RuBisCO. A diminuição da actividade aparenta ser o resultado de uma diminuição do Kcat
aparente e não de uma alteração do estado de activação. Também têm sido observadas diminuições semelhantes na Kcat da RuBisCO em resposta ao stresse hídrico de longa duração
no trevo (Medrano et al., 1997 in Parry et al., 2002).
Foi demonstrado que a ligação do inibidor CA1P à RuBisCO protege a enzima da degradação proteolítica (Khan et al., 1999). A RuBisCO que se encontra activa na carboxilação da RuBP é relativamente resistente à degradação por proteases; por outro lado, a RuBisCO não-carbamilada com locais activos disponíveis é muito susceptível à proteólise. As interacções com inibidores podem apresentar vantagens in vivo, de modo a prevenir a degradação por proteases da RuBisCO não activa.
Como já foi mencionado, a libertação dos inibidores de ligação forte necessita da participação da RuBisCO activase e da hidrólise de ATP (Zhang e Portis, 1999). A remoção dos inibidores pela RuBisCO activase pode ser impedida porque, como já foi sugerido por alguns trabalhos, as concentrações de ATP são reduzidas por acção da seca (Lawlor, 2002). Além disso, a RuBisCO activase é susceptível a temperaturas elevadas (Crafts-Brandner e Salvucci, 2000) que podem estar associadas ao stresse hídrico.
No trigo, a diminuição do THR não conduziu ao aumento dos inibidores de ligação forte. Este resultado contradiz estimativas indirectas da percentagem de locais catalíticos bloqueados por inibidores (calculados a partir das actividades máxima e total), o qual sugeria que um número elevado de locais deveriam estar bloqueados por inibidores. Esta discrepância
2. Introdução Geral pode ser explicada por outros factores, dado que o trigo não contém grandes quantidades de CA1P (Servaites et al., 1986).
Existem dois mecanismos a ser considerados em relação à diminuição da actividade da RuBisCO. Primeiro, a RuBisCO reverte a um estado de baixa actividade in vitro através de uma mudança conformacional (Schmidt et al., 1984). No entanto, a hipótese deste mecanismo ter um papel regulador in vivo tem recebido pouca atenção. A segunda hipótese é a inibição da RuBisCO por ligação da RuBP aos locais não-carbamilados (Parry et al., 2002).
Em resumo, em condições de campo, quando o stresse hídrico moderado se desenvolve lentamente, a diminuição do CO2 intracelular após o fecho estomático
aparentemente induz, a longo prazo, a regulação da maquinaria fotossintética, nomeadamente da RuBisCO, de modo a ajustar ao fluxo de carbono disponível (Chaves et al., 2002).
2.5 2.5 2.5
2.5.... Estratégias de Sobrevivência à S Estratégias de Sobrevivência à S Estratégias de Sobrevivência à Seca Estratégias de Sobrevivência à Secaeca eca
2.5 2.5 2.5
2.5....1. 1. 1. 1. Aspectos GeraisAspectos GeraisAspectos GeraisAspectos Gerais
A adaptação das plantas à seca tem sido classificada em três estratégias: escapar, evitar e tolerar (Chaves et al., 2003). No entanto, estas estratégias não são mutuamente exclusivas e, na prática, as plantas combinam uma variedade de tipos de resposta (Ludlow, 1989 in Chaves et al., 2003).
As plantas que escapam à seca exibem um elevado grau de plasticidade no seu desenvolvimento, sendo capazes de completar o seu ciclo de vida antes da ocorrência do défice hídrico. Estas estratégias têm como base o sucesso reprodutivo antes do stresse se agravar, sendo de particular importância nas plantas de locais áridos, em que há combinação de ciclos de vida curtos com elevadas taxas de crescimento e de trocas gasosas, maximizando o aproveitamento dos recursos hídricos, florescendo e produzindo sementes antes que as reservas de água se esgotem (Chaves et al., 2002, 2003). Outro mecanismo para um sucesso reprodutivo consiste em mobilizar parte dos recursos para o desenvolvimento de frutos, mecanismo associado à capacidade da planta em armazenar reservas em alguns órgãos (caules e raízes) e mobilizá-los para a produção de frutos, resposta bem documentada em algumas plantas cultivadas, como os cereais (Gebbing e Schnyder, 1999; Bruce et al., 2002) e alguns legumes (Chaves et al., 2002). Esta capacidade aumenta em plantas sujeitas à seca (Rodrigues et al., 1995).
2. Introdução Geral As plantas também podem superar as condições de seca evitando a desidratação dos tecidos, enquanto mantém o potencial hídrico nos tecidos o mais elevado possível, ou tolerando um potencial hídrico baixo no tecido. A desidratação é evitada em plantas anuais ou perenes e está associada com uma variedade de características adaptativas. Estas envolvem diminuição das perdas de água e maximização da tomada de água. As perdas de água podem ser minimizadas pelo fecho dos estomas, por redução da superfície de contacto com a atmosfera através do enrolamento das folhas (Price et al., 2002), através de uma camada densa de tricomas aumentando a reflexão (Werker, 2000), através de ângulos fechados das folhas, ou diminuindo a área da copa através da redução do crescimento e da queda das folhas mais velhas. A tomada de água é maximizada ajustando o padrão de alocação de carbono, nomeadamente aumentando o investimento na formação de raízes (Jackson, 2002). Foram obtidos aumentos significativos na produção de espécies agrícolas, cruzando-as com plantas de regiões semi-áridas, resultando num melhoramento da profundidade de enraizamento (Blum, 1984 in Chaves et al., 2003).
A dormência parcial das plantas como sobrevivência à estação seca é outra estratégia encontrada. A tolerância extrema à desidratação encontra-se nas denominadas plantas da ressurreição, bem como em alguns fetos, plantas não-vasculares (musgos), algas e líquenes. As folhas das plantas da ressurreição podem equilibrar com o ar perto de 0-2% (v/v) de humidade relativa e ainda conseguirem recuperar totalmente a sua actividade fisiológica após re-hidratação (Chaves et al., 2003).
2.5 2.5 2.5
2.5....2. Modificações Anatómicas, Morfológicas e Metabólicas na 2. Modificações Anatómicas, Morfológicas e Metabólicas na 2. Modificações Anatómicas, Morfológicas e Metabólicas na 2. Modificações Anatómicas, Morfológicas e Metabólicas na R
R R
Resposesposesposesposta ao Stresse Hta ao Stresse Hta ao Stresse Hta ao Stresse Hídricoídricoídricoídrico
Para além da diminuição dos recursos hídricos, a seca do solo também induz uma diminuição de nutrientes, em particular o azoto (mas também o cálcio), com fortes efeitos interactivos no crescimento e funcionamento da planta (Chaves et al., 2003). Esta é uma área de investigação de grande importância, necessitando de métodos inovadores que permitem distinguir os efeitos causados pelo défice hídrico dos efeitos causados pela deficiente nutrição mineral, ambos causados pela seca.
Os ajustamentos da planta aos baixos recursos dos ambientes áridos incluem alteração da estrutura foliar, aumento da proporção de assimilados mobilizados para as raízes (Poorter e Nagel, 2000), reduzidas taxas de crescimento e baixas taxas de turnover dos órgãos (Enquist e Niklas, 2002).
2. Introdução Geral Enquanto que órgãos com curtos períodos de vida podem ser eliminados em resposta ao stresse, órgãos com longos períodos de vida têm que optimizar o seu ganho em recursos. Este é o caso da mudança de demografia foliar observado na espécie desértica perenial Cryptantha flava, cujo período de vida foliar duplicou em condições de seca (Casper et al., 2001). Esta resposta resulta em taxas de turnover foliar reduzidas e assim elevada eficiência do uso de nutrientes. Isto também significa que as plantas podem compensar parcialmente taxas de ganho de carbono mais reduzidas investindo menos na construção de novas folhas.
As folhas com maior longevidade necessitam sobreviver a períodos em que as condições são mais hostis. Para tal, necessitam de diversas protecções que variam desde características anatómicas/morfológicas, como a esclerofilia para resistir a climas extremos e à herbivoria (Lucas et al., 2000; Burghardt e Riederer, 2003), uma densa camada de tricomas como na oliveira (Olea europaea L.) para aumentar a reflectância da luz e manter a humidade (Werker, 2000), ou ângulos foliares fechados (Lizana et al., 2006), até mecanismos bioquímicos que permitem a dissipação do excesso de energia radiante (Young et al., 1997).
As folhas esclerófilas resistem a condições climáticas extremas e à herbivoria (Lucas et al., 2000; Burghardt e Riederer, 2003), e são normalmente pequenas e grossas, possuindo uma reduzida área foliar específica (SLA) e baixa concentração de azoto por unidade de massa foliar. Uma diminuição na SLA também pode ocorrer nas plantas herbáceas como resposta à seca, resultando num aumento no investimento de tecidos estruturais, o que permite uma maior resistência a condições ambientais desfavoráveis (Chaves et al., 2003). Outras protecções incluem mecanismos bioquímicos de dissipação de energia, como no caso do ciclo das xantofilas (Young et al., 1997).
Folhas pequenas estão bem adaptadas à elevada luminosidade e à elevada temperatura que existem na maioria das regiões áridas, porque o seu tamanho permite uma maior dissipação do calor e um controlo eficaz das perdas de água através do fecho dos estomas (Jarvis e McNaughton, 1986 in Chaves et al., 2003).
A senescência, que leva a uma diminuição da reserva de açúcares na planta, é um factor importante na interrupção da formação do fruto, em condições de stresse hídrico. Assim, os fenótipos com senescência retardada podem ser importantes em culturas onde o rendimento seja limitado pelos recursos e a acumulação e utilização das reservas dos caules seja insuficiente para suportar o crescimento do fruto em condições de seca. Contudo, nos cereais, observa-se uma interacção interessante entre a disponibilidade hídrica e a disponibilidade de azoto. Em condições de disponibilidade de água não-limitante, azoto abundante atrasou a senescência mas reduziu o rendimento da produção de grão, enquanto