GABA-induced inactivation of
Cebus
apella
V2 neurons: effects on orientation
tuning and direction selectivity
A.K. Jansen-Amorim
1, M. Fiorani
2and R. Gattass
21Instituto de Cieˆncias Biolo´gicas, Universidade Federal do Para´, Bele´m, PA, Brasil 2Instituto de Biofı´sica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil
Abstract
We investigated the GABA-induced inactivation of V2 neurons and terminals on the receptive field properties of this area in an anesthetized and paralyzedCebus apellamonkey. Extracellular single-unit activity was recorded using tungsten microelectrodes in a monkey before and after pressure-injection of a 0.25 or 0.5 M GABA solution. The visual stimulus consisted of a bar moving in 8 possible directions. In total, 24 V2 neurons were studied before and after blocker injections in 4 experimental sessions following GABA injection into area V2. A group of 10 neurons were studied over a short period. An additional 6 neurons were investigated over a long period after the GABA injection. A third group of 8 neurons were studied over a very long period. Overall, these 24 neurons displayed an early (1-20 min) significant general decrease in excitability with concomitant changes in orientation or direction selectivity. GABA inactivation in area V2 produced robust inhibition in 80% and a significant change in directional selectivity in 60% of the neurons examined. These GABA projections are capable of modulating not only levels of spontaneous and driven activity of V2 neurons but also receptive field properties such as direction selectivity.
Key words: Visual cortex; Single units; Extracellular recordings; New World monkeys
Introduction
Since the early works of Hubel and Wiesel (1) on the response properties of V1 neurons, a substantial amount of data on the functional organization of the visual cortex has been accumulated. In each visual cortical area, neurons have a unique inventory of receptive field properties. Each of these properties is combined with those of neighboring neurons to form orderly representa-tions of stimulus features; these different representarepresenta-tions are superimposed within the same population of neurons (1). Since the discovery that cells in the visual cortex are selective for the orientation of contours, the mechanisms underlying cortical orientation selectivity have been the subject of intense debate. These features are widely believed to emerge from particular combinations of ascending, descending and lateral projections (2,3). The major excitatory input to cells in all layers is derived from a massive convergent input from other cortical cells. This cortical input has the potential to enhance orientation and direction selectivity via excitatory interactions both within the same orientation column (4) and between columns with like orientation preferences (5).
Neurons in early visual areas, such as V1 and V2, are selective for the orientation and direction of movement of visual stimuli. It has been suggested that some receptive field properties of cortical neurons, such as orientation and direction selectivity, may be attributed to the inhibitory influence of intrinsic circuits on incoming information (6,7). The inactivation of intrinsic inhibitory processes impairs both orientation and direction selectivity (8,9). The record-ing of neurons targeted and inactivated by gama-amino butyric acid (GABA) at sites in cat area 18 revealed that inactivation at sites with similar orientation preference produced a significantly greater effect on directionality relative to the minor effect on directionality observed following inactivation of regions of cross-orientation. Effects on orientation tuning were produced almost exclusively by cross-orientation sites, while changes in directionality were produced mainly by sites of the same orientation (10,11). Studies that used labeling with tritiated nipecotic acid to identify GABAergic neurons at the site of inactivation showed that orientation-tuning V2 neurons change their specificity due to the inhibition of
Correspondence: R. Gattass, Instituto de Biofı´sica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, 21941-900 Rio de Janeiro, RJ, Brasil. E-mail: [email protected]
cross-orientation-selective neurons via GABAergic neurons (7). In conclusion, in the primary (V1) and secondary (V2) visual areas of monkeys and cats, the orientation and direction selectivity of neurons depend on the inhibitory influence of basket cells projecting to orientation- and direction-selective functional modules (6,7,11-13).
In the present study, we investigated whether extrinsic GABA-mediated projections (feed-forward or feedback) and/or intrinsic projections from area V2 directly interfered with the orientation and direction selectivity of V2 neurons. V2 has a surface area of about 625 mm2. The injections of GABA encompassed 4-10 mm2of the cortical area (or a
volume of 8-20 mm3), representing up to 1.6% of V2 (14). The injections of lidocaine encompassed about 36 mm2 (96 mm3), representing up to 6.8% of V2, and almost its entire width of 6-7 mm. These estimates are consistent with the research of Hupe´ et al. (15). They performed a quantitative study on the spread of GABA after pressure injections into the cortex and predicted that the injected volume would spread throughout the extracellular space, occupying an ellipsoid shape. Considering the extent of area V2 determined by Rosa et al. (14), we extended the predictions of Hupe´ et al. (15) regarding the relationship of injected volume and occupied extracellular volume. We predicted that GABA injection volumes between 120 and 600 nL would inactivate 0.02-0.16% of area V2, while 1.8 and 6mL lidocaine would inactivate 0.2-1.6% of V2.
We studied the receptive field properties of V2 neurons before and after the inactivation of a small portion of area V2 in the capuchin monkey (Cebus apella). Several aspects of the visual system of this New World monkey, including the thalamic organization (16-18), intrinsic circuitry of V1 (19-21), and topographical characteristics of areas V1, V2, MT, and V4 (14,22-25) have been studied for almost two decades, making this monkey a suitable experimental model for this study. In addition, we recently studied the effect of GABA inactivation of visual area MT (26), visual area V4 (27), and previously (2004) the pulvinar (28) on the direction and orientation selectivity of V2 neurons. These studies enabled us to make direct comparisons regarding GABA inactivation on the feedback connections from MT and V4.
Material and Methods
Animals
One adult male Cebus apellamonkey was used in 5 recording sessions, performed once a week. All the experimental protocols were conducted following the National Institutes of Health (NIH) guidelines for animal research and were approved by the Committee for Animal Care and Use of the Instituto de Biofı´sica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro (2011).
Location of V2 and preparations for the recording sessions
V2 is a belt of cortex located in the opercular region of
the occipital pole in front of V1. It is limited posteriorly by V1, except in the most anterior region of the calcarine sulcus, which borders the area of prostriata of Sanides (14,21). V2 is limited anteriorly by V3 except at the representation of the fovea region at the lateral surface between the lunate sulcus and the inferior occipital sulcus, where V2 borders V4 (21). InC. apella, V2 is wider (8-11 mm) in the region of intermediate representation of the visual field (about 106eccentricity) and narrows (4-5 mm) in the bottom of the sulci representing the foveal region (14).
Prior to the recording sessions, and under anesthesia and aseptic conditions, a head bolt and a recording chamber were attached to the skull. The position of the recording chamber was determined by visualizing the vascularization of the sulci through the skull. The superior temporal sulcus (STS) is always visible and it constitutes an important anatomical landmark. The intraparietal sulcus is also quite visible, but it is less conspicuous than the STS. The lunate sulcus is far less clear than the STS and the intraparietal sulcus. The location of the posterior border of V2 is 2-3 mm posterior to the lunate sulcus. Using stereotaxic coordinates and the position of the cortical sulci, we positioned the recording chamber such that we had access to area V2. During each recording session, the animal was anesthetized with a 5% ketamine hydrochloride solution (30 mg/kg, intramuscular; Ketalar, Parke Davis, Brazil) and maintained with a mixture of 70% nitrous oxide and 30% oxygen combined with a continuous intravenous infusion of fentanyl citrate (0.003 mg?kg-1?h-1). After endotracheal intubation with a newborn#3.5 cannula, the monkey was also immobilized with pancuronium bromide (0.1 mg?kg-1?h-1) and maintained under mechanical ventila-tion (Primate ventilaventila-tion pump, Harvard Apparatus, USA) adjusted to obtain 3-4% expired CO2. Electrocardiogram
readings, body temperature, and end-tidal CO2 were
monitored continuously. Post-surgical analgesia was maintained for 3 days with the administration of fentanyl skin patches (Durogesic1, Janssen-Cilag, Brazil).
After implant of the recording chamber, the bone inside the chamber was removed, leaving the dura intact. After 10 days, a thick fibrous growth covered the dura and prevented the drying of the cortex.
Recording sessions
To locate V2, we penetrated the cortex with 1 MV-impedance tungsten microelectrodes (Frederik Haer 250 m-w, USA), using stereotaxic coordinates and sulcal landmarks (14,23). The V2 stereotaxic coordinates were used to allow access to the central lower field representa-tion of area V2 in subsequent recording sessions. The injection and recording sites were typically located 6-10 mm from the midline, 1 mm behind the lunate sulcus, at the depth of 4-6 mm from the cortical surface.
Inactivation devices
two stainless steel tubes (external diameter: 400mm; internal diameter: 200mm) mounted around a tungsten microelectrode. The entire system measured approxi-mately 800mm. A pneumatic pico pump (Model PV 820, World Precision Instrument, USA) delivered the GABA (Sigma, USA) solution to the two external stainless steel tubes through a polyethylene connection. GABA was diluted in deionized water and stored at 46C. Lidocaine was used straight from the bottle. With this system, it was possible to deliver 120 to 600 nL of a 0.25 M GABA solution or 1.8 to 6mL 2% lidocaine (Xylocaine1, Astra Zeneca, USA) to the region of interest. The injections were administered over a period of 30 s.
After the topographical site was localized in the V2, a single tungsten microelectrode was replaced by the injection system, with stainless steel tubes placed approxi-mately 800mm apart, to inactivate V2 neurons. Single-unit activity from area V2 was recorded using tungsten microelectrodes. The activity was amplified (10,000X) and filtered (band pass, 60-6000 Hz), and single spikes were sampled at 166 kHz by a waveform discriminator system (SPS-8701, Signal Processing System, Australia). The computer-based discriminator SPS-8701 uses a template-matching algorithm to sort up to three spikes. Extracellular single-unit spike events were stored using CORTEX software (Laboratory of Neuropsychology, NIMH/NIH, USA) for offline analysis. The receptive fields were initially localized and mapped using a hand-plot mapping procedure.
The receptive field automatic mapping procedure was based on computing the latency-corrected neuronal activity in response to elongated bars moving in 1 of 8 directions of motion. Initially, peristimulus time histograms (PSTHs) were computed based on 10 stimulus presenta-tions, using a bin width of 10 ms. Single-trial spike trains were used to produce PSTHs that were aligned to stimulus onset. The PSTHs were then smoothed using a normal convolution filter with a 200-ms time window, which resulted in the time spike density function. This continuous and derivable function well characterizes the neuronal firing pattern (see Ref. 26).
Inactivation paradigm
Area V2 was inactivated until a significant reduction of the recorded activity at the injection site was obtained. Data were collected immediately before and after the injection, and several blocks of recording protocols were acquired until recovery of V2 cellular activity. The recording sessions typically continued for 14 h. The interval between experimental sessions was at least 10 days. With this paradigm we could not discriminate whether extrinsic GABA-mediated projections (feed-forward or feedback) or intrinsic projections from area V2 were interfering with the orientation or direction selectivity of V2 neurons. We therefore added very large injections of a local anesthetic to the paradigm to try to
segregate the effect on the responsiveness of the neuron from that on the circuit underlying directional or orientation selectivity. For these reasons we used very large injections of 2% lidocaine (1.8-6mL) to compare to restricted (120-600 nL) injections of 0.25 M GABA.
Recordings from area V2 were obtained before (control) and during several sequential time blocks at different time points (1-300 min) after GABA inactivation of area V2. The neuronal responses at the injection site in area V2 were analyzed by comparing an effective direction-selective polar plot (polargram) with a post-stimulus raster in a histogram plotted as the spike density functions of the neuronal discharge. These effects were compared to the inactivation with the local anesthetic lidocaine.
The electrodes were used once and discarded. The inactivation system was sterilized before use with neutral detergent, bleach, and Germi-Rio (Rioquı´mica, Brazil).
Visual stimulation
Prior to the recording sessions, gas-permeable con-tact lenses were used to focus the monkey’s eyes on a computer monitor placed 57 cm in front of the animal. The positions of the blind spot and fovea were plotted onto the computer screen with a reversible ophthalmoscope. The stimulus consisted of a thin white bar (1860.56) that
appeared in four random orientations (06, 456, 906, and 1356), crossed the screen in a direction perpendicular to its orientation at a velocity of 106/s, and passed through the receptive fields of all recorded neurons. We con-tinuously tested the direction of motion selectivity before and after GABA injection. Note that we did not segregate orientation selectivity from axis-of-movement selectivity. Therefore, the moving bar in this configuration was used to determine both the direction and axis of movement selectivity.
Data analysis
grouping the conditions with the same stimulus orientation moving in opposite directions, we tested this bidirectional component. The four groups of data generated by this procedure were then submitted to 1WA to evaluate changes in the selectivity of the cell for axis of movement. Probability values mentioned in the text, which were related to the changes in direction selectivity, were evaluated by ANOVA unless otherwise specified.
The selectivity of the neurons was examined with a standard test of circular tuning to determine the magni-tude of the GABA-induced changes in both direction and orientation selectivity across the population. The orienta-tion selectivity index (OI) was calculated for each neuron according to the following formula, as proposed by Sato et al. (6): OI=[(SRi sin(20i))2+(S[Ri cos(20i))2]0.5/SRi,
where Rirepresents the magnitude of response to each
stimulus orientation,0i. The rate of spontaneous activity,
which was measured for the 700 ms window before stimulus presentation, was subtracted from the stimulus-driven responses. The response to each stimulus orientation is expressed as a vector with direction 20i
and length Ri. This index varied between 1 and 0, with an
OI value of 1 indicating complete selectivity to a particular orientation and a value of 0 indicating equal responsive-ness to all orientations.
Direction selectivity indices (DI) were also quantified by the directional index, as described by Wang et al. (29). An index of 1 indicates sharply tuned cells, whereas an index of 0 indicates non-tuned cells. We used a paired Studentt-test to evaluate changes in the mean OI and DI in the cell population. Using this test, we also evaluated the recovery time after GABA injection by comparing cell activity in the control condition both before the GABA injection and after the GABA-induced effects had vanished. In addition, we used the individual index values to reveal any bias in the population. We considered a change greater than 0.20 in the OI or DI to be significant. Population bias was evaluated using thex2test.
Histological procedures
After a complete set of inactivation sessions, the animal was killed for histological processing to localize the injection and recording sites and to examine the structure of the cortical layers. At the end of the last recording session, the animal was deeply anesthetized with sodium pentobarbitone (30 mg/kg) and perfused intracardially with saline, followed by 2% paraformaldehyde in phosphate-buffered saline (PBS), 2% paraformaldehyde in PBS supplemented with 2.5% glycerol, PBS supplemented with 5% glycerol, and PBS supplemented with 10% glycerol. Frozen sections (70mm thick) were cut on a cryostat and mounted on glass slides or kept in PBS at 46C. The sections were stained using the Nissl (cresyl violet) stain and were analyzed on a slide projector and microscope to determine the locations of the electrode tracks and the absence of cortical lesions.
Results
GABA inactivation of visual area V2 produced a general decrease in the excitability of the neurons, which included a decrease in spontaneous and driven activities followed by changes in direction and/or orientation selectivity.
The results of the V2 inactivation in the same area of the recordings were combined into three groups. The first group, composed of 6 neurons, was subjected to 1 injection of 120 nL 0.5 M GABA. The second group, composed of 8 neurons, was subjected to 2 injections (1.8 and 6mL) of 2% lidocaine. The third group, composed of 8 neurons, was subjected to 1 injection of 600 nL 0.25 M GABA. We recorded a total of 24 cells in V2 during inactivation of this region.
The inactivation of all neurons produced significant effects in the majority (80%) of cases. The statistical analysis by 2WA showed that in slightly more than half of the cases (8/14), the cells were either directionally selective (1WA) or bidirectionally selective (2WA) and lost these properties after the GABA injections. The GABA inactivation also caused three previously non-direction selective cells to become selective to movement of the bar. Of those three, two became directionally selective (1WA) and one became bidirectionally selective (2WA).
Effect on V2 neurons
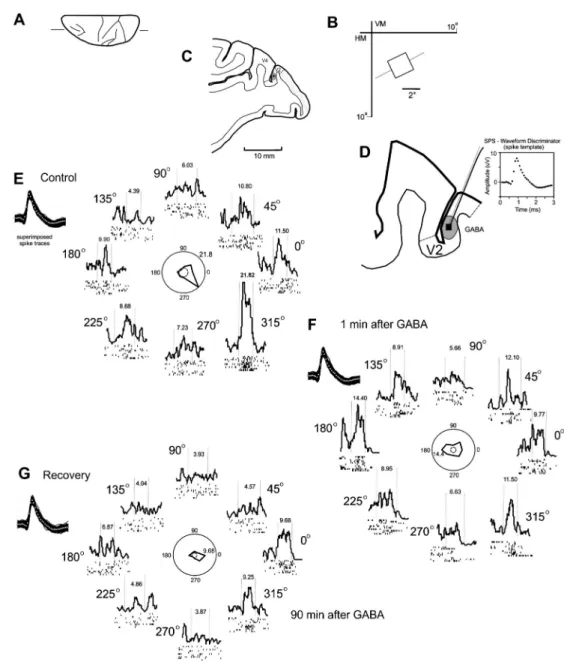
The injection of 120 nL 0.5 M GABA in V2 caused a local decrease in the spontaneous and driven activity of V2 neurons. Before the injection, there was a high level of spontaneous activity and a strong response to the moving bar. Figure 1 illustrates a neuron from area V2 that lost its unidirectional selectivity (2WA, P,0.05,F=56, d.f.=862)
as a result of GABA-induced inactivation. Under the control conditions, this neuron was highly selective to movement in the direction of 3156. The response decreased with the injection of GABA, while the response to the direction of 1806increased (1WA, P.0.05). After 90 min, the neuron again displayed selectivity toward a value quantitatively similar to initial control conditions (2WA, P,0.05), despite an overall lower excitability of the neuron.
Figure 2 illustrates the effect of an injection of 120 nL 0.5 M GABA on the firing rate of 6 V2 neurons. The spontaneous activity and best direction response, in most neurons, decreased after the GABA injection, similar to what is observed following the inactivation of MT and V4. This effect is not consistent for all directions, as some neurons decreased while others increased the firing rate after GABA injection.
Time course of the GABA effect
displayed a decreased response to a bar in their preferred direction. A similar effect was observed following an injection of 120 nL 0.5 M GABA. Indeed, this decrease in responsiveness was also apparent following inactivation of MT and V4. The initial decrease of spontaneous neuronal activity was not as marked as the observed decrease following the injection of 120 nL 0.25 M GABA.
A direct effect of the GABA injection in area V2 on the suppression and subsequent recovery of response to the best orientation or direction in the V2 cells was established. The effect of an injection of GABA in other directions on the response to the stimulus is variable, as some neurons decreased while others increased their rate of firing following the injection.
Lidocaine effect
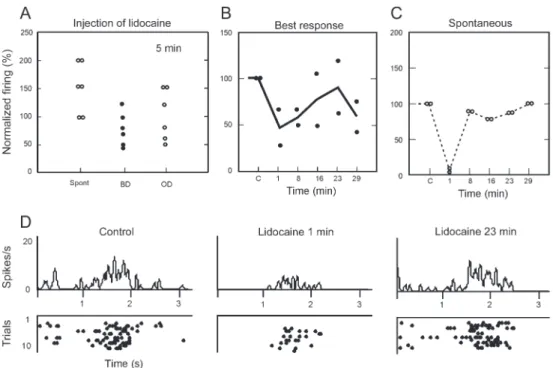
In the second group, neurons received 2% lidocaine. Figure 4 illustrates the response of V2 neurons that lost direction selectivity as a result of lidocaine-induced inactivation (1WA, P,0.05, F=41, d.f.=8). This cell exhibited directional selectivity during the control condition
that for both the best direction and the spontaneous activity, the trend is toward the return of the neuronal cell firing to control levels. The main effect of lidocaine injection was a reduced response to the best direction or orientation. In other directions or orientations, the result varied.
In most cases, lidocaine increased the amount of spontaneous activity (Figure 5A). Figure 5B and C illustrate the effect of lidocaine in two neurons over approximately 30 min of recording. Figure 5D illustrates
signal/noise ratio. The response recovery over time was equivalent to that of the control 23 min after injection. Thus, the effect of lidocaine appears to have three phases: the general inhibition of the response and spontaneous activity, the increase in the signal/noise ratio, and the return to control response level.
Figure 6 shows changes in the direction and orienta-tion tuning of 24 V2 cells after GABA- and lidocaine-induced inactivation of the area V2. When changes greater than 0.2 were used as a criterion, 14 neurons (29.2%) changed their direction or orientation tuning.
There was no trend in the sample of cells. Direction tuning of V2 neurons either increased (4/24) or decreased (5/24) after injections of GABA or lidocaine. In addition, these cells decreased (2/24) and increased (3/24) their orientation selectivity. The effect of GABA inactivation of V2, unlike the effect produced by inactivation of MT and V4 areas, decreases the indices of orientation or direction selectivity.
Discussion
A number of previous studies have described the intrinsic connections in area V2 in the monkey and cat (6,7,10-13,30). Hupe´ et al. (15) performed a quantitative study on the spread of GABA after pressure injection into the cortex. An injected volume of 13 nL 0.1 M GABA would occupy an extracellular volume of approximately 60 nL. For injected volumes greater than 60 nL, it would theoretically be difficult to predict the shape of the spreading. Hupe´ et al. (15) also determined that, when larger volumes of GABA were injected, the inactivation period and the time needed for the neurons to recover would also be greater. Our observations are in agreement Figure 5.Effects and time course of an injection of 2% lidocaine into area V2 on the response of 6 neurons.A, Effect on the normalized firing rate for the spontaneous activity (Spont), best direction (BD) and other direction (OD) after injection.B, Time course of the effect of lidocaine injection on the best response andC, on the spontaneous activity of 6 neurons.D, Post-stimulus time histograms and spike raster of a neuron at three different times: before (control), and after (1 and 23 min) the injection.
with Hupe´ et al. (15) and their later results (31). In Hupe´ et al. (31), the neurons required 40 min to recover to baseline after a 0.9mL injection of 0.1 M GABA. The duration of the inhibition described in the present study is consistent with that reported by Hupe´ et al. (31).
Inactivation techniques
Lidocaine, a fast action blocker, has an immediate effect that lasts less than 30 min (32,33). This anesthetic works by blocking the sodium channels of the body and the axon of the neuron, thus preventing the initiation and conduct of the neural signal. The main disadvantage appears to be the inactivation of passage fibers.
The evidence demonstrating that GABA may be an inhibitory neurotransmitter in the visual cortex is attributed to Sillito (34). More recently microiontophoresis and pressure injections have been used to study the effects produced by inactivation of GABA on the line orientation and direction in areas 17 and 18 of cat and monkey (6,7,10-13,30). GABAAreceptors are located mainly in the
cell body and act through activation of chloride channels. These receptors are selectively blocked by bicuculline, while the chloride channel itself is blocked by picrotoxin. GABA can be applied numerous times, produce inactiva-tion of short and long durainactiva-tion (26-28), and does not affect fibers of passage. Its effect depends on the mode of application (pressure or iontophoresis) producing inacti-vation within a distance of 2-4 mm (31).
Injections of lidocaine should produce an effect on the neurons of area V2 circuitry that is more intense and more widespread given that all neurons have the sodium channel. Injections of GABA should affect a specific neural population: those with GABAAreceptors and thus
produce a less intense and more focal effect. This theoretical prediction is consistent with the results obtained in this study. However, the optimal experimental design to answer that question definitively would be the sequential injection of two blockers in the same experi-ment, which would otherwise have other impeding issues.
Evaluation of the early and late effects of GABA inactivation
GABA inactivation of V2 intrinsic circuitry produced early (up to 20 min) and late (20-150 min) after-injection effects on intrinsic V2 neurons. The early effects consisted of a general decrease in neuronal excitability, which corresponded to a depression in the spontaneous and driven activities. The later effects generally reflected changes in the orientation and/or direction selectivity of the V2 neurons.
The V2-driven activity typically fell below 50% of the control level during the first 10 min after injection. The different effects of GABA inactivation on the spontaneous and stimulus-driven activity were complex, and we attributed them to the properties of the GABAergic channels. This dissociation suggests that different and
possibly independent mechanisms underlie the changes in the stimulus-driven and spontaneous activity in the cortex.
Duration of inactivation
Prior to the start of these experiments, we were unable to predict the long-lasting effects of GABA injection into area V2. We believe that the inhibitory effects detected in the present study are equivalent to the effects of iontophoretic injections of GABA described in the litera-ture (7,11,12). With iontophoresis, however, the amount of GABA released is usually much smaller than that released from a pressure injection. It is therefore possible that, in addition to an initial inactivation effect, a pressure injection causes late effects in neuronal activities that are not observed when lower quantities of GABA are injected. Several changes occurred in the V2 neurons, suggesting that V2 neurons could be influenced by GABA inactiva-tion. These effects persisted for 40-299 min.
The effects of pharmacological inactivation are usually reversible and are evaluated in comparison to the pre-injection state. A return to the resting or pre-pre-injection spontaneous and driven activity is indicative of a non-toxic or non-destructive effect on the central nervous system tissue. Therefore, we based our results on a sample of cells in which a clear trend towards the recovery of the driven activity toward baseline was observed.
V2 cortical circuits
Several studies have demonstrated the role of intrinsic circuits in the regulation of receptive field properties (6,7,10-13,30). The intrinsic connectivity of V2 was studied by Rockland (35) in S. sciureus and Macaca spp. The connections of area V2 originate from pyramidal neurons in layers III and V. The extent of the tangential connections from V2 varied from 2.5 to 3.0 mm (36). In Saimiri sciureus the areas of orientation in V2 formed granules; the size and distance between these granules were double those in V1 (37). In V2, in Macaca fascicularis (38) the results found were very similar to those ofS. sciureus(37). In the present study, we provide evidence that intrinsic horizontal and vertical connectivity from area V2 is capable of not only modulating the spontaneous and driven activity of V2 neurons but also modifying V2 receptive field properties, including orienta-tion and direcorienta-tion selectivity.
We propose the following hypothetical inactivation circuits to explain the results described in the present study. The intrinsic circuits modify the properties of the receptive field, probably through excitatory and inhibitory neurons. The most common effect observed in the first 5-10 min of GABA injection in V2 was a decrease in both spontaneous and driven activity of V2 neurons. We propose that pyramidal neurons that modulate directional selectivity in V2 contain GABAAreceptors (6) that capture
propagation of activity in the neurons that project to both the superficial and deep layers of V2. There is also a decrease in the release of the neurotransmitters from the neurons of area V2. As a result, the excitatory synapses decrease, causing a decrease in spontaneous and driven activity of the V2 neurons. The injections affect all direction selectivity columns, resulting in a decrease of spontaneous and driven activity of the neurons in all directions.
A loss of selectivity was the most frequently detected receptive field alteration in the V2 neurons after GABA inactivation. We hypothesize the existence of a circuit involving a projection of neurons (likely pyramidal neurons) from deep and superficial layers of area V2 containing GABAA receptors. The excitability of these
neurons would decrease after the activation of GABAA
receptors. This decrease in excitability would influence the pyramidal neurons in the V2 area that receive these projections and would also influence intrinsic inhibitory neurons. Intrinsic inhibitory interneurons decrease their influence on neuronal afferents to neighboring columns and cause a loss of direction selectivity for the majority of neurons. The directionality of the remaining 10% of neurons in our population became selective after the GABA injection. Therefore, we propose that the inactiva-tion of area V2 has partial and asymmetrical effects, which causes some direction columns to remain active while others become suppressed. This asymmetrical inhibition would generate direction selectivity in neurons that were pan-directional before the injection.
Optimization of the use of the animal model
We used one C. apella monkey in 5 recording sessions performed once a week to collect data from V2 neurons immediately before and after the injection of GABA. We aimed to record from as many cells as necessary in several experimental sessions in one animal
to have a statistically significant sample. The use of a single animal is justified to preserve as many primates as possible. Other studies that used only one animal but had statistically significant samples have been previously published (39,40).
GABA inactivation on the intrinsic, feed-forward and feedback circuits
We should consider the effect of GABA inactivation on the feed-forward, intrinsic and feedback circuits. There are four possible targets for the GABA injections in V2. The first is a direct effect on the recorded neuron. The second is an indirect effect involving an intrinsic V2 neuron that projects to the recorded neuron. The third is an indirect effect involving feed-forward projections, and the fourth is an indirect effect involving feedback projec-tions. The results obtained with small injections of GABA on the orientation and direction selectivity of V2 neurons are different from the results obtained with injections of GABA in the feedback loop of V2. While injections in V4 or MT increase the orientation and direction indices, direct injections in V2 decrease the orientation and direction indices. The results suggest that small amounts of GABA interfere only with intrinsic neurons that receive axonal terminals from feedback circuits.
The large injections of lidocaine produced a robust inhibitory effect on the activity of the V2 cells and they were used as control for the inhibitory component of GABA injections.
Acknowledgments
Research supported by PRONEX, CNPq, FUJB, FAPERJ, FINEP, and CEPG. A.K. Jansen-Amorim received CNPq fellowships for her graduate studies. R. Gattass is the recipient of a CNPq research fellowship.
References
1. Hubel DH, Wiesel TN. Receptive fields functional architec-ture of monkey striate cortex.J Physiol1968; 195: 215-243. 2. Lamme VA, Roelfsema PR. The distinct modes of vision offered by feedforward recurrent processing.Trends Neurosci 2000; 23: 571-579, doi: 10.1016/S0166-2236(00)01657-X. 3. Angelucci A, Levitt JB, Lund JS. Anatomical origins of the
classical receptive field modulatory surround field of single neurons in macaque visual cortical area V1.Prog Brain Res 2002; 136: 373-388, doi: 10.1016/S0079-6123(02)36031-X. 4. Douglas RJ, Martin KA. A functional microcircuit for cat
visual cortex.J Physiol1991; 440: 735-769.
5. Gilbert CD, Wiesel TN. Columnar specificity of intrinsic horizontal corticocortical connections in cat visual cortex.J Neurosci1989; 9: 2432-2442.
6. Sato H, Katsuyama N, Tamura H, Hata Y, Tsumoto T. Mechanisms underlying orientation selectivity of neurons in the primary visual cortex of the macaqueJ Physiol 1996; 494: 757-771.
7. Crook JM, Kisvarday ZF, Eysel UT. Evidence for a contribution of lateral inhibition to orientation tuning direction selectivity in cat visual cortex: reversible inactivation of functionally characterized sites combined with neuroanato-mical tracing techniques.Eur J Neurosci1998; 10: 2056-2075, doi: 10.1046/j.1460-9568.1998.00218.x.
8. Sillito AM. The contribution of inhibitory mechanisms to the receptive field properties of neurones in the striate cortex of the cat.J Physiol1975; 250: 305-329.
9. Sillito AM. The effectiveness of bicuculline as an antagonist of GABA visually evoked inhibition in the cat’s striate cortex. J Physiol1975; 250: 287-304.
10. Crook JM, Eysel UT. GABA-induced inactivation of func-tionally characterized sites in cat visual cortex (area 18): effects on orientation tuning. J Neurosci 1992; 12: 1816-1825.
(area 18): effects on direction selectivity. J Neurophysiol 1996; 75: 2071-2088.
12. Crook JM, Kisvarday ZF, Eysel UT. GABA-induced inactivation of functionally characterized sites in cat striate cortex: effects on orientation tuning direction selectivity. Vis Neurosci1997;14:141-158,doi:10.1017/S095252380000883X. 13. Sato H, Katsuyama N, Tamura H, Hata Y, Tsumoto T. Mechanisms underlying direction selectivity of neurons in the primary visual cortex of the macaque.J Neurophysiol 1995; 74: 1382-1394.
14. Rosa MG, Sousa AP, Gattass R. Representation of the visual field in the second visual area in theCebusmonkey.J Comp Neurol1988; 275: 326-345, doi: 10.1002/cne.902750303. 15. Hupe´ JM, Chouvet G, Bullier J. Spatial temporal parameters
of cortical inactivation by GABA.J Neurosci Methods1999; 86: 129-143, doi: 10.1016/S0165-0270(98)00162-9. 16. Gattass R, Oswaldo-Cruz E, Sousa APB. Visuotopic
organization of theCebuspulvinar: a double representation the contralateral hemifield.Brain Res1978; 152: 1-16, doi: 10.1016/0006-8993(78)90130-0.
17. Gattass R, Sousa APB, Oswaldo-Cruz E. Single unit response types in the pulvinar of the Cebus monkey to multisensory stimulation.Brain Res1978; 158: 75-87, doi: 10.1016/0006-8993(78)90007-0.
18. Gattass R, Oswaldo-Cruz E, Sousa APB. Visual receptive fields of units in the pulvinar ofCebusmonkey.Brain Res 1979; 160: 413-430, doi: 10.1016/0006-8993(79)91070-9. 19. Amorim AKJ, Picanco-Diniz CW. Morphometric analysis of
intrinsic axon terminals ofCebusmonkey area 17.Braz J Med Biol Res1996; 29: 1363-1368.
20. Amorim AKJ, Picanco-Diniz CW. Intrinsic projections of Cebus-monkey area 17: cell morphology axon terminals. Rev Bras Biol1996; 56 (Suppl 1): 209-219.
21. Amorim AKJ, Picanco-Diniz CW. Horizontal projections of area 17 inCebusmonkeys: metric features modular laminar distribution.Braz J Med Biol Res1997; 30: 1489-1501, doi: 10.1590/S0100-879X1997001200018.
22. Gattass R, Sousa APB, Rosa MG. Visual topography of V1 in theCebusmonkey.J Comp Neurol1987; 259: 529-548, doi: 10.1002/cne.902590404.
23. Fiorani M, Gattass R, Rosa MG, Sousa APB. Visual area MT in theCebusmonkey: location visuotopic organization variability.J Comp Neurol1989; 287: 98-118, doi: 10.1002/ cne.902870108.
24. Pin˜on MC, Gattass R, Sousa APB. Area V4 in Cebus monkey: extent visuotopic organization.Cereb Cortex1998; 8: 685-701, doi: 10.1093/cercor/8.8.685.
25. Gattass R, Nascimento-Silva S, Soares JG, Lima B, Jansen AK, Diogo AC, et al. Cortical visual areas in monkeys: location, topography, connections, columns, plasticity cor-tical dynamics.Philos Trans R Soc Lond B Biol Sci2005; 360: 709-731, doi: 10.1098/rstb.2005.1629.
26. Jansen-Amorim AK, Lima B, Fiorani M, Gattass R. GABA inactivation of visual area MT modifies the responsive-ness direction selectivity of V2 neurons in Cebus
monkeys. Vis Neurosci 2011; 28: 513-527, doi: 10.1017/ S0952523811000411.
27. Jansen-Amorim AK, Fiorani M, Gattass R. GABA inactiva-tion of area V4 changes receptive-field properties of V2 neurons inCebus monkeys.Exp Neurol2012; 235: 553-562, doi: 10.1016/j.expneurol.2012.03.008.
28. Soares JG, Diogo AC, Fiorani M, Souza APB, Gattass R. Effects of inactivation of the lateral pulvinar on response properties of second visual area cells inCebusmonkeys. Clin Exp Pharmacol Physiol 2004; 31:580-590, doi: 10.1111/j.1440-1681.2004.04051.x.
29. Wang C, Waleszczyk WJ, Burke W, Dreher B. Modulatory influence of feedback projections from area 21a on neuronal activities in striate cortex of the cat.Cereb Cortex2000; 10: 1217-1232, doi: 10.1093/cercor/10.12.1217.
30. Kisvarday ZF, Crook JM, Buzas P, Eysel UT. Combined physiological-anatomical approaches to study lateral inhibi-tion.J Neurosci Methods2000; 103: 91-106, doi: 10.1016/ S0165-0270(00)00299-5.
31. Hupe´ JM, James AC, Girard P, Bullier J. Response modulations by static texture surround in area V1 of the macaque monkey do not depend on feedback connections from V2.J Neurophysiol2001; 85: 146-163.
32. Martin JH, Cooper SE, Ghez C. Differential effects of local inactivation within motor cortex red nucleus on perfomance of an elbow task in the cat. Exp Brain Res 1993; 94: 418-428.
33. Maunsell JHR, Nealey TA, DePriest DD. Magnocellular parvocellular contributions to responses in the middle temporal visual area (MT) of the macaque monkey. J Neurosci1990; 10: 3323-3334.
34. Sillito AM. Proceedings: Modification of the receptive field properties of neurones in the visual cortex by bicuculline, a GABA antagonist.J Physiol1974; 239: 36P-37P.
35. Rockland KS. A reticular pattern of intrinsic connections in primate area V2 (area 18).J Comp Neurol1985; 235: 467-478, doi: 10.1002/cne.902350405.
36. Livingstone MS, Hubel DH. Specificity of intrinsic connec-tions in primate primary visual cortex.J Neurosci1984; 4: 2830-2835.
37. Malach R, Tootell RB, Malonek D. Relationship between orientation domains, cytochrome oxidase stripes, and intrin-sic horizontal connections in squirrel monkey area V2.Cereb Cortex1994; 4: 151-165, doi: 10.1093/cercor/4.2.151. 38. Levitt JB, Yoshioka T, Lund JS. Intrinsic cortical connections
in macaque visual area V2: evidence for interaction between different functional streams. J Comp Neurol 1994; 342: 551-570, doi: 10.1002/cne.903420405. 39. Lima B, Fiorani M, Gattass R. Modulation by context of a
scene in monkey anterior inferotemporal cortex during a saccadic eye movement task.An Acad Bras Cienc2003; 75: 71-76, doi: 10.1590/S0001-37652003000100008. 40. Lima B, Fiorani M, Gattass R. Changes of ongoing activity in