FOOD CONCENTRATION AND TEMPERATURE EFFECTS ON
LIFE CYCLE CHARACTERISTICS OF TROPICAL CLADOCERA
(Daphnia gessneri
Herbst,
Diaphanosoma sarsi
Richard,
Moina reticulata
(Daday)): I. DEVELOPMENT TIME.
E l s a . R. H A R D Y1 , 2
, A . D U N C A N2
ABSTRACT — The effects of food concentration and temperature on embryonic and postem-bryonic duration of three tropical species, Daphnia gessneri (1.5mm), Diaphanosoma sarsi (1.2mm) and Moina reticulata (0.8mm), were investigated as part of life cycle studies which included growth, body size and reproduction. These are the very first experimental studies un-dertaken on these species. The long-term growth experiments were performed under controlled laboratory conditions at all combinations of temperature (22"C, 27"C and 32"C) and constant food concentration (0.03, 0.05, 0.10, 0.25, 0.50 and 1.00 mgC/L) of the unicellular green alga Scenedesmus acutus. Animals were examined twice daily throughout their life cycle from the neonate to third adult instar. In all three species, temperature exerted the most powerful influ-ence on embryonic duration but there was also a smaller food effect. In D. gessneri, postembry-onic durations remained more or less the same at food levels 0.25 mgC/L but were influenced by temperature. At food concentrations of 0.1 mgC/L or lower, postembryonic durations became increasingly prolonged, particularly at high temperatures. This threshold concentration is affected by temperature: in D. gessneri, it was 0.1 mgC/L at 22o
C and 27o
C but higher at 32o
C (between 0.25 and 0.50 mgC/L). At the same temperature of 27o
C, the food threshold level varied be-tween species: it was higher (0.25 mgC/L) for D. sarsi and lower (0.05 mgC/L) for M. reticulata compared with D. gessneri (0.1 mgC/L). In both embryonic and postembryonic durations there is a body size effect as the absolute durations were longest in the largest species and shortest in the smallest species In all three species, prolongation of postembryonic duration at combina-tions of high temperature and lowered food levels was accompanied by increased number of ju-venile instars.
Key- words: Daphnia gessneri, Diaphanosoma sarsi, Moina reticulata; Tropical; Food concen-tration-temperature experiments; Embryonic(Dc ) ; Postembryonic development (Dj) Ratio (DJ Dc ); Food Threshold.
Efeitos da Temperatura e Concentração de Alimento na Historia de Vida de Tropical Cladocera (Daphnia Gessneri Herbst,Diaphanosoma Sarsi Richard, Moina reticulata Daday,)) I. Tempo de desenvolvimento.
R E S U M O — O s efeitos d e t e m p e r a t u r a e c o n c e n t r a ç ã o de a l i m e n t o na d u r a ç ã o do desenvolvimento embrionário e postembrionário de três espécies tropicais, Daphnia gessneri (1.5mm), Diaphanosoma sarsi (1.2mm) and Moina reticulata (0.8mm), foram investigados como parte dos estudos de ciclo de vida incluindo crescimento, tamanho do corpo e reprodução. Estes são os primeiros estudos experimentais realizados com estas espécies. Os experimentos de crescimento de longa duração foram feitos em condições controladas de laboratório em todas as combinações de temperature (22°C, 27°C e 32°C) e concentração constante de alimento (0.03, 0.05,0.10,0.25, 0.50 and 1.00 mgC/L) de alga verde unicelular Scenedesmus acutus. Os animais foram analisados duas vezes por dia durante o ciclo de vida desde neonata ao terceiro estádio
1 National Institute of Amazon Research (INPA), Department of Aquatic Biology, Manaus, Brazil, CEP 69011.
adulto. Nas três espécies, a temperatura exerceu um maior efeito no tempo do desenvolvimento embrionário mas ocorreu também um pequeno efeito de concentração de alimento. Em Daphnia gessneri, a duração postembriônica permaneceu mais ou menos a mesma na concentração de alimento de 0.25 mgC/L mas foi influenciada pela temperature. Na concentração de 0.1 mgC/L ou mais baixa, a duração postembrionária tornou-se mais prolongada, particularmente em alta temperatura. O valor absoluto do nivel crítico de concentração de alimento foi afetado pela temperatura: em D. gessneri, foi 0.1 mgC/L em 22°C e 27°C mas aumentou em 32°C (entre 0.25 e 0.50 mgC/L). Na mesma temperatura de 27°C, o nivel crítico de alimento variou entre as espécies: foi mais alta (0.25 mgC/L) para D. sarsi e mais baixa (0.05 mgC/L) para M. reticulata comparada com D. gessneri (0.1 mgC/L). Em ambos, no desenvolvimento embrionário e postembrionário ocorreu o efeito do tamanho do corpo visto que as durações absolutas foram mais longas nas espécies maiores e mais curtas.na espécie menor. Nas três espécies, o prolongamento do desenvolvimento postembrionário nas combinações de alta temperatura e menores concentrações de alimento foi acompanhado por aumento no número de estádios juvenis. Palavras c h a v e s : Daphnia gessneri, Diaphanosoma sarsi, Moina reticulata; Tropical; Experimentos; Temperatura e concentração de alimento; Embrionico ( D ) ; Postembrionico (D.); Ratio ( D / D ) ; Nível crítico de alimento.
INTRODUCTION
It has been reported m a n y t i m e s that t h e duration of e m b r y o n i c d e v e l -o p m e n t in c r u s t a c e a n s is a functi-on -of temperature only ( I N G L E et al., 1937; E S S L O V A , 1 9 5 9 ; H A L L , 1 9 6 4 ; K O R I N E K , 1 9 7 0 ; M U N R O & W H I T E , 1975; review in B O T T R E L L et al., 1 9 7 6 ; M A G A D Z A , 1 9 7 7 ; L E V E Q U E & S A I N T - J E A N , 1 9 8 3 ; H E R Z I G , 1984), w h e r e a s the duration of p o s t - e m b r y o n i c d e v e l o p m e n t is in-f l u e n c e d b y in-f o o d l e v e l a s w e l l ( H R B A C K O V A - E S S L O V A , 1 9 6 3 ; W E G L E N S K A , 1 9 7 1 ; K O R I N E K , 1971). M a n y a u t h o r s h a v e s h o w n that cladocerans cultured at high food con-centrations h a v e a shorter duration of p o s t - e m b r y o n i c d e v e l o p m e n t t h a n those cultured at low food levels ( D E B E R N A R D I et al, 1978; G R A S & S A I N T J E A N , 1 9 7 8 ; L E I & A R M I -T A G E , 1 9 8 0 ; V I J V E R B E R G , 1980; K A N K A A L A & W U L F F , 1 9 8 1 ; R O C H A , 1 9 8 3 ; O R C U T T & P O R -T E R , 1 9 8 4 ; J A Y A -T U N G A , 1 9 8 6 ) .
M o s t of the published information is f o r t e m p e r a t e s p e c i e s o n l y , l a r g e l y Daphnia, and there are few q u a n t i t a -tive e x p e r i m e n t a l studies on tropical species such as those of J A Y A T U N G A ( 1986) w h o investigated d e v e l o p m e n t t i m e s of Sri L a n k a n c l a d o c e r a n s . A u -thors w o r k i n g with tropical planktonic cladocerans such as G R A S & S A I N T -J E A N ( 1 9 7 8 ) , M U R U G A N ( 1 9 7 5 ) , J A N A & P A L ( 1 9 8 4 , 1 9 8 5 ) a n d S A I N T - J E A N & B O N O U ( 1 9 9 4 ) fed their experimental a n i m a l s with u n d e -fined lake or pond water. Quantitative studies a r e important as d e v e l o p m e n -tal d u r a t i o n s p r o v i d e the t i m e e l e m e n t in g r o w t h and production d e t e r m i n a -tions. It is of interest to k n o w w h e t h e r t h e largely s m a l l s i z e d t r o p i c a l c l a d o c e r a n s h a b i t u a t e d to n a r r o w t e m perature ranges respond like w a r m e d -u p t e m p e r a t e forms.
c o m b i n a t i o n , using species c o m m o n l y found in the A m a z o n " v á r z e a " lakes: Daphnia gessneri, Moina reticulata and Diaphanosoma sarsi, with third instars adult sizes of 1.5mm, 0.8mm a n d 1.2mm. T h e s e are the first such e x p e r i m e n t a l studies on these species and in Latin America. T h e experimen-tal t e m p e r a t u r e s ranged from 22-32"C - high t e m p e r a t u r e s for temperate species but s p a n n i n g the full range e x p e -rienced by tropical f o r m s . F o o d lev-e l s w lev-e r lev-e s lev-e l lev-e c t lev-e d to c o v lev-e r a l i k lev-e l y r a n g e from l i m i t i n g to n o n - l i m i t i n g food c o n c e n t r a t i o n s in t h e l i g h t of J A Y A T U N G A ' s ( 1 9 8 6 ) p r i o r e x p e r i -e n c -e with Sri L a n k a n c l a d o c -e r a n s .
MATERIALS AND METHODS
Stock cultures
T h e c l a d o c e r a n s p e c i e s studied w e r e collected from L a k e Jacaretinga in the A m a z o n State, Brazil, and trans-p o r t e d t o R o y a l H o l l o w a y C o l l e g e w h e r e they w e r e m a i n t a i n e d in 1 litre-b e a k e r c u l t u r e s u s i n g f i l t e r e d Alderhurst P o n d water at 27"C, in an Astell H e a r s o n incubator u n d e r a c o n -trolled 12 h o u r d a y / 1 2 hour dark reg i m e . T h e s e s t o c k c u l t u r e s of c l a -d o c e r a n s w e r e fe-d from stock cultures of t h e u n i c e l l u l a r g r e e n a l g a , Scenedesmus acutus ( C h l o r o p h y c e a e , C h l o r o c o c c a l e s ) w h i c h w e r e m a i n -t a i n e d in -t h e e x p o n e n -t i a l p h a s e of g r o w t h by subculturing. In this c o n dition, the food alga had cell d i m e n -sions of 10.34 χ 4.68 μ πι, cell volume of 102 μ πι3
· and cell carbon content of 11.78 p g C ( R O C H A & D U N C A N , 1985). F o r a b o u t three w e e k s prior to
an e x p e r i m e n t a l r u n , t h e s t o c k cla d o c e r a n c u l t u r e s w e r e fed daily at a food level of 1.0 m g C / L .
Preparation of experimental
food
S a m p l e s of Scenedesmus acutus
taken from t h e 59 day old liquid m e dium culture in e x p o n e n t i a l p h a s e of growth w e r e centrifuged to s e d i m e n t the cells and w a s h e d twice in distilled water by re centrifugation and resus p e n s i o n to r e m o v e t h e n u t r i e n t m e d i u m . T h e carbon content of the final concentrate of algae collected quanti tatively w a s d e t e r m i n e d by the wet o x i d a t i o n t e c h n i q u e ( C O D ) ( M A C K E R E T H et ai, 1978). F o o d m e d i u m with a c o n c e n t r a t i o n of 1.0 m g C / L was prepared by adding appro priate a m o u n t s of GFFfiltered p o n d water to the stock food suspension and other l o w e r food levels w e r e prepared by careful dilution of the 1.0 m g C / L s u s p e n s i o n .
Experimental chambers
Figura 1. Continuons flow system food concentration and temperature effects on the life cycle characteristics of tropical cladocera Daplinia gessneri Herbst, Diaphanosoma sarsi Richard, Moina reticulata (Daday): I Development time. E.R. HARDY & A. DUNCAN.
i m m e r s e d in a temperaturecontrolled waterbath w h i c h maintained both the animals and t h e p u m p e d food supply at the e x p e r i m e n t a l t e m p e r a t u r e u n d e r natural daylight c o n d i t i o n s . T h e other kind of experimental s y s t e m adopted was batch culture using 2 5 0 ml bottles w h o s e f o o d m e d i u m w a s c h a n g e d daily. T h e s e bottles w e r e turned on t h e i r s h o r t a x e s at I r p m i n s i d e an Astell H e a r s o n i n c u b a t o r w i t h light and t e m p e r a t u r e control. E a c h bottle c o n t a i n e d o n e i n d i v i d u a l a n d t h e r e w e r e 4 replicates.
Experimental design and
procedure
Daphnia gessneri was reared at three temperatures (22°, 27" and 32"C) a n d f i v e f o o d c o n c e n t r a t i o n s of
Scenedesmus acutus ( 1 . 0 , 0 . 5 , 0 . 2 5 ,
0.1 and 0.05 m g C / L ) , g i v i n g 15 ex perimental conditions; Diaphanosoma sarsi and Moina reticulata w e r e cul tured at the s a m e food levels but only o n e temperature (27"C). An additional food level ( 0 . 0 3 m g C / L ) w a s tested in
M. reticulata.
s u r e m e n t of length, d e t e r m i n a t i o n of instar stage ( c a r a p a c e s ) and counting the n u m b e r of e g g s or e m b r y o s in the b r o o d p o u c h e s of adults. T h e duration of t h e a d u l t i n s t a r s g a v e e m b r y o n i c durations. A n i m a l s w e r e replaced into a clean f l o w t h r o u g h c h a m b e r filled with freshly p r e p a r e d food m e d i u m . With experienced handling, mortalities w e r e a l m o s t nil a m o n g s t wellfed ani mals at the h i g h e r food concentrations but attained levels of 3 0 % (Daphnia)
to 4 0 % (Moina) at the lowest food lev e l s ( 0 . 0 5 m g C / L ) m o s t l y d u r i n g e c d y s i s w h e n a n i m a l s w e r e especially vulnerable. A n y accidental deaths or l o s s e s of a n i m a l s w e r e not replaced in flow c u l t u r e s but n e o n a t e s w e r e re placed in batch cultures.
RESULTS
Embryonic development
Table 1 gives the mean durations of e m b r y o n i c development in the three cladoceran species at e a c h of the ex p e r i m e n t f o o d t e m p e r a t u r e c o m b i n a
tions in flow c u l t u r e . T h e s e m e a n s , which w e r e calculated from between 612 o b s e r v a t i o n s of all the individu als ( u p to 20) in o n e flow c h a m b e r , represent the t i m e from release of o o cytes from the o v a r y into t h e b r o o d p o u c h to t h e r e l e a s e of first instars from the brood p o u c h , a s detected by twice daily o b s e r v a t i o n .
In D. gessneri w i t h t h e m o s t c o m p l e t e data set, it is clear that t e m p e r a t u r e h a s a p o w e r f u l i n f l u e n c e , d o u b l i n g the e m b r y o n i c duration with a 10°C decrease in temperature. T h e r e is also a food effect which results in a less p r o n o u n c e d prolongation of e m b r y o n i c d u r a t i o n as food levels b e c o m e m o r e l i m i t i n g . M o r e o v e r , the absence of Dc values for 0.05 m g C / L
at 27°C and for 0.10 and 0.05 m g C / L at 32°C is b e c a u s e all t h e j u v e n i l e s died before maturing to an adult with adult instars. T h e shortest d u r a t i o n occurred at 32"C and 1.0 m g C / L and the m o s t p r o l o n g e d e m b r y o n i c dura tion at 22"C and 0.25 m g C / L . T h e r e is a s i m i l a r food effect on e m b r y o n i c
Table 1. The duration of the embryonic development ( D ) of Daphnia gesseneri, Moina reticulata and Diaphanosoma sarsi in various combinations of temperature and food concentration in flow culture Duration in hours; mean±Standart deviation; number of observations in brackets.
TEMPERATURE
°C 1,0
FOOD CONCENTRATION (mgC.L:1 )
0,5 0,25 0,1 0,05 0,03
Daphnia Gessneri
22 50±4,6(12) 56±10.6{12) 63±11,5(12) 58±16,3(9) 54±10,0{7) 27 40±14,7(12) 38±16,0{12) 54±10,8{12) 54±16,9(12)
32 24±0,0(12) 28±9,7(12) 31±11,5 (12) Moina Reticulata
27 24±0,0(12) 24±0,4(12) 24±0,4(12) 25±3,4(12) 31±7,0{3) 51*2,5(7) Diaphanosoma Sarsi
27 24±0,0{12) 29±10,4(9) 34±12,4(9) 40±11,4(6) 44±20,4(11) * The females did not complete three broods.
duration in Μ. reticulata and D. sarsi
(Table 1). T h e e m b r y o n i c duration of
Moina at food levels 1.0 to 0.25 m g C / L w a s c o n s t a n t at 2 4 h o u r s and w a s both m u c h s h o r t e r a n d less variable than t h o s e of t h e o t h e r t w o s p e c i e s (Table 1).
A c o m p a r i s o n of the three spe cies at o n e t e m p e r a t u r e s h o w s consid e r a b l e v a r i a t i o n in e m b r y o n i c d u r a tions at optimal food levels, that of the largest species (Daphnia) being m u c h l o n g e r than the t w o s m a l l e r s p e c i e s . At the m o s t s e v e r e food level of 0.05 m g C / L , the durations vary with spe cies size (54 hr, 1.5mm; 4 4 hr, 1.2mm; 31 hr, 0 . 8 m m ) (Table 1).
Table 2 gives the results of fit ting a c u r v i l i n e a r r e g r e s s i o n of t h e form D = a . Th
to d e s c r i b e , for Daphnia gessneri only, the relationship between e m b r y o n i c d e v e l o p m e n t ( D in hours) and temperature (T in degrees Celsius) for each tested food level. Statistically significant regressions w e r e obtained for food levels from 1.00 to 0.25 m g C / L but not for 0.1 m g C / L ( P = 0 . 2 5 ) or
for 0.05 m g C / L in which the life cycle w a s c o m p l e t e d u n d e r flow conditions at 22°C only (Table 1). Table 3 gives the results of c o v a r i a n c e a n a l y s i s of t h e t h r e e s i g n i f i c a n t r e g r e s s i o n s in Table 2. T h i s s h o w s that the regres sion coefficients (slopes) w e r e not sig nificantly different but that the eleva tion of the regression for 0.25 m g C / L w a s significantly l o w e r than the other t w o higher food levels.
An attempt w a s m a d e to fit simi lar curvilinear regressions to quantify the r e l a t i o n s h i p b e t w e e n e m b r y o n i c d e v e l o p m e n t and food c o n c e n t r a t i o n for each tested t e m p e r a t u r e a n d s p e cies. Only one significant relationship w a s f o u n d , t h a t for Diaphanosoma sarsi: lnY = 3.19 0.1791nX (df= 1,42; F = 1 3 0 ; P = 0 . 0 0 1 ) .
Post-embryonic development
T h e duration of p o s t e m b r y o n i c d e v e l o p m e n t ( = D . j u v e n i l e d u r a t i o n ) w a s taken to extend from the release of n e o n a t e s from the f e m a l e ' s brood pouch (time 0) to the first a p p e a r a n c e
Table 2. Curvilinear regressions relating the duration of embryonic development to temperature at various food concentrations for Daphnia gessneri in flow culture.
R e g r e s s i o n e q u a t i o n lny = Ina b In X
Y = duration of embryonic development in hours; X = temperature in "C df = degrees of freedom; F = variance ratio; Ρ = level of significance FOOD LEVEL (MgC. L1
) In a b df F Ρ
Daphnia gessneri
1,0 9,76 -1,88 1,29 27,9 0.001
0,5 9,86 -1,90 1,32 87,6 0.001
0,25 10,03 -1,88 1,31 5,7 0.025
0,1 8,87 -1,54 1,18 2,6 0 . 2 5
0,05 *
Table 3 . Covariance analysis of the significant regressions of duration of embryonic develop ment on temperature at various food levels for Daphnia gessneri. The regression coefficients were compared by the SS-STP test and the differences between elevations by the S-N-K test. Regression coefficients and means underlined are not significantly different at P=0.05 level; Group numbers are given in ascending order of magnitude.
FOOD LEVEL GROUP REGRESSION COEF.±SE mgC.L1
1,0 1 -1.88±0.30 0,5 2 -1.90±0.33 0,25 3 -1.88±0.33
mgC.L1
GROUP ADJUSTED MEAN+SE 1,0 1 3.59±0.15 0,5 2 3.64±0.15 0,25 3 3.85±0.15
Comparisons of S l o p e s df F
2,95
Copararison of Elevations
df
2,98 F
8.61
S S - S T P
0.001 0 , 9 9 1 2 3
Ρ
0.000 S-N-K 1 2 3
of the p r i m i p a r o u s female carrying her first b r o o d . T h i s w a s easiest to o b s e r v e on i n d i v i d u a l s in batch c u l t u r e . T h u s the d u r a t i o n of the j u v e n i l e stage a l s o e s t i m a t e s the a g e of the p r i m i p a r a fe
m a l e . In practice, the actual m o m e n t of release of n e o n a t e s or o o c y t e s w a s o n l y r a r e l y o b s e r v e d b u t w e r e r e c o r d e d at the next o b s e r v a t i o n t i m e . T h e p r e s e n c e of an e c d y s e d c a r a p a c e
Table 4. Duration of post-embryonic development (hours) to primipara and of instars (in paren thesis) attained in various combinations of food and temperature in, Daphnia gessneri. Moina reticulata and Diaphanosoma sarsi. Ν = 4. Batch culture.
TEMPERATURE °C 1,0
FOOD CONCENTRATION (mgC.L' )
0,5 0,25 0,1 0 , 0 5 0,03 Daphnia gessneri
22
27
32 Moina reticulata
27
Diaphanosoma sarsi 2 7
- not tested
177±11.4 (V-VII) 1 7 4 ±1 2 . 0
(V-VII) 132±0.0
(IV-VI)
54.0±0.0
186±12.0 (VI-VII) 156±16.9
(V-VII) 156±0.0
(V.VI)
48.0±0.0 (III)
186±20.7 (VIII) 162±12.0
(VI-VII) 162±12.0
(V-VI)
2 0 7 ±3 5 . 8 (VIII) 192±0.0 (VIII-IX) 2 0 7 ±1 8 . 0
(VI-VII)
48.0±0.0 48.0±0.0
72.0±13.8 72.0±13.8 90.0±12.0 116.0±13. (III-IV) (IV) (IV-V) (V)
2 4 0 ±5 6 . 7 (Vll-X) 198±22.8
(IX) 2 4 6 ±5 3 . 2
(VII-VIII)
6 0 . 0 . ±0 . 0 90.0±12.0 (III-IV) (IV-V)
a c c o m p a n i e s t h e r e l e a s e of the first instar n e o n a t e s and p e r m i t s the iden tification of the instar stage of t h e fe male.
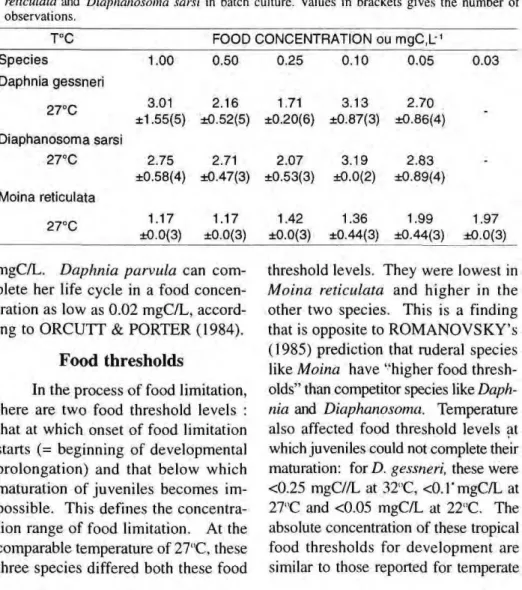
Table 4 g i v e s t h e m e a n p o s t e m bryonic d u r a t i o n s (± S D ) for the three s p e c i e s at t h e s a m e t e m p e r a t u r e and food level c o m b i n a t i o n s as before but taken from t h e b a t c h culture experi ments. In D. gessneri, there is a gen
250-,
eral pattern of p r o l o n g a t i o n of post embryonic development with d e c r e a s e in temperature, with s o m e variability at t h e higher food levels, in Table 5, t h r e e significant c u r v i l i n e a r r e g r e s sions of duration on temperature could be fitted to the data for t h e top food levels but not for 0.1 and 0.05 m g C / L. C o v a r i a n c e analysis in Table 6 of t h e t h r e e s i g n i f i c a n t r e g r e s s i o n s
200-
150-
100-Oaphnia
..22°C •27°C
•~.32°C
50-Moma
27° C
-o27°C
03 FOOD LEVEL (mg CI
TO
Figura 2. The effect of food concentration on the mean duration of postembryonic development
s h o w e d t h a t n e i t h e r t h e s l o p e s ( P = 0 . 2 4 ) n o r t h e elevations (P=0.26) of t h e s e differed. T h i s m e a n s that t e m p e r a t u r e influences juvenile dura tion in Daphnia gessneri in the higher food levels but not in the lower ones.
T h e striking effect in Table 4 is the prolongation of the postembry onic d u r a t i o n in the l o w e r food lev els from 0.1 m g C / L downwards, in D. gessneri, f r o m 0 . 2 5 m g C / L d o w n wards in D. sarsi and from 0.05 m g C / L d o w n w a r d s in M. reticulata. Table 7 gives t h e results for fitting curvilin ear regression of j u v e n i l e duration on food concentration at each tested tem perature. It p r o v e d possible to fit sig nificant regressions for D. gessneri at all three temperatures and for D. sarsi
at 2 7 " C b u t n o t f o r M. reticulata
( P = 0 . 1 4 ) . T h i s last species s h o w s a rather different pattern of r e s p o n s e with a c o n s t a n t j u v e n i l e duration of 4 8 hours for food levels from 0.5 to 0.1 m g C / L (Table 4 ) . However, a sig nificant regression could be fitted to the d a t a for food levels 0 . 1 , 0.05 and
0 . 0 3 m g C / L ; this w a s l n Y = 1.26 1.081nX (d.f. 1,7; F = 2 4 9 ; P = < 0 . 0 0 1 ) . Figure 2 illustrates these relationships between D and food concentration in the
ι
three cladocerans and shows several inter esting features. O n e is a body size effect, namely that postembryonic duration was longest in the largest species D. gessneri
(3rd instar adult size, 1.5mm), shortest in the smallest s p e c i e s M. reticulata
( 0 . 8 m m ) and w a s i n t e r m e d i a t e in D. sarsi ( 1 . 2 m m ) . T h i s figure also s h o w s that the food threshold level at w h i c h food limitation starts (= marked prolon gation of D.) is m u c h lower in Moina
(<0.1 mgC/L), higher in Diaphanosoma
(0.25 m g C / L ) than in Daphnia (at 0.1 m g C / L ) in a m a n n e r w h i c h parallels e m b r y o n i c d u r a t i o n . T h e a b e r r a n t p o i n t at 27°C/1.0 m g C / L in F i g u r e 2 c o m e s from a high d e g r e e of variabil ity in individual D. values and general l o w e r "fitness" ("difficult" ecdysis) in what s e e m s to be " e x c e s s " food for this species.
Table 4 also gives the instar stage o f the p r i m i p a r o u s f e m a l e s for e a c h
Table 5. Curvilinear regressions relating the duration of post-embryonic development to
tem-perature at various foog concentration for Daphnia gessneri in batch culture.
Regression equation 1 nY = 1 η a - 1 η X
Υ = post-embryonic development in hours; X = temperature in °C df = degrees of freedon; F = variance ratio; Ρ level of significance
FOOD CONCENTRATION
(mgC.L1
)
Daphnia gessneri
1
0,5
0 , 2 5
0,1
0,05
In a b df F Ρ
7,55 - 0 , 7 5 1,10 1 9 , 7 5 0.001
6 , 6 7 - 0 , 4 7 1,10 9,96 0 . 0 1 0
6,34 - 0 , 3 7 1,10 4,72 0 . 0 5
5,25 0,01 1,8 0 , 0 0 6 0 . 9 4
temperaturefood condition. T h e gen eral pattern of response in D. gessneri
is that the females b e c a m e primiparous at instar I V V at the h i g h e r t e m p e r a tures and h i g h e r food levels but were not able to p r o d u c e a b r o o d of y o u n g until instars V I I I I X X at lower tem peratures and low food levels. In both
Diaphanosoma and Moina at 2 7 " C , primiparous females appeared at instar III in t h e h i g h e s t food levels but at later and later instars with declining food levels. Diaphanosoma matured at i n s t a r s V V I in 0 . 0 5 m g C / L a n d
Moina at i n s t a r I V in 0 . 0 3 m g C / L . Only in Moina, the pattern of response differed in that there w a s n o c h a n g e in the duration of p o s t e m b r y o n i c devel o p m e n t until they w e r e reared in food levels of 0.050.03 m g C / L . In general, the r e s p o n s e to s e v e r e c o n d i t i o n s of both high t e m p e r a t u r e and low food level a p p e a r s to i n v o l v e d e l a y i n g the p r i m i p a r a ' s stage of d e v e l o p m e n t and, inevitably, increasing h e r a g e .
Table 8 gives values of the ratio for D./Dc for all three species reared in
batch culture at the comparable tempera ture of 27°C. T h e pattern s h o w n by
Moina reticulata is the simplest the ratio increases with greater prolongation of D. as food levels became limiting and, as shown in Table 4, this is associated with an increased n u m b e r of juvenile instars from III to V. T h e pattern of D7 Dc with food level is m o r e complex in
the other t w o species: the lowest ratio occurs at 0.25 m g C / L and it increases on either side of this food level. T h e increased ratios at food levels lower than 0.25 m g C / L are d u e to food limitation,
t o g e t h e r w i t h a s s o c i a t e d i n c r e a s e d n u m b e r o f j u v e n i l e i n s t a r s , a s in
Moina: from V to I X in Daphnia and from IV to VI in Diaphanosoma. We d o not k n o w the c a u s e s of the larger ratios in food levels larger than 0.25 m g C / L in t h e s e t w o s p e c i e s , e x c e p t that these are not associated with in c r e a s e d n u m b e r of j u v e n i l e i n s t a r s (Table 4 ) . In general, the replicated individuals reared in the food levels of 0.5 and 1.0 m g C / L were m o r e variable in D and D and less "fit".
T h e relationship of p o s t e m b r y onic duration to both t e m p e r a t u r e and food concentration w a s e x a m i n e d for
D. gessneri by m e a n s of m u l t i p l e re gression analysis (Table 9). The best fit to the d a t a w a s obtained using the equation 1/D. = a b . 1/T + c.lnF. An a n a l y s i s of t h e v a r i a n c e a s s o c i a t e d w i t h f o o d c o n c e n t r a t i o n w a s t h r e e times greater than that d u e to tempera ture.
DISCUSSION
T h e three species studied are all truly tropical (Lake Jacaretinga, 3"S).
Daphnia gessneri w a s first described in 1965, has b e e n recorded only from n o r t h e r n S o u t h A m e r i c a a n d is t h e only Daphnia species recorded in the wellworked L a k e J a c a r e t i n g a and its o t h e r l o c a l i t i e s . B o t h Moina a n d
Diaphanosoma are c o m m o n e r genera in the South A m e r i c a , particularly in the turbid A m a z o n i a n lakes, but little is k n o w n about their distribution else w h e r e .
portant factor as t e m p e r a t u r e for the d u r a t i o n of d e v e l o p m e n t of tropical p l a n k t o n i c c l a d o c e r a n s as of temper ate species. T h e r e is a significant pro l o n g a t i o n of d e v e l o p m e n t as e i t h e r t e m p e r a t u r e or food level decreases as well as t e m p e r a t u r e f o o d synergistic effects reported by O R C U T T & P O R T E R , ( 1 9 8 4 ) . T h e s e effects are shown for D. gessneri, Diaphanosoma sarsi
and Moina reticulata for the first time.
Embryonic duration
Temperature w a s the main factor controlling the e m b r y o n i c duration of
Daphnia gessneri reared on the higher food levels, with a halving of the du ration with a 10"C increase in tempera ture (Table 1). However, at lower food l e v e l s t h e r e is s o m e e v i d e n c e of a
f o o d l e v e l e f f e c t o n D , s h o w n by longer D , v a l u e s which c a u s e d n o n significant regressions of D , on t e m perature or inability to m e a s u r e Dc in
tested food levels u n d e r flow c o n d i tions (Tables 1, 2 ) . Both of these ef fects m a y b e d u e to t h e p o o r nutri tional condition of the m o t h e r under food limiting condition, particularly at t h e t w o h i g h e r t e m p e r a t u r e s . O R C U T T & P O R T E R ( 1 9 8 4 ) , w h o quantified their experimental food lev e l s ( 0 . 0 2 to 2.0 m g C / L ) , h a v e a l s o found a significant foodlevel effect as well as synergistic t e m p e r a t u r e f o o d effects on e m b r y o n i c d e v e l o p m e n t of the Georgian (USA) Daphnia parvula.
C o m p a r e d with Daphnia, t h e food level effect on Dc of Moina reticulata
a n d Diaphanosoma sarsi w a s b o t h
Table 6. Covariance analysis of the regressions comparing the duration of post-embryonic de-velopment on temperature at different food concentrations for Daphnia gessneri. The regres-sion coefficient were compared by the SStp test and difference between elevations by the
S-N - K test. Regression coeficients and mens underlined are not significantly different at Ρ = 0.05
level.
Regression equation InY = Ina - b InX
Y = duration of post-embryonic development in hous; X = food concentration in mgC.L ';
df = degrees of freedom; F = variance ratio; Ρ = level of significance
C O M P A R I S O N " O F S L O P E S
FOOD LEVEL mgC.L-1
1.0
0.5
0.25
mgC.L1
1.0
0.5
0.25
GROUP REGRESSION COEFF+SE
1 -0.75±0.17
2 -0.47±0.15
3 -0.37+0.17
GROUP ADJUSTED MEAN±SE
1 5.07±0.05
2 5.10±0.05
3 5.13±0.05 df
2,30 F
1,45
Ρ SS-STP
0,24 3 2 1
COMPARISON
OF ELEVATIONS
df F
2,33 1.39
Ρ
0.26
S-N-K
Table 7. Curvilinear regressions relating the duration of post-embryonic development to food
concentration at various temperature for Daphnia gessneri, Moina reticulata and Diaphanosoma
sarsi in batch culture
Regression equation InY = Ina - b InX
Y = post-embryonic development in hours; X = food concetration in mgC.L"'
TEMPERATURE °C In a b df F Ρ
Daphnia gessneri
2 2 5 . 1 4 - 0 . 0 8 7 1,18 8 . 6 3 0 . 0 0 8
2 7 5 . 0 7 - 0 . 0 6 0 1,16 6 . 8 9 0 . 0 1 8
3 2 4 . 8 9 - 0 . 1 6 0 1,18 5 0 . 7 0 0 . 0 0 0
Moina reticulata
2 7 4 3 . 3 8 - 8 . 0 5 1,18 3 . 2 2 0 . 1 4
Diaphanosoma sarsi
2 7 4 . 2 0 -0.21 1,17 4 5 . 1 0 . 0 0 0
larger and started at lower food levels (Table 1). J A Y A T U N G A ( 1 9 8 6 ) , w h o is the only o t h e r w o r k e r on tropical cladocerans u s i n g defined food levels, found a s i g n i f i c a n t p r o l o n g a t i o n of e m b r y o n i c duration in the Sri Lankan
Moina micrura at 0.05 m g C / L ) in both 27"C and 32«'C).
Postembryonic duration
T h e d e p e n d e n c e of j u v e n i l e d u ration on food concentration has been shown for all three cladoceran species but h o w t e m p e r a t u r e affects this re sponse could be demonstrated only for
Daphnia.
The pattern of response in Daph-nia gessneri is that the j u v e n i l e phase prolongation was c o n t i n u o u s as food level declined, w a s greatest at the t w o lower food levels and most severe at the highest temperature. T h i s pattern of Dj o n food level w a s statistically significant at each t e m p e r a t u r e (Table 7), there is n o statistical difference be tween regressions for D( on t e m p e r a
ture (Table 6) and the multiple r e g r e s sion in Table 9 shows that the variance associated with food concentration was three times greater than that due to tem perature. The most severe food limita tion occurred in the combination of low food and high temperature which repre sents extreme conditions for a tropical spe cies whose normal temperature range in the field is a few degrees either side of 27"C.
A similar pattern of response to that of Daphnia described above was shown by
Table 8. The mean values of the ratio Dj/ Dc and standard deviation for Daphnia gessneri, Moina reticulata and Diaphanosoma sarsi in batch culture. Values in brackets gives the number of observations.
T°C FOOD CONCENTRATION ou mgC.L*1
S p e c i e s 1.00 0.50 0.25 0 . 1 0 0 . 0 5 0 . 0 3
Daphnia g e s s n e r i
2 7 ° C 3.01 2 . 1 6 1.71 3 . 1 3 2 . 7 0
2 7 ° C
±1.55(5) ±0.52(5) ±0.20(6) ±0.87(3) ±0.86(4)
D i a p h a n o s o m a sarsi
2 7 ° C 2 . 7 5 2.71 2 . 0 7 3 . 1 9 2 . 8 3
-±0.58(4) ±0.47(3) ±0.53(3) ±0.0(2) ±0.89(4)
Moina reticulata
2 7 ° C 1.17 1.17 1.42 1.36 1.99 1.97
2 7 ° C
±0.0(3) ±0.0(3) ±0.0(3) ±0.44(3) ±0.44(3) ±0.0(3)
m g C / L . Daphnia parvula can c o m plete her life c y c l e in a food c o n c e n tration as low as 0.02 m g C / L , accord ing to O R C U T T & P O R T E R ( 1 9 8 4 ) .
Food thresholds
In the process of food limitation, t h e r e are t w o food threshold levels : that at w h i c h onset of food limitation starts (= b e g i n n i n g of d e v e l o p m e n t a l p r o l o n g a t i o n ) and that b e l o w w h i c h maturation of j u v e n i l e s b e c o m e s im possible. This defines the concentra tion r a n g e of food limitation. At the c o m p a r a b l e temperature of 27"C, these three s p e c i e s differed both these food
threshold levels. T h e y w e r e lowest in
Moina reticulata and h i g h e r in t h e o t h e r t w o species. T h i s is a finding that is opposite to R O M A N O V S K Y ' s (1985) prediction that ruderal species like Moina h a v e "higher food thresh olds" than competitor species like Daph-nia and Diaphanosoma. Temperature also affected food threshold levels at which juveniles could not complete their maturation: for D. gessneri, these were <0.25 mgC//L at 32°C, <0.rmgC/L at 27"C and <0.05 m g C / L at 22°C. The absolute concentration of these tropical food t h r e s h o l d s for d e v e l o p m e n t are similar to those reported for temperate
Table 9. Parametrs of the multiple regressions relating the effect of food concentration and temperature on
the duration of post-embryonic development of Daphnia gessneri.
Regression equation: 1/D. = a - b. 1/T + In F
D. = juvenile duration in hours; Τ = temperature in °C; F = food concentration in mgC.L" '
df = degress of fredom; F = variance ratio; Ρ = level of significance.
Daphnia gessneri
a b c df F Ρ
0 . 0 0 8 7 -0.611 0 . 0 0 0 5 4 2 , 5 5 2 4 . 3 7 0.001
Due to food concentration 3 6 . 3 9 0.001
and other tropical species (ROCHA, 1983; O R C U T T & P O R T E R , 1 9 8 4 , JAYATUNGA, 1986).
Numbers of juvenile instars
LEI & A R M I T A G E (1980) suggest that an increase in postembryonic develop ment time could arise either from an in crease in the duration of the individual ju venile instars without an increase in their number or from an increase in the number of juvenile instars without an increase in their individual duration or from an in crease in both duration and numbers of ju venile instars. From our evidence in Table 4, all three species lengthen their juvenile phase mainly by adding more juvenile in stars and that this occurs only at limiting food levels. However, at 32°C Daphnia gessneri is capable of increasing both du ration and numbers of juvenile instars as the food level is changed from 1.0 mgC/L t o 0 . 0 5 m g C / L .
The ratio D/D
J e
BONOU et al. (1991) and SAINT J E A N & B O N O U ( 1 9 9 4 ) s h o w very clearly the relationship between the num ber of juvenile instars and the size of the D / D ratio in Moina micrura from fish
j c
ponds in the Ivory C o a s t . Both papers show that in the 30°C experiments all in dividuals attained the primipara stage with 2 instars and a D / D ratio of 0.98 in 1991
.1 C
p a p e r and 0 . 8 9 in t h e 1994 p a p e r . Whereas there were 2instar, 3instar and 4instar primiparas present in the 26( )
C ex periments, with D / Dc ratios of 1.12 and
1.03, respectively. From our results in Table 6, these were wellfed Moina but there was probably some difference be
tween the food conditions in these two periods as well as a temperature difference. In general, the response to food limitation appears to involve delaying the primipara's development stage thus, inevitably, increas ing her age.
Body size effect
Within the three tropical cladocerans studied here, the smallest Moina reticulata
attained maturity in the fastest time at all food levels and under similar conditions compared with Diaphanosoma sarsi and
Daphnia gessneri, both larger animals. This confirms its classification by R O M A N O V S K Y (1985) and S A I N T JEAN & B O N O U (1994) as an opportu nistic ruderal species.
It provides some evidence to support the hypothesis that body size and duration of postembryonic development of cla docerans are positively related ( H A L L E T Α., 1976; ALLAN & GOULDEN, 1980).
ACKNOWLEDGMENTS
We wish to thank the National Re search Council of Brazil (CNPq) for fi nancial support to EH during her earlier postgraduate research and CAPES and the National Institute for Amazon Research (LNPA) for funding a leave of absence dur ing which this paper could be written at Royal Holloway College University of London using facilities for which we also give thanks.
Literature Cited
ALLAN, J.D.; GOULDEN, CE. 1980. Some as-pects of reproductive variation among
freshwater zooplankton. In: Evolution
and Ecology of Zooplankton
American Society of Limnology and Oceanography Special Symposium.
BONOU, C.A.; PAGANO, Μ.; SAINT-JEAN, L. 1991. Developpement et croissance
en poids de Moina (cf) micrura et de
Mesocyclops ogunnus dans un milieu saumatre tropical: les etangs de
piscicul-ture de Layo (Cotes-d'lvoire). Rev.
Hydrokiol. nop. 24(4):287-303. BOTTRELL. HI I. 1975. The relation between
temperature and duration of egg devel-opment in some epiphytic Cladocera and Copepoda from the River Thames, Read-ing, with a discussion of temperature
functions. Oecologia (Beri.),18: 63-84. BOTTRELL, H.H.;DUNCAN,A.; GLIWICZ,
Z.M.; GRYGIEREKJE.; HERZ1G, Α.; HILLBRICHT-ILKO W S K A , Α.; K U R A S A W A , H.; L A R S S O N , P.; W E G L E N S K A T . 1976. A review of some problems in zooplankton
produc-tion studies. Norwegian Journal
Zool-ogy, 24,419-456.
D E B E R N A R D I . R.; L A C Q U A , P.; SALDAVINI, E. 1978. Effects of tem-perature and food on development times
and growth in Daphnia obtusa Kurz. and
Simocephalus vetulus (O.F. Muller)
(Crustacea, Cladocera). Mem. 1st.
itai.ldrohiol. 36: 171-191.
DUNCAN, A. 1989. Food limitation and body size in the life cycles of planktonic
roti-fers and cladocerans. Hydrohiologia
186/187:11-28.
ESSLOVA.M. 1959. Embryonic development
of parthenogenetic eggs of Daphnia
pulex. Acta Soc.Zool.Bohem. 23.80-88. GRAS.R.; SAINT-JEAN,L. 1976. Duree du
d e v e l o p p e m e n t embryonnaire c h e z quelqucs especes de cladoccres et de c o p e p o d e s du lac Tchad.Cah. O . R . S . T . O . M . , s e r .
Hydrohiologia, X,4:233-254.
GRAS,R.; SAINT-JEAN,L. 1978. Duree et caracteristiques du developpment juve-nile dc quelqucs cladoccres du lac Tchad. Cah. O . R . S . T . O . M . . scr.
Hydrohiologia.X\l,2:119-136.
HALL, D.J. 1964. An experimental approach to the dynamics of a natural population of Daphnia goleata mendotae. Ecology.
45: 94-112.
HALL,D.J.; THRELKELD.S.T.; BURNS,C.W.; CROWLEYRH. 1976. The size efficiency hypothesis and the size structure of
zoop-lankton communities. Ann.Rev.Ecol.Syst.
7:177-208.
HERZIG,A. 1984. Temperature and life cycle
strategies of Diaphanosoma hrachiurum:
An experimental study on development,
growth and survival. Arc hi ν urn
Hydrohiologie, 101,(1/2) 143-178 H R B A C K O V A - E S S L O V A , M. 1962.
Postembryonic development of
cladocer-ans. I. Daphnia pulex group.
Vestn.Cesk.Spol.Zool. 26:212-233. HRBACKOVA-ESSLOVA, M. 1963. The
de-velopment of three species of Daphnia
in the surface water of the Slapy
Reservoir.Int. Revue gcs. Hydrobiol.
48:325-333.
INGLE.Z; WOOD,T; Β ANTA, A.M. 1937. A study of longevity, growth, reproduction,
and heart rate in Daphnia longispina as
influenced by limitation in quantity of
food. Journal of Experimental Zoology,
76:325-352.
JANA.B.B.; PAL.G.P. 1984. The life history
parameters of Diaphanosoma excisum
(Cladocera) grown in different cuhuring
media. Hydrohiologia, 118,205-212.
JANA.B.B.; PAL.G.P. 1985. The life history
parameters of Moina micrura (Kurz)
grown in different culturing media.
Wa-ter Res AS), 863-867.
JAYATUNGA.Y.N.A. 1986. The influence of
food and temperature on the life cycle characteristics of tropical cladoceran species from Kalawewa Reservoir, Sri Uinka. Ph.D Thesis. University of Lon-don. 41 Op.
KANKAALA.R; WULFF.F. 1981. experimental studies on temperature dependent embryonic and postembryonic developmental rates of
Bosmina longispina marítima (Cladocera) in
KORINEK, V. 1970. The embryonic and
post-embryonic development of Daphnia
hyalina from Lake Maggiore. Mem.ist Ital.ldrobiol. 26,85-95
LAMPERTW. 1875. A laboratory system for the cultivation of large numbers of Daphniae under controlled conditions.
Archiv Hydrobiologie, supplAS: 138-140.
LEI, C ; ARMITAGE.K.B. 1980. Growth, de-velopment and body size of field and
laboratory populations of Daphnia
ambigua. Oikos, 35,31-48.
LEVEQUE, C ; SAINT-JEAN.L. 1983. Sec-ondary production (zooplankton and
benthos). In: Lake Chad. (eds. J.P.
Carmouze, J.R. Durand and C.Leveque), pp 385-425. Dr. W. Junk Publishers, The Hague.
MACKERETH,F.J.H; HERON,J;
TALLING,JF. 1978. Water Analysis:
Some revised methods for Limnologists.
Freshwater Biological Association, 36,120pp.
MAGADZA,C.H.D. 1977. Determination of development period at various
tempera-tures in a tropical cladoccran Moina
dú-bia. De Guerne and Richard
Transac-tions Rhod. Scientist Association. 59,
6:41-45.
MUNRO.I.G., WHITE, R.W.G. 1975. Com-parison of the influence of temperature on the egg development and growth of
Daphnia longispina O.F. Muller (Crus-tacea: Cladocera) from two habitats in
Southern England. Oecologia (Beri.),
20,157-166.
MURUGAN.N. 1975. Egg production,
devel-opment and growth in Moina micrura
Kurz ( 1 8 7 4 ) (Cladocera:Moinidac).
Freshwat.Biol. 5:245-250
ORCUTT,J.D.; PORTER, K G . 1984. The syn-ergistic effects of temperature and food concentration on life history parameters of Daphnia parvula. Oecologia (Beri.), 63,300-306.
ROCHA.O. 1983. The influence of
food-tem-perature combinations on the duration of development, body size, growth and fecundity of Daphnia species. Ph. D
The-sis. University of London, 337pp. R O C H A A ; DUNCAN.A. 1985. The
relation-ship between cell carbon and cell volume
in freshwater algal species used in
zoop-lanktonic studies. Journal of Plankton
Research, "'(2):279-294.
ROMANOVSKY.Y.E. 1985. Food limitation and life history strategies in cladoceran
crustaceans. Arch.Hydrobiql. Beili.
Ergebn.Limnol.2\:363-312
S A I N T - J E A N . L . ; B O N O U . C . A . 1994. Growth, Production and Demography of
Moina micrura in Tropical Brackish Fish Ponds ( L a y o , Ivory Coast).
Hydrobiologia, 272,125-146.
VIJVERBERG.J. 1980. Effect of temperature in laboratory studies on development and growth of Cladocera and Copcpoda from Tjeukemeer, The Netherlands.
Fresh.Biol. 10:317-340.
WEGLENSKA.T 1971. The influence of vari-ous concentrations of natural food on the development, fecundity and production of planktonic crustacean filtrators.
Ekologie Polska, 19,427-473.