Effect of
In Vitro
Syncytium Formation on
the Severity of Human Metapneumovirus
Disease in a Murine Model
Laetitia Aerts1, Marie-Hélène Cavanagh1, Julia Dubois1, Julie Carbonneau1, Chantal Rhéaume1, Sophie Lavigne2, Christian Couture2, Marie-Ève Hamelin1, Guy Boivin1
*
1Centre de Recherche en Infectiologie of the Centre Hospitalier Universitaire de Québec and Université Laval, Quebec, Canada,2Anatomopathologie et cytologie, Institut Universitaire de Cardiologie et de Pneumologie de Québec and Université Laval, Quebec City, QC, Canada
Abstract
Human metapneumovirus (HMPV) is an important cause of acute respiratory tract infections (ARTI) in children, elderly individuals and immunocompromised patients.In vitro, different
HMPV strains can induce variable cytopathic effects ranging from large multinucleated syn-cytia to focal cell rounding. In this study, we investigated the impact of differentin vitro
phe-notypes of two HMPV strains on viral replication and disease severity in a BALB/c mouse model. We first generated two recombinant GFP-expressing HMPV viruses: C-85473, a syncytium-inducing strain (rC-85473) belonging to the A1 subtype and CAN98-75, a focal cell rounding-inducing strain (rCAN98-75) of the B2 subtype. We subsequently exchanged the F genes of both strains to create the chimeric viruses rC-85473_F and rCAN98-75_F. We demonstrated that the F protein was the sole protein responsible for the syncytium phe-notype and that viruses carrying a syncytium-inducing F protein replicated to significantly higher titersin vitro.In vivo, however, the virulence and replicative capacity of the different
HMPV strains did not appear to be solely dependent on the F gene but also on the viral background, with the strains containing the C-85473 background inducing more weight loss as well as increased lung viral titers, pro-inflammatory cytokines and inflammation than strains containing the CAN98-75 background. In conclusion, the F protein is the main deter-minant of syncytium formation and replication kineticsin vitro, although it is not the only
fac-tor implicated in HMPV disease severity in mice.
Introduction
Human metapneumovirus (HMPV) is a major cause of acute respiratory tract infections (ARTI) in children, elderly individuals and immunocompromised patients [1]. For instance, HMPV accounts for 10 to 15% of all hospitalizations for ARTI in children [2]. Clinical signs as-sociated with HMPV are similar to those asas-sociated with human respiratory syncytial virus
a11111
OPEN ACCESS
Citation:Aerts L, Cavanagh MH, Dubois J, Carbonneau J, Rhéaume C, Lavigne S, et al. (2015) Effect ofIn VitroSyncytium Formation on the Severity of Human Metapneumovirus Disease in a Murine Model. PLoS ONE 10(3): e0120283. doi:10.1371/ journal.pone.0120283
Academic Editor:Steven M. Varga, University of Iowa, UNITED STATES
Received:December 12, 2014
Accepted:February 5, 2015
Published:March 24, 2015
Copyright:© 2015 Aerts et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability Statement:HMPV Fusion gene sequences are available at GenBank accession numbers AY145289.1 (CAN98-75) and KM408076.1 (C-85473).
Funding:This work was supported by a grant from the Canadian Institutes of Health Research to GB (273261). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
(HRSV), ranging from mild respiratory problems to bronchiolitis and pneumonia [3]. Based on phylogenetic analysis of the F and G genes, HMPV strains can be classified into two main lineages (A and B), each containing 2 or 3 sub-lineages (A1, A2a, A2b, B1 and B2) [4,5]. Whether these HMPV lineages are associated with different clinical outcomes remains unclear; some groups found no evidence for differential severity between HMPV lineages [6,7], whereas others suggested more severe clinical disease associated with HMPV-A [8] or HMPV-B [9,10] strains. However, a recent study by our group, suggested that viral load rather than HMPV lineage was an independent risk factor for severe disease [11].
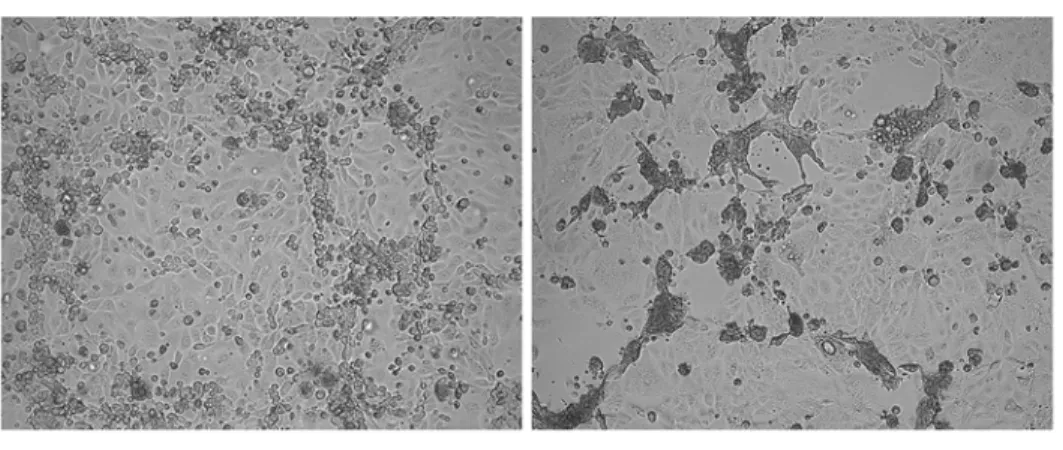
HMPV culture is notoriously difficult. Viruses have to be cultured several weeks before cy-topathic effects (CPE) occur and regular addition of exogenous trypsin is required. In addition, CPE differ greatly between strains ranging from typical HRSV-like syncytia to focal cell round-ing in tertiary monkey kidney cells such as LLC-MK2 cells (Fig. 1a).
HMPV is a member of thePneumovirinaesubfamily within theParamyxoviridaefamily [12]. Paramyxovirus entry into the host cells occurs through fusion of the cell membrane with the viral envelope. This fusion is mediated by viral surface glycoproteins. Membrane fusion of members of thePneumovirinaesubfamily (including HRSV and HMPV) is unique among paramyxoviruses, because the fusion (F) glycoprotein alone is sufficient for membrane fusion
Fig 1. Cytopathic effects of HMPV strains and recombinant HMPV viruses.(a) microscopic images of cytopathic effects induced by HMPV infection of LLC-MK2 monolayers. CAN98–75 (B2) induces focal
cell-rounding (left) whereas C-85473 (A1) induces multinucleated syncytia (right). Magnification = 10x. (b) Representation of the genomes of the 4 recombinant viruses used in this study; rC-85473 and rCAN98–75
represent the wild-type strains, rC-85473_F represents the chimeric rC-85473 strain in which the F gene has been replaced with that of CAN98–75 and rCAN98–75_F represent the chimeric rCAN98–75 in which the F
gene has been replaced with that of C-85473.
to occur without the requirement of an additional attachment glycoprotein [13,14]. The para-myxovirus F protein is synthesized as an inactive F0precursor protein that requires proteolytic cleavage into 2 disulfide-linked subunits (F1and F2) to be activated and capable of inducing membrane fusion. Proteolytic cleavage reveals a hydrophobic fusion peptide located at the N-terminus of the F1subunit, which is inserted into target cell membranes to initiate folding of two heptad repeats within the F1subunit, HRA and HRB, into an irreversible six-helix bundle. These conformational changes result in the formation of a fusion pore [15].
Although it is generally recognized that activated F protein-mediated cell-cell fusion is the cause of syncytium formation, the exact reasons why some strains induce large syncytia and oth-ers do not remain to be established. In addition, some HMPV strains are dependent on low-pH for membrane fusionin vitro[16]. Several studies have tried to elucidate which amino acids could be responsible for this pH-dependency. One group proposed that pH-dependency is de-pendent on the HMPV lineage [17]. Using 2 subtype A strains and 2 subtype B strains, they sug-gested that Gly294was responsible for pH-dependency of some subtype A viruses and that subtype A viruses carrying Glu294did not induce syncytium at any pH. On the other hand, both subtype B viruses with Glu at position 294 proved to induce syncytium in a pH-independent fashion. They proposed that a tetrad of variable amino acids at positions 294, 296, 396 and 404 was likely involved in protonation of conserved histidine residues at positions 368 and 435 in pH-dependent strains [18]. Chang et al, on the other hand, used the prototype pH-dependent strain CAN97–83 (A2) to propose that the tetrad at positions 294, 296, 396 and 438 was in-volved in destabilizing the histidine at position 435 [19]. However, due to the very limited num-ber of strains analyzed in these studies, it is very difficult to generalize these results. More importantly, the relevance of pH-dependency or—independency on severity of HMPV infection remains unexamined.
Moreover, within our collection of HMPV isolates, we have encountered type A strains that do induce syncytium at neutral pH and type B strains that do not. By comparing the type A1 strain C-85473 (syncytium phenotype) and the type B2 strain CAN98–75 (focal cell rounding phenotype) (Fig. 1a), we initially observed increased replication kineticsin vitroand increased virulence in BALB/c mice with the former strain. We then generated the recombinant viruses from these strains containing GFP as a reporter gene and we further swapped the F genes in both viruses (Fig. 1b). Herein, we report that replacing the F gene of a focal cell rounding-inducing strain with that of a syncytium-forming strain, or vice versa, is sufficient to alter the phenotype of the strainin vitro. We also investigated replication kinetics of all 4 recombinant strains in cell culture and in the lungs of infected BALB/c mice. We found that HMPV strains carrying the syncytium-inducing F protein replicated to higher titersin vitrothan non-syncy-tium F protein, but that the F protein was not the only contributing factor to HMPV disease se-verity in animals.
Materials and Methods
Cells and HMPV strains
LLC-MK2 cells (ATCC CCL-7) were maintained in minimal essential medium (MEM) (Life Technologies) supplemented with 10% fetal bovine serum (FBS) (Wisent).
supplemented with 0.0002% trypsin (Sigma). Virus stocks were concentrated on Amicon col-umns (Fisher Scientific) as previously described [20].
Virus quantification
Viral titers were determined by 10-fold serial dilutions of recombinant virus or lung homoge-nates in 24-well plates containing LLC-MK2 cells as previously reported [21]. Virus titers were reported as 50% tissue culture infectious doses (TCID50) per ml. TCID50values were calculated by the Reed and Muench method.
Alternatively, the number of PFU/ml was calculated to determine the MOI forin vitro infec-tion experiments (syncytium assay, real-time cell analysis, replicainfec-tion kinetics). Immunostain-ing of infected cells was performed with MAb 1017, a monoclonal antibody directed against the HMPV F protein (a gift from MedImmune), followed by peroxidase-labeled goat anti-hamster immunoglobulin (Cederlane) and TruBlue peroxidase substrate (KPL/Mandel) as pre-viously described [22].
Construction of antigenome- and supporting protein plasmids
A pSP72 plasmid (Promega) was used to generate the antigenome plasmids as previously re-ported [23]. Briefly, an NdeI to HpaI fragment was removed from plasmid pSP72 (Promega) and replaced by a T7 terminator, the hepatitis delta virus (HDV) ribozyme and a T7 promoter to yield pSP72-T7T-δ-T7P. cDNA was generated from viral RNA using the Superscript II re-verse transcriptase (Life technologies). PCR was carried out using PFU turbo polymerase (Life Technologies). The cDNA encoding the antigenome of C-85473 or CAN98–75 was assembled from 3 or 4 PCR fragments, cloned into temporary pJET plasmids (Thermo Scientific) and se-quenced before being cloned into the pSP72 plasmid. The GFP gene was flanked by the N gene start region and the F gene end region of the respective strains and inserted between the N gene and the antigenomic leader sequence using the restriction sites MluI and StuI for CAN98–75 and MluI and NheI for C-85473. Subsequently, the F genes were interchanged be-tween the 2 stains by site-directed mutagenesis using the Phusion DNA polymerase (New En-gland Biolabs) in the case of CAN98–75_F and using the commercial Gibson Assembly Cloning Kit (New England Biolabs) in the case of C-85473_F (S1 Fig.).
The N, P, L, and M2.1 ORFs of CAN98–75 were amplified by PCR using primers spanning the start and stop codons and flanked by XhoI and NotI restriction sites and were subsequently cloned in the multiple cloning site of the pTNT vector (Promega) to create supporting
protein plasmids.
All plasmids were sequenced using the ABI 3730 DNA analyzer and analyzed using BioEdit, version 7.2.0 prior to further use.
Recombinant virus rescue
and an aliquot was used to verify the sequence of the F gene (GenBank accession numbers: KM408076.1 and AY145289.1 for C-85473 and CAN98–75, respectively).
Syncytium quantification
Confluent monolayers of LLC-MK2 cells in black 24-well plates with flat and clear bottom (ibidi) were infected with each of the recombinant GFP-expressing HMPV viruses at an MOI of 0.01 in quadruplicate. Trypsine (0.0002%) was added every other day. Syncytium formation was evaluated on a daily basis using a fluorescent microscope. Three photographs were taken of each infected well at 20x magnification. In each field, 40 nuclei were counted and the num-ber of nuclei per cell was calculated.
Real time cell analysis
Real time cell analysis (RTCA) was performed using the xCELLigence System (ACEA). Fiftyμl of cell culture medium was added to each well of a 96-well E-Plate (ACEA) to obtain back-ground readings. LLC-MK2 cells were then added at 12 500 cells per well in 100μl of culture medium. The E-Plates were subsequently incubated for 30 min at room temperature and placed on the RTCA MP station (ACEA) located in an incubator (at 37°C and 5% CO2). The Cell Index (CI) values were measured automatically every 30 min. When CI reached a plateau (24 h after seeding), cells were washed 2 times with 200μl of PBS and 6 wells were infected with 150μl of each of the recombinant HMPV viruses at an MOI of 0.01. Infected E-plates were placed back into the RTCA MP station and the CI values were measured automatically every 30 min for 7 days. Trypsin (0.0002%) was added every other day. Cell indexes were nor-malized to mock-infected wells and the time until CI was reduced by 50% was calculated for each virus.
In vitro replication kinetics assay
Confluent monolayers of LLC-MK2 cells in 24-well plates were washed twice with PBS and in-fected with recombinant HMPV viruses at an MOI of 0.01. Trypsin (0.0002%) was added every other day. Three infected wells were harvested every 24 h for 7 days and supernatants were stored at -80°C. End-point titrations were performed on all samples to determine viral titers re-ported as TCID50/ml.
Animal studies
In vivo replication kinetics assay
Lungs were removed on days 3 through 6 pi, weighed and homogenized in 1 ml of PBS then centrifuged at 2000 rpm for 10 min. The supernatant was used to determine viral titers re-ported as TCID50/g of lung.
Cytokine analysis
An aliquot of 150μl of lung homogenates was added to 150μl of 50 mM KPO4, pH 6.0 buffer containing 0.2% CHAPS (Sigma) and 0.2% protease inhibitor cocktail (Sigma) and then stored at -20°C. On the day of the experiment, samples were centrifuged at 13,000 × g for 10 min at 4°C and then 50μl of the supernatant were used for cytokine quantification using a commercial multiplex mouse cytokine bead assay (Bio-Rad) according to the manufacturers’instructions. Experiments were performed in flat bottom 96-well plate and results were analyzed with the Luminex system (QIAGEN).
Histopathological analysis
Lungs were removed on day 5 pi, and fixed with 4% buffered formalin. Fixed lungs were subse-quently embedded in paraffin, sectioned in slices of 5μm, and stained with hematoxylin-eosin. Scoring of histologic parameters was performed by a medical biologist (SL) and an anatomic pathologist (CC), both with experience in pulmonary pathology, independently and blind to experimental data, on digitalized slides scanned at a resolution of 400X magnification (Nano-zoomer scanner and viewer, Hamamatsu, Japan). A semi-quantitative scale was used to score bronchial/endobronchial, peribronchial, perivascular, interstitial, pleural and intra alveolar in-flammation, capillary vascular congestion and pulmonary edema [21].
Statistical analysis
Repeated-Measure ANOVA’s with tukey post-hoc tests were used to analyze all the data except those involving RTCA for which two-tailed Student t-tests were used. All statistical analyses were performed using Prism 6.
Results
In vitro
and
in vivo
properties of two clinical HMPV strains
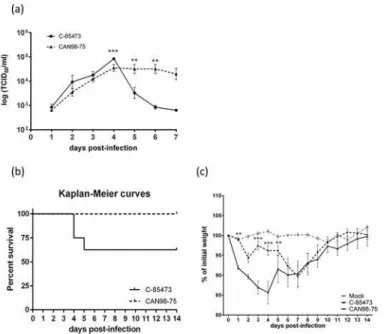
LLC-MK2 monolayers were infected with either C-85473 or CAN98–75 viruses at an MOI of 0.1 to evaluatein vitroreplication kinetics. The syncytium-inducing C-85473 strain replicated to a significantly higher titer than the focal cell rounding strain CAN98–75 on day 4 pi (8.6 ± 0.8 vs 4.2 ± 1.3 x104TCID50/ml; p<0.001) (Fig. 2a). After day 4, however, viral titers of
Generation of recombinant HMPV strains
Based on these observations, we postulated that the F protein, leading to syncytium formation, was also responsible for the increase in HMPV virulence. Thus, we generated recombinant vi-ruses for each strain and swapped the F proteins of both vivi-ruses in order to investigate the con-tribution of individual F proteins in each viral background. Full-length antigenome-plasmids were created for rC-85473, rCAN98–75, rC-85473_F and rCAN98–75 (S1 Fig.). Following co-transfection of the antigenome-plasmids and supporting plasmids into BSR T7/5 cells, GFP ex-pression was observed on average 48 h post-transfection. LLC-MK2 cells were then added to amplify viral production and recombinant viruses were rescued within 5 days of transfection.
In vitro
phenotype of four recombinant HMPV strains
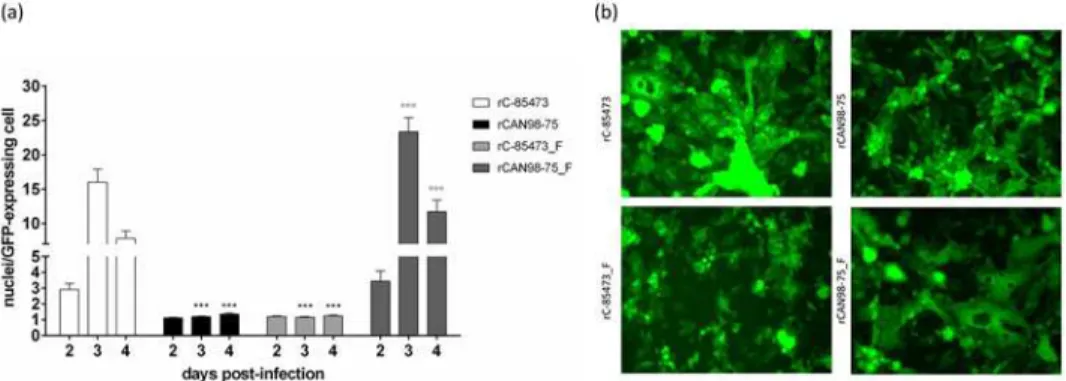
Thein vitrophenotype of each HMPV recombinant virus was investigated using fluorescence microscopy. The wild-type (WT) C-85473 strain is known to induce large syncytia and the re-combinant strain rC-85473 conserved the same phenotype with 15.9 ± 1.9 nuclei per GFP-ex-pressing cell on day 3 p.i (Fig. 3). On the other hand, the recombinant rCAN98–75 induced mostly focal cell rounding, similar to its WT counterpart with 1.2 ± 0.04 nuclei per GFP-ex-pressing cell on day 3 p.i. When the F gene of rC-85473 was replaced with that of rCAN98–75 (rC-85473_F), the phenotype was reverted and less syncytia were observed with 1.2 ± 0.05 nu-clei per GFP-expressing cell on day 3 p.i. Finally, rCAN98–75_F induced more syncytia with 23.3 ± 2.1 nuclei per GFP-expressing cell on day 3 p.i. These data confirmed that syncytium formation was primarily dependent on the F gene of the strain C-85473.
Fig 2. Replicative capacity of the clinical HMPV isolates C-85473 and CAN98–75in vitroand virulence
in BALB/c mice.(a) Replicative capacity of C-85473 and CAN98–75 strans in LLC-MK2 cells at an MOI of
0.1. (b) Kaplan-Meier survival curves of mice infected with 1x106TCID
50of C-85473 or CAN98–75. (c)
Weight loss curves of BALB/c mice infected with 1x106TCID
50of C-85473 or CAN98–75 or mock infected.
**, p<0.01;***, p<0.001 comparing C-85473 to CAN98–75 using Repeated Measures Two-way ANOVA.
Effects of four recombinant HMPV strains on cell state
We also investigated the effect of HMPV infection on the state of LLC-MK2 monolayers using RTCA. This method measures the change in electrical impedance across a cell monolayer in real-time. A parameter called cell index (CI) is used to quantify cell status based on the de-tected cell-electrode impedance; an elevated CI means that cells have fully adhered to the well and have proliferated, whereas a low CI indicates changes in morphology and viability of the cell monolayer. The changes in CI during an infection experiment using RTCA is shown in
Fig. 4aand mean time until the normalized CI was reduced by 50% is reported inFig. 4b. On average, it took rCAN98–75 28 h longer to reduce the CI by 50% than rC-85473 ((118 ± 9 h compared to 91 ± 8 h). Exchanging the F protein resulted in reverse phenotypes with 50% re-duction of CI obtained by 98 ± 4 h and 115 ± 2 h for rCAN98–75_F and rC-85473_F, respec-tively. These data show that the HMPV strains carrying the syncytium-inducing F protein from C-85473 alter the cell state of infected cells faster than viruses carrying the F protein from CAN98–75.
Fig 3. Syncytium formation induced by recombinant HMPV strains.(a) LLC-MK2 monolayers in 24 well-plates were infected with rHMPV at an MOI of 0.01 in quadruplicate. On days 2 through 4 pi, pictures were taken using fluorescent microscopy in 3 random fields (20x magnification) per well and the number of nuclei per GFP-expressing cell was calculated.***, p<0.001 comparing all other strains to rC-85473 and °°°, p<0.001 comparing all other strains to rCAN98–75 using Repeated Measures Two-way ANOVA. (b) An example of the observed difference in syncytium formation between the 4 recombinant strains on day 3 pi.
doi:10.1371/journal.pone.0120283.g003
Fig 4. Real-time cell analysis of recombinant HMPV strains.LLC-MK2 monolayers in 96 well-plates were infected with rHMPV at an MOI of 0.01 (a) Output of one real-time cell analysis (RTCA) experiment; data was normalized using mock-infected wells and normalized cell index is plotted. (b) Mean time until cell index is reduced by 50% from 4 independent experiments is plotted.*, p<0.05 using unpaired, two-tailed Student t-test.
In vitro
replication kinetics assay
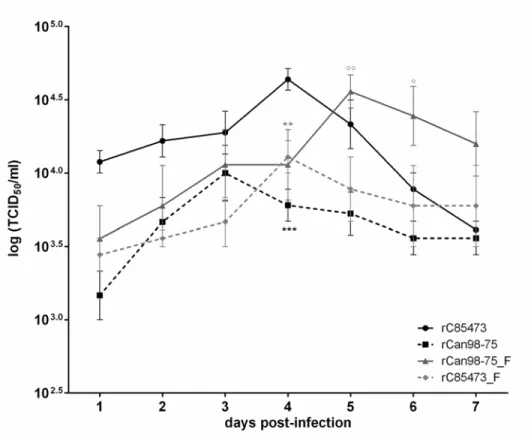
We next sought to investigate the replicative capacity of each recombinant virusin vitro, using an MOI of 0.01 (Fig. 5). Over a 7-day period, rC-85473 and rCAN98–75_F (maximum titers of 4.5 ± 0.7 and 3.8 ± 0.8 x104TCID50/ml, respectively) replicated to significantly higher titers (p<0.05) than rCAN98–75 and rC-85473_F (maximum titers of 1.2 ± 0.5 and 1.3 ± 0.4 x104
TCID50/ml, respectively). Of note, chimeric viruses (rCAN98–75_F and rC-85473_F) reached their peak titers 24 h later than recombinant WT viruses (rC-85473 and rCAN98–75). This ex-periment confirms that syncytium-inducing strains replicate to higher viral titers than non-syncytium inducing strains.
Replication kinetics of HMPV strains in lungs of BALB/c mice
To investigate the replicative capacity of all 4 recombinant virusesin vivo, BALB/c mice were infected with 6x105TCID50of recombinant HMPV strains. Backtitrations of the inoculum confirmed that the same amount of recombinant HMPV was given to all groups (6.3, 6.0, 6.8 and 6.4 x 105TCID50/mouse for rC-85473, rCAN98–75, rCAN98–75_F and rC-85473_F, re-spectively). Such inoculum did not induce mortality in any of the groups. Lungs of infected mice were harvested on day 3 through 6 post-infection to determine viral titers. All 4 recombi-nant viruses reached their peak of replication on day 4 pi (Fig. 6a). rC-85473 replicated to the Fig 5.In vitroreplicative capacity of recombinant HMPV strains.LLC-MK2 monolayers in 24 well-plates were infected with 4 rHMPV strains at an MOI of 0.01 in triplicate, one well per condition was harvested every 24 h for 7 days, frozen, sonicated and titrated on LLC-MK2 cells.***, p<0.001;**p<0.01 comparing all other strains to rC-85473 and °°, p<0.01; °, p<0.05 comparing all other strains to rCAN98–75 using Repeated Measures Two-way ANOVA.
highest titer (7.2 ± 2.1 x103TCID50/g lung) whereas rCAN98–75 had the lowest peak (4.6 ± 1.3 x102TCID50/g lung). Conversely, the chimeric strains rCAN98–75_F and rC-85473_F repli-cated to similar peak titers (3.2 ± 1.01 and 3.0 ± 0.9 x103TCID50/g lung, respectively).
Weight loss of HMPV-infected mice
Mice infected with 6x105TCID50of recombinant HMPV strains or mock infected were moni-tored for 14 days for clinical signs and weight loss. All infected mice lost between 3 and 7 per-cent of their initial weight between days 1 and 3, but only viruses with the C-85473 background (rC-85473 and rC85473_F) continued to lose weight on days 5–7, with statistically significant differences observed between rC-85473-viruses and rCAN98–75 viruses on days 6–9 (Fig. 6b). No significant difference in weight loss was observed between the 2 viruses with CAN98–75 background (rCAN98–75 and rCAN98–75_F). rC-85473-infected mice appeared to recuperate a little bit faster than mice infected with rC-85473_F with a statistically significant difference observed on day 9 pi only. Thus, the severity of the HMPV symptoms correlated better with the viral background than the F protein.
Pulmonary cytokine levels of HMPV-infected mice
On days 3 through 6 pi of the previously described experiment, an aliquot of lung homogenates was used to determine pulmonary cytokine/chemokine levels using a multiplexed bead assay (Fig. 7).
IL-2 peaked on day 5 pi for all 4 recombinant viruses and IL-2 levels were similar between groups at each time point. IL-6 also peaked on day 5 pi, but IL-6 levels were significantly higher in viruses with a C-85473 background compared to viruses with a CAN98–75 background on days 5 pi. IL-12 levels remained relatively stable between days 3 and 6 pi, except for rCAN98– 75 for which IL-12 levels were reduced by half by day 5 pi. Significantly higher levels of IL-12 were observed with the viruses harboring the C-85473 background compared to the rCAN98– 75 virus on all analyzed time-points. However, introducing the F protein of C-85473 into rCAN98–75 significantly increased IL-12 levels on days 5 and 6 pi. IFN-γlevels reached a pla-teau on days 5 and 6 pi for both viruses with a C-85473 background as well as for rCAN98– 75_F, whereas IFN-γlevels declined from day 3 onward for rCAN98–75. KC levels were
Fig 6. Lung viral titers and weight loss of BALB/c mice infected with recombinant HMPV strains. BALB/c mice were infected with 6x105TCID
50of rHMPV (as determined by backtitration). (a) On days 3 through 6, four mice per group were euthanized to determine pulmonary viral titers. (b) Six mice per group were monitored for weight loss on a daily basis for 14 days.***, p<0.001;*p<0.05 comparing all other strains to rC-85473.°°°, p<0.001; °, p<0.05 comparing all other strains to rCAN98–75 using Repeated Measures Two-way ANOVA.
highest on day 3 pi, with a second peak on day 5 pi for all viruses except rCAN98–75, for which KC levels increased a day later. KC levels were significantly higher for recombinant vi-ruses with a C-85473 background compared to rCAN98–75. Furthermore, the introduction of the F protein from C-85473 into the CAN98–75 background significantly increased KC levels on days 3, 5 and 6 pi compared to the wild-type rCAN98–75.
Fig 7. Pulmonary cytokine levels of BALB/c mice infected with recombinant HMPV strains.BALB/c mice were infected with 6x105TCID
50of rHMPV (as determined by back-titration). On days 3 through 6, four mice per group were euthanized to determine pro-inflammatory cytokine/chemokine levels in the lungs of infected mice. Mock-infected mice are representad as day 0.***, p<0.001;**, p<0.01;*p<0.05 comparing all other strains to rC-85473.°°°, p<0.001; °°, p<0.01; °, p<0.05 comparing all other strains to rCAN98–75 using Repeated Measures Two-way ANOVA.
MCP-1 levels were consistently significantly higher for viruses with a C-85473 background compared to rCAN98–75. Moreover, on day 5 pi, a significantly higher MCP-1 level was ob-served in mice infected with rCAN98–75_F compared to rCAN98–75. MIP-1αlevels peaked on day 5 pi for all strains except rCAN98–75, which peaked on or before day 3 pi. Furthermore, significantly higher MIP-1αlevels were observed on days 5 and 6 pi for 85473 and rC-85473_F as well as for rCAN98–75_F, compared to rCAN98–75. In general RANTES levels were lower for strain rCAN98–75 than for the other 3 viruses with statistically significant dif-ferences observed at early time-points.
Lung histopathology studies of HMPV-infected mice
On day 5 of the previously described experiment, lungs were harvested from 4 more mice per group to assess pulmonary inflammation. None of the groups showed signs of vascular conges-tion, pulmonary edema or bronchial inflammation. Moreover, none of the mice infected with rCAN98–75 showed signs of pulmonary inflammation for any of the analyzed parameters. Conversely, mice infected with rC-85473 or rC-85473_F showed mild, moderate or moderate to marked scores, with no significant differences in scores for any of the parameters between these two groups (total inflammation scores of 7.4 ± 0.7 and 7.9 ± 0.5 for 85473 and rC-85473_F, respectively) (Fig. 8). Importantly, the introduction of the syncytium-inducing F pro-tein into the rCAN98–75 background significantly increased histopathology scores for peri-bronchial, perivascular, interstitial and intra-alveolar inflammation (data not shown) as well as total inflammation (total inflammation score of 5.8 ± 0.6). In summary, although the C-85473 background induced significantly more pulmonary inflammation, the introduction of the syn-cytium-inducing F protein into the rCAN98–75 background significantly increased
lung inflammation.
Discussion
In the current study, we examined the effects of the F protein from two different HMPV strains, producing large syncytia in cell culture or not, onin vitroandin vivoreplication kinet-ics and virulence. We generated recombinant HMPV viruses representing either the
Fig 8. Total pulmonary inflammation scores of BALB/c mice infected with recombinant HMPV strains. BALB/c mice were infected with 6x105TCID
50of rHMPV (as determined by back-titration). On days 5, four mice per group were euthanized to determine histopathology scores of the lungs of infected mice.***, p<0.001; comparing all other strains to rC-85473.°°°, p<0.001 comparing all other strains to rCAN98–75 using Repeated Measures Two-way ANOVA.
syncytium-inducing phenotype (strain rC-85473) or the focal cell rounding phenotype (strain rCAN98–75) and we subsequently exchanged the F genes of both strains. We demonstrated that syncytium phenotype mainly depends on the F protein and that viruses carrying an F pro-tein that induces syncytium-formation replicate to higher titersin vitro. However, although the F protein appears to contribute to HMPV virulence, other genetic markers within the HMPV genome seem to impact on disease severity in mice.
HMPV is an important respiratory pathogen that can cause upper and lower RTIs. Virologi-cal risk factors for severe HMPV disease, such as HMPV subtype or lineage, have been the ob-ject of several investigations, with conflicting results [6,8–10,24]. We first sought to investigate whether thein vitrophenotype, namely syncytium formation, might be an indication of effi-cient HMPV replicative capacity. Previously, the syncytium-inducing strain NL/1/99 (subtype B1) was found to replicate to higher titers in Vero-118 cells than NL/1/00 (subtype A1), a strain that does not produce syncytium at neutral pH [23,25]. Similarly, we found that the syncy-tium-inducing strain C-85473 replicated to higher titers in LLC-MK2 cells than the focal cell rounding strain CAN98–75. However, to our knowledge, the impact ofin vitrophenotype on
in vivoreplication i.e., on lung titers, had not yet been examined. For this purpose, we infected BALB/c mice with either C-85473 or CAN98–75 clinical isolates and found that the former replicated to higher titers in the lungs on day 4 pi than CAN98–75 in 3 independent experi-ments (data from one representative experiment are shown inFig. 2). Furthermore, mortality was only observed in mice infected with strain C-85473.
Similarly to other paramyxoviruses, HMPV enters the host cell by fusion of viral and cellu-lar membranes, a step mediated by surface glycoproteins. On its surface, HMPV carries 3 gly-coproteins (F, G and SH) of which the F protein is the most conserved among HMPV strains [12]. As such, the HMPV F protein also shares structural features with other paramyxovirus F proteins; it is a class I viral fusion protein synthesized as inactive precursors (F0) that must be cleaved into 2 disulfide-linked F2-F1subunits to be fusion-competent [15]. Unlike members of theParamyxovirinaesubfamily, but similarly to other members of thePneumovirinae subfami-ly including HRSV, the HMPV F protein mediates membrane fusion in the absence of a sepa-rate viral attachment protein [16,26]. Furthermore, by transfecting cells with recombinant HMPV F proteins, it was demonstrated that the F protein alone is able to induce syncytium formation in cell culture [16,17].
To investigate the role of the F protein on viral replication bothin vitroandin vivo, we used reverse genetics to generate GFP-expressing recombinant viruses with either the C-85473 or the CAN98–75 genome and we further exchanged the F genes of both strains. Inserting the F gene of C-85473 into the CAN98–75 background restored the syncytium phenotype of strain C-85473, confirming that the F protein is responsible for syncytium formation independently of the genomic background.
Moreover, using RTCA, we demonstrated that the syncytium-inducing rHMPV strains in-duced changes in cell state 24 h before the focal cell rounding viruses. RTCA is a new electronic cell sensor array, in which the impedance, displayed as CI values, is continuously measured to evaluate cellular integrity [27,28]. This parameter includes cell proliferation, adhesion, viability, morphology and motility. This novel technique has been used to evaluate CPE and their inhibi-tion by neutralizing antibodies directed against flaviviruses [28] and influenza A viruses [27], but also to evaluate antiviral activity [29,30] and cytotoxicity [31]. Here, we demonstrate the usefulness of RTCA in investigating the dynamics of HMPV infection for the first time.
interfering RNAs that could alter the immune responsein vivo[32], but this led to lower viral stock titers (by about 1 log). Although the change in genomic background did not alter peak viral titers of viruses expressing the same F protein, both chimeric viruses reached their peak of replication a day later than WT rHMPV strains; this could indicate some influence of the geno-mic background on F protein incorporation and/or expression.
Notably, we investigated whetherin vitrophenotypes also correlated within vivoreplication titers. We observed that peak lung titers occurred on day 4 pi for all recombinant HMPV virus-es. Again, rC-85473 was associated with the highest viral titers and introducing the F protein of CAN98–75 into this strain significantly reduced peak viral titers (Fig. 6a). Conversely, the rCAN98–75 strain generated the lowest peak viral titers and introducing the F protein of C-85473 significantly increased maximum pulmonary viral titers. However, the overlap in the replication curves of the two chimeric viruses suggests thatin vivoproperties were not exclu-sively dependent on the F protein. This was even more evident by looking at weight loss curves where strains were primarily segregated by their genomic background and not by their F pro-tein. Similarly, pro-inflammatory cytokine/chemokine levels and pulmonary inflammation scores were increased in the rC-85473 strains independently of the F protein, although rCAN98–75_F induced higher cytokine levels (particularly MIP-1α, MCP-1, IL-12 and KC) and significantly more pulmonary inflammation than the prototypic rCAN98–75 strain, sug-gesting that the F protein does have an effect on HMPV pathogenesis as well.
We investigated 8 cytokines/chemokines (IL-2, IL-6, IL-12, IFN-γ, KC, MIP-1α, MCP-1 and RANTES) that had previously been described to be up-regulated in the lungs of HMPV-in-fected mice [21,33,34]. Although different inoculums and cytokine detection methods were used in previously-published studies, we also observed up-regulation of all cytokines/chemo-kines in mice infected with C-85473 strains, but only MIP-1αand MCP-1, two chemokines in-volved in the recruitment and activation of leukocytes, as well as IL-12, a cytokine inin-volved in Th1 differentiation, were significantly up-regulated for rCAN98–75 compared to mock-infected mice (represented as day 0 on the graph).
Unlike HRSV, HMPV does not encode non-structural proteins known to inhibit antiviral immune responses. Therefore, other viral proteins must be involved in immune evasion mech-anisms. Among HMPV-expressed proteins, the SH and G glycoproteins, the P phosphoprotein and the M2–2 protein have been found to have immune evasive properties. The presence of the SH protein reduced the expression of TNF-α, IL-6, KC and MCP-1 in mice infected with WT HMPV compared to mice infected with HMPV lacking the SH protein, in an NF-κ B-dependent manner [35]. The G protein was found to reduce cytokine/chemokine levels in cell cultures by inhibiting RIG-1 signaling [36,37]. Furthermore, the G protein inhibited
TLR4-signaling in dendritic cells [38]. Goutagny et al. observed that a HMPV-B1 strain im-paired type I IFN production, specifically by prevented RIG-I-mediated sensing of HMPV viral RNA, in a P protein-dependent manner [39]. Finally, the M2–2 protein inhibited MAVS-in-duced IFN-βgene transcriptionin vitro[40]. One or several of these viral proteins may be im-plicated in the observed differences in pathogenesis between strains with the C-85473 and CAN98–75 genomic background and this will be further investigated.
same tetrad as NL/1/94, a pH-independent syncytium-inducing strain [18]. This suggests that residues at other positions are likely to influence syncytium formation at neutral pH. Given the importance of the heptad repeats located in F1subdomain of the F protein for viral fusion, it would be interesting to examine the effect of the five amino acid changes in HRA and the two amino acid changes in HRB that differentiate our two prototype strains, on syncytium forma-tion at neutral pH.
In conclusion, we have demonstrated the importance of the HMPV F protein for syncytium formation andin vitroreplication and further showed that the F protein contributes to some extent, but not exclusively, to the virulence potential of different strains in mice. Therefore, it is unlikely that thein vitrophenotype of an HMPV strain alone is sufficient to predict the severity of HMPV disease. Other syncytium-inducing and non-syncytium inducing HMPV strains of different subtypes should be studied to confirm our results. Furthermore the effects of other viral genes on HMPV pathogenesis should be investigated using recombinant viruses in differ-ent animal models.
Supporting Information
S1 Fig. Construction of HMPV antigenome plasmids.Schematic representation of the clon-ing steps for rCAN98–75 (a) and for rC-85473 (b). Three or four cDNA fragments (A-C for rC-85473 and A-D for rCAN98–75) were cloned into the pSP72-T7T-δ-T7Pvector. The ob-tained antigenome plasmids were then used to swap the F genes. (c) Schematic representation of the cloning steps used to obtain rCAN98–75_F; a fragment covering M to SH was amplified from the rCAN98–75 antigenomic plasmid and cloned into the temporary pJET vector. Simul-taneously, the F gene was amplified from C-85473 cDNA and also cloned into pJET. Both plas-mids were digested and the F gene was ligated into the temporary pJET plasmid. From this vector, a fragment covering the region M to M2 was amplified and used as primers for site-di-rected mutagenesis of the rCAN98–75 antigenomic plasmid. (d) A schematic representation of the cloning steps used to obtain 85473_F; a fragment containing the region P to M2 of rC-85473 was cloned into a temporary pJET vector. A fragment of the F gene of rCAN98–75, flanked by the restriction sites EcoRV and PacI, was amplified by PCR and subsequently cloned into the temporary pJET vector containing the rC-85473 fragment. From this vector, fragment 1 was amplified by PCR. Fragment 2 was amplified directly from the plasmid containing the rCAN98–75 antigenomic vector and fragments 3 and 4 were amplified from the rC-85473 anti-genomic vector. Finally the pSP72 vector was digested to obtain fragment 5. All 5 fragments were then ligated using the Gibson Assembly cloning kit.
(TIF)
Acknowledgments
We would like to thank Jean-Christophe Le Bayon for his help in setting-up the reverse genet-ics system for HMPV.
Author Contributions
References
1. Kahn JS. Epidemiology of human metapneumovirus. Clin Microbiol Rev. 2006; 19(3):546–57. doi:10.
1128/CMR.00014-06PMID:16847085
2. Feuillet F, Lina B, Rosa-Calatrava M, Boivin G. Ten years of human metapneumovirus research. J Clin Virol. 2012; 53(2):97–105. doi:10.1016/j.jcv.2011.10.002PMID:22074934
3. Papenburg J, Boivin G. The distinguishing features of human metapneumovirus and respiratory syncy-tial virus. Rev Med Virol. 2010; 20(4):245–60. doi:10.1002/rmv.651PMID:20586081
4. Papenburg J, Carbonneau J, Isabel S, Bergeron MG, Williams JV, De Serres G, et al. Genetic diversity and molecular evolution of the major human metapneumovirus surface glycoproteins over a decade. J Clin Virol. 2013; 58(3):541–7. doi:10.1016/j.jcv.2013.08.029PMID:24041471
5. Huck B, Scharf G, Neumann-Haefelin D, Puppe W, Weigl J, Falcone V. Novel human metapneumo-virus sublineage. Emerg Infect Dis. 2006; 12(1):147–50. Epub 2006/02/24. doi:10.3201/eid1201.
050772PMID:16494734
6. Agapov E, Sumino KC, Gaudreault-Keener M, Storch GA, Holtzman MJ. Genetic variability of human metapneumovirus infection: evidence of a shift in viral genotype without a change in illness. J Infect Dis. 2006; 193(3):396–403. doi:10.1086/499310PMID:16388487
7. Xiao NG, Xie ZP, Zhang B, Yuan XH, Song JR, Gao HC, et al. Prevalence and clinical and molecular characterization of human metapneumovirus in children with acute respiratory infection in China. Pediatr Infect Dis J. 2010; 29(2):131–4. Epub 2010/02/06. PMID:20135829
8. Vicente D, Montes M, Cilla G, Perez-Yarza EG, Perez-Trallero E. Differences in clinical severity be-tween genotype A and genotype B human metapneumovirus infection in children. Clin Infect Dis. 2006; 42(12):e111–3. doi:10.1086/504378PMID:16705567
9. Papenburg J, Hamelin ME, Ouhoummane N, Carbonneau J, Ouakki M, Raymond F, et al. Comparison of risk factors for human metapneumovirus and respiratory syncytial virus disease severity in young children. J Infect Dis. 2012; 206(2):178–89. Epub 2012/05/04. doi:10.1093/infdis/jis333PMID:
22551815
10. Pitoiset C, Darniot M, Huet F, Aho SL, Pothier P, Manoha C. Human metapneumovirus genotypes and severity of disease in young children (n = 100) during a 7-year study in Dijon hospital, France. Journal of medical virology. 2010; 82(10):1782–9. doi:10.1002/jmv.21884PMID:20827777
11. Roussy J-F, Carbonneau J, Ouakki M, Papenburg J, Hamelin M-È, De Serres G, et al. Human metap-neumovirus viral load is an important risk factor for disease severity in young children. J Clin Virol. 2014; 60(2):133–40. doi:10.1016/j.jcv.2014.03.001PMID:24686044
12. van den Hoogen BG, Herfst S, Sprong L, Cane PA, Forleo-Neto E, de Swart RL, et al. Antigenic and ge-netic variability of human metapneumoviruses. Emerg Infect Dis. 2004; 10(4):658–66. doi:10.3201/
eid1004.030393PMID:15200856
13. Cox RG, Williams JV. Breaking in: human metapneumovirus fusion and entry. Viruses. 2013; 5(1): 192–210. doi:10.3390/v5010192PMID:23325326
14. Chang A, Dutch RE. Paramyxovirus fusion and entry: multiple paths to a common end. Viruses. 2012; 4(4):613–36. Epub 2012/05/17. doi:10.3390/v4040613PMID:22590688
15. Lamb RA, Jardetzky TS. Structural basis of viral invasion: lessons from paramyxovirus F. Curr Opin Struct Biol. 2007; 17(4):427–36. doi:10.1016/j.sbi.2007.08.016PMID:17870467
16. Schowalter RM, Smith SE, Dutch RE. Characterization of human metapneumovirus F protein-promoted membrane fusion: critical roles for proteolytic processing and low pH. J Virol. 2006; 80(22):10931–41.
Epub 2006/09/15. doi:10.1128/JVI.01287-06PMID:16971452
17. Herfst S, Mas V, Ver LS, Wierda RJ, Osterhaus AD, Fouchier RA, et al. Low-pH-induced membrane fu-sion mediated by human metapneumovirus F protein is a rare, strain-dependent phenomenon. J Virol. 2008; 82(17):8891–5. Epub 2008/07/04. doi:10.1128/JVI.00472-08PMID:18596097
18. Mas V, Herfst S, Osterhaus AD, Fouchier RA, Melero JA. Residues of the human metapneumovirus fu-sion (F) protein critical for its strain-related fufu-sion phenotype: implications for the virus replication cycle. J Virol. 2011; 85(23):12650–61. Epub 2011/09/23. doi:10.1128/JVI.05485-11PMID:21937649
19. Chang A, Hackett BA, Winter CC, Buchholz UJ, Dutch RE. Potential electrostatic interactions in multi-ple regions affect human metapneumovirus F-mediated membrane fusion. J Virol. 2012; 86(18): 9843–53. doi:10.1128/JVI.00639-12PMID:22761366
21. Hamelin ME, Yim K, Kuhn KH, Cragin RP, Boukhvalova M, Blanco JC, et al. Pathogenesis of human metapneumovirus lung infection in BALB/c mice and cotton rats. J Virol. 2005; 79(14):8894–903. doi:
10.1128/JVI.79.14.8894-8903.2005PMID:15994783
22. Deffrasnes C, Hamelin ME, Prince GA, Boivin G. Identification and evaluation of a highly effective fu-sion inhibitor for human metapneumovirus. Antimicrob Agents Chemother. 2008; 52(1):279–87. doi:
10.1128/AAC.00793-07PMID:17967906
23. Herfst S, de Graaf M, Schickli JH, Tang RS, Kaur J, Yang CF, et al. Recovery of human metapneumo-virus genetic lineages A and B from cloned cDNA. J Virol. 2004; 78(15):8264–70. doi:10.1128/JVI.78.
15.8264-8270.2004PMID:15254198
24. Sloots TP, Mackay IM, Bialasiewicz S, Jacob KC, McQueen E, Harnett GB, et al. Human metapneumo-virus, Australia, 2001–2004. Emerg Infect Dis. 2006; 12(8):1263–6. doi:10.3201/eid1208.051239
PMID:16965711
25. de Graaf M, Herfst S, Schrauwen EJA, van den Hoogen BG, Osterhaus ADME, Fouchier RAM. An im-proved plaque reduction virus neutralization assay for human metapneumovirus. Journal of virological methods. 2007; 143(2):169–74. PMID:17420056
26. Karron RA, Buonagurio DA, Georgiu AF, Whitehead SS, Adamus JE, Clements-Mann ML, et al. Respi-ratory syncytial virus (RSV) SH and G proteins are not essential for viral replication in vitro: clinical eval-uation and molecular characterization of a cold-passaged, attenuated RSV subgroup B mutant. Proc Natl Acad Sci U S A. 1997; 94(25):13961–6. Epub 1998/02/12. PMID:9391135
27. Tian D, Zhang W, He J, Liu Y, Song Z, Zhou Z, et al. Novel, real-time cell analysis for measuring viral cytopathogenesis and the efficacy of neutralizing antibodies to the 2009 Influenza A (H1N1) virus. PLoS ONE. 2012; 7(2):e31965. doi:10.1371/journal.pone.0031965PMID:22363775
28. Fang Y, Ye P, Wang X, Xu X, Reisen W. Real-time monitoring of flavivirus induced cytopathogenesis using cell electric impedance technology. Journal of virological methods. 2011; 173(2):251–8. doi:10.
1016/j.jviromet.2011.02.013PMID:21349291
29. Sharma BN, Li R, Bernhoff E, Gutteberg TJ, Rinaldo CH. Fluoroquinolones inhibit human polyomavirus BK (BKV) replication in primary human kidney cells. Antiviral Res. 2011; 92(1):115–23. Epub 2011/07/
30. doi:10.1016/j.antiviral.2011.07.012PMID:21798289
30. Sharma BN, Marschall M, Henriksen S, Rinaldo CH. Antiviral effects of artesunate on polyomavirus BK replication in primary human kidney cells. Antimicrob Agents Chemother. 2014; 58(1):279–89. Epub
2013/10/23. doi:10.1128/aac.01800-13PMID:24145549
31. Fonteh PN, Keter FK, Meyer D. New bis(thiosemicarbazonate) gold(III) complexes inhibit HIV replica-tion at cytostatic concentrareplica-tions: Potential for incorporareplica-tion into virostatic cocktails. J Inorg Biochem. 2011; 105(9):1173–80. doi:10.1016/j.jinorgbio.2011.05.011PMID:21708102
32. van den Hoogen BG, van Boheemen S, de Rijck J, van Nieuwkoop S, Smith DJ, Laksono B, et al. Ex-cessive production and extreme editing of human metapneumovirus defective interfering RNA is asso-ciated with type I IFN induction. J Gen Virol. 2014; 95(8):1625–33. doi:10.1099/vir.0.066100-0PMID:
24760760
33. Herd KA, Nelson M, Mahalingam S, Tindle RW. Pulmonary infection of mice with human metapneumo-virus induces local cytotoxic T-cell and immunoregulatory cytokine responses similar to those seen with human respiratory syncytial virus. J Gen Virol. 2010; 91(5):1302–10. Epub 2010/01/08. doi:10.
1099/vir.0.015396-0PMID:20053825
34. Huck B, Neumann-Haefelin D, Schmitt-Graeff A, Weckmann M, Mattes J, Ehl S, et al. Human metap-neumovirus induces more severe disease and stronger innate immune response in BALB/c mice as compared with respiratory syncytial virus. Respir Res. 2007; 8:6. Epub 2007/01/30. doi: 10.1186/1465-9921-8-6PMID:17257445
35. Bao X, Kolli D, Liu T, Shan Y, Garofalo RP, Casola A. Human metapneumovirus small hydrophobic pro-tein inhibits NF-kappaB transcriptional activity. J Virol. 2008; 82(16):8224–9. doi:
10.1128/JVI.02584-07PMID:18550666
36. Bao X, Kolli D, Ren J, Liu T, Garofalo RP, Casola A. Human metapneumovirus glycoprotein G disrupts mitochondrial signaling in airway epithelial cells. PLoS One. 2013; 8(4):e62568. doi:10.1371/journal. pone.0062568PMID:23626834
37. Bao X, Liu T, Shan Y, Li K, Garofalo RP, Casola A. Human metapneumovirus glycoprotein G inhibits in-nate immune responses. PLoS Pathog. 2008; 4(5):e1000077. doi:10.1371/journal.ppat.1000077 PMID:18516301
38. Kolli D, Bao X, Liu T, Hong C, Wang T, Garofalo RP, et al. Human metapneumovirus glycoprotein G in-hibits TLR4-dependent signaling in monocyte-derived dendritic cells. J Immunol. 2011; 187(1):47–54.
doi:10.4049/jimmunol.1002589PMID:21632720
interference of RIG-I ligand recognition by HMPV-B1 phosphoprotein. J Immunol. 2010; 184(3): 1168–79. doi:10.4049/jimmunol.0902750PMID:20042593
40. Ren J, Wang Q, Kolli D, Prusak DJ, Tseng CT, Chen ZJ, et al. Human metapneumovirus M2–2 protein
inhibits innate cellular signaling by targeting MAVS. J Virol. 2012; 86(23):13049–61. doi:10.1128/JVI.