Phylogenetic incongruence inferred with two mitochondrial genes in Mepraia spp. and Triatoma eratyrusiformis (Hemiptera, Reduviidae)
Texto
Imagem
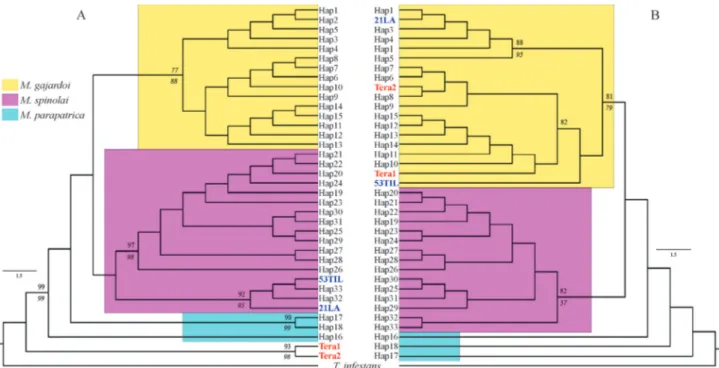
Documentos relacionados
Therefore, is possible to conclude that the species Chrysoperla externa from Jaboticabal, SP, to this genetic marker – COI – forms a single population and that in the use of
molecular marker for populations, species, and phy- logenetic relationships in Triatominae (Hemiptera: Reduviidae), vectors of Chagas disease. Nuclear rDNA
A presente dissertação tentará fazer uma análise sobre a situação atual de alunos internacionais da ULED de pós-licenciatura, a fim de acompanhar o desenvolvimento atual do ensino
O diabetes melittus é uma doença que pode afetar o ambiente bucal ao predispor o paciente a uma série de alterações, como infecções fúngicas, doença
Contudo, quando entram em cena os conjuntos de documentos acumulados por indivíduos, todos os princípios consagrados parecem ser colocados em suspeição, descortinando um
Since Swales’ original contribution, genre has made a signifi cant contribution to EAP, both in research and pedagogy, and especially in the teaching of writing to ESL
Os objetivos específicos foram: - desenvolver modelos matemáticos para simular sistemas de controle da força de contato entre o mecanismo de corte em plataforma de colhedoras e
Assim, neste caderno, apresentamos três unidades, envolvendo cinco línguas: na pri- meira, encontramos imagens e textos com os quais nos deparamos casualmente, sensibi- lizando para
