APLICAÇÕES DAS ENZIMAS LIGNINOLÍTICAS DE
Pleurotus pulmonarius
NA ELIMINAÇÃO DE
POLUENTES AMBIENTAIS
DANIELA FARANI DE SOUZA
Maringá 2008
DANIELA FARANI DE SOUZA
APLICAÇÕES DAS ENZIMAS LIGNINOLÍTICAS DE
Pleurotus pulmonarius
NA ELIMINAÇÃO DE
POLUENTES AMBIENTAIS
Tese apresentada ao programa de Pós Graduação em Ciências Biológicas da Universidade Estadual de Maringá, como parte dos requisitos para obtenção do título de Doutor em Ciências Biológicas.
Orientadora
APRESENTAÇÃO
Esta tese de doutoramento está apresentada na forma de dois artigos científicos
REMOVAL OF PENTACHLOROPHENOL BY Pleurotus pulmonarius IN
SUBMERGED CULTURES UNDER LIGNINOLYTIC AND NON-LIGNINOLYTIC CONDITIONS. Daniela Farani de Souza, Cristina Giatti Marques de Souza, Maria Aparecida Ferreira Costa, Cinthia Gandolfi Bôer, Márcia Fernandes Nishiyama, Heloísa Bressan Gonçalves, and Rosane Marina Peralta, a ser submetido ao periódico científico CHEMOSPHERE.
THE RATIO Mn PEROXIDASE/LACCASE IS HIGHLY AFFECTED BY THE INITIAL
MOISTURE CONTENT ON CORN COB SOLID STATE CULTURES OF Pleurotus
pulmonarius. Daniela Farani De Souza, Cissa Kelmer Bracht, Ana Maria
RESUMO
Os elementos estruturais principais da madeira são celulose, hemicelulose e lignina. Lignina é um polímero aromático impermeável a água, encontrado em todas as plantas superiores. A madeira e outros tecidos vasculares geralmente contém 20 a 30% de lignina. Este polímero é sintetizado pela polimerização oxidativa de três álcoois cinamílicos substituídos (precursores fenólicos) iniciada pelas lacases e/ou peroxidases. Estas reações inespecíficas criam um biopolímero tridimensional, heterogêneo e com alta massa molecular. Como uma macromolécula aromática complexa, a lignina fornece resistência e rigidez às paredes celulares e tecidos de todas as plantas vasculares por estar em estreita associação com a celulose e as hemiceluloses. Além disso, a lignina está envolvida no transporte de água nos vegetais, além de formar uma barreira contra a destruição microbiana por dificultar o acesso aos polissacarídeos rapidamente assimiláveis.
Devido à sua heterogeneidade e tipos de ligações que contém, a lignina não pode ser clivada por enzimas hidrolíticas como a maioria dos outros polímeros naturais (celulose, amido, proteínas, etc). A lignina é, entretanto, degradada pelos fungos causadores das podridões branca, marrom e macia da madeira. Os fungos causadores da podridão branca da madeira são capazes de degradar tanto os carboidratos quanto a lignina, enquanto os causadores das podridões marrom e macia utilizam preferencialmente a celulose e as hemiceluloses. Fungos causadores da podridão branca são os únicos organismos capazes de degradar a lignina a CO2 e água. Entretanto, eles não conseguem utilizar este substrato como única fonte de carbono e energia.
Manganês peroxidase (MnP, EC 1.11.1.13) é uma heme-proteína glicosilada extracelular. Esta enzima é expressa em múltiplas formas com massas moleculares variando entre 40 e 48 kDa e pI entre 2,9 e 7,0. A enzima tem sido descrita em muitos basidiomicetos causadores da podridão branca. A principal função da MnP é a oxidação do Mn+2 a Mn+3. MnP utiliza Mn+2 como substrato preferencial (doador de electrons). O íon Mn+3 gerado pela mnP é um oxidante potente, porém instável em meio aquoso. Mn+3 é estabilizado pela formação de complexos com ácidos orgânicos fisiológicos, oxalatos e malonatos, por exemplo, e difunde-se para longe do sítio ativo. Estes quelatos Mn+3-ácidos orgânicos são oxidantes potentes capazes de oxidar grande variedade de substratos incluindo compostos fenólicos e corantes.
As lacases (benzenodiol:oxigênio oxido-redutases, EC 1.10.3.2) são enzimas multi-cobre pertencentes ao grupo das oxidases azuis, que catalisam a oxidação de ampla variedade de compostos aromáticos (particularmente fenóis) com a redução do oxigênio a água. Numa reação típica, um substrato fenólico perde um elétron dando origem a um radical ariloxil. Esta espécie ativa pode ser convertida a uma quinona num segundo estágio da oxidação.
Fungos causadores da podridão branca e suas enzimas ligninolíticas, possuidoras de ampla especificidade pelo substrato, tem grande aplicação potencial nos processos de biopolpação e biobranqueamento, bem como na bioremediação de poluentes aromáticos com similaridades estruturais a lignina. O uso de fungos que degradam a lignina ou de suas enzimas ligninolíticas isoladas no tratamento da polpa do papel poderia fornecer maior seletividade na remoção da lignina comparada aos processos químicos.
Pleurotus sp pertence a sub classe dos fungos causadores da podridão
branca produtores de Mn peroxidases e lacases mas não lignina peroxidases. Uma das espécies mais importantes é Pleurotus pulmonarius (FR) Quélet, que foi utilizada
neste trabalho.
No primeiro artigo foi avaliada a habilidade do P. pulmonarius remover
pentaclorofenol (PCF) em culturas submersas ligninolíticas e não ligninolíticas. Para se obter uma condição ligninolítica, compostos fenólicos solúveis de sabugo de milho foram utilizados como indutores das enzimas ligninolíticas. Os compostos fenólicos aumentaram a produção de lacase de 32,1±3,0 U/L no meio basal para 532 U/L, enquanto nenhuma alteração na produção da Mn peroxidase foi observada (18,0 U/L). Cerca de 70% da concentração inicial de PCF (25 ppm) foi removida nas culturas submersas sob condições ligninolíticas, enquanto não mais do que 20% do PCF foi removido nas culturas sob condições não ligninolíticas. A produção de lacase parece ser fundamental para a remoção do PCF por P. pulmonarius.
No segundo artigo, a produção de lacase e Mn peroxidase por P. pulmonarius
e a sua capacidade em descolorir os corantes industriais Remazol Brilliant Blue R (RBBR), vermelho do congo, azul de metileno e violeta de etila foram estudados em culturas em estado sólido. A condição de alta umidade inicial (80-90%) promoveu fortemente a expressão da lacase, enquanto a Mn peroxidase foi produzida principalmente nas culturas desenvolvidas em baixas umidades iniciais (50-70%). Culturas contendo altas atividades de Mn peroxidase foram mais eficientes na descoloração dos corantes vermelho do congo, violeta de etila e azul de metileno. Os filtrados livres de células com alta atividade Mn peroxidase foram mais eficientes em descolorir vermelho do congo, violeta de etila e azul de metileno, enquanto RBBR foi igualmente descolorido por extratos com alta atividade Mn peroxidase e por extratos com alta atividade de lacase. Nenhum extrato conseguiu descolorir Poly R478. Os dados obtidos sugerem que a Mn peroxidase de P. pulmonarius é mais
eficiente que sua lacase na descoloração dos corantes testados.
Os resultados obtidos neste estudo mostram claramente o potencial de
Pleurotus pulmonarius na remoção de compostos aromáticos e no tratamento de
efluentes contaminados com corantes.
Palavras-chaves: Pleurotus pulmonarius, descoloração de corantes,
SUMMARY
The major structural elements of wood are cellulose, hemicellulose and lignin. Lignin is a water-impermeable aromatic polymer found in all higher plants. Wood and other vascular tissues generally are 20-30% lignin. It is synthesized by oxidative polymerization of the precursors p-coumaryl alcohol, coniferyl alcohol and sinapyl alcohol, initiated by laccase and/or peroxidase. These unspecific reactions create a high molecular weight, heterogeneous and three-dimensional biopolymer. As a complex aromatic macromolecule, it provides strength and rigidity to cell walls and tissues of all vascular plants by acting as a glue between the polysaccharide filaments and fibers. In addition, lignin is involved in water transport in plants and forms a barrier against microbial destruction by protecting the readily assimilable polysaccharides.
Because of the bond types and their heterogeneity, lignin cannot be cleaved by hydrolytic enzymes as most other natural polymers (cellulose, starch, proteins, etc). Lignin is decayed by white-rot, brown-rot and soft-rot fungi. White-rot fungi are able to degrade both carbohydrates and lignin, whereas brown-rot and soft-rot fungi prefer cellulose and hemicelluloses as substrates. Fungi causing white-rot are so far the only organisms known that degrade lignin to CO2 and H2O, but they cannot use this substrate as sole carbon and energy source.
Lignin biodegradation is an oxidative process, involving enzymes such as lignin peroxidase (LiP), manganese peroxidase (MnP), versatile peroxidase (VP) and laccase (lcc). The peroxidases and laccases have the ability to catalyze one-electron oxidations resulting in the formation of radicals which undergo numerous spontaneous reactions. These, in turn, lead to various bond cleavages including aromatic ring fission. In recent years, more emphasis has been placed on analyzing specially Mn peroxidases and laccases, because these enzymes are produced by most white-rot fungi, including species whick lack LiP.
unstable in aqueous media. Mn3+ is stabilized by forming complexes with physiologically occurring organic acids, such as malonate or oxalate, and then it diffuses away from the active site. Mn3+ organic acid chelates are strong diffusible oxidants capable of oxidizing a large variety of substrates, including phenolic lignin model compounds and several dyes.
Laccases (benzenediol:oxygen oxidoreductase, EC 1.10.3.2) are multicopper enzymes belonging to the blue oxidases that catalyse the one-electron oxidation of a wide variety of aromatic compounds (particularly phenols) with the reduction of oxygen to water. In a typical laccase reaction, a phenolic substrate is subjected to an one-electron oxidation giving rise to an aryloxyradical. This active species can be converted to a quinone in the second stage of oxidation.
White-rot fungi and their ligninolytic enzymes, with broad substrate specificity, have potential applications in biopulping and biobleaching, as well as in the bioremediation of aromatic pollutants with structural similarities to lignin. The use of lignin-degrading fungi or isolated ligninolytic enzymes for the treatment of pulp could potentially provide greater selectivity than chemical processes in the removal of lignin.
Other area where white-rot fungi and their ligninolytic enzymes can be useful is in bioremediation. Bioremediation is the process by which hazardous wastes are biologically converted to harmless compounds, or to levels that are below concentration limits. White-rot fungi have been found to degrade and mineralize xenobiotic compounds, such as polycyclic aromatic hydrocarbons, industrial dyes and other soil pollutants, such as atrazine and pentachlorophenol. Degradation and detoxification of these pollutants involve oxidative enzymes which are not very specific. Since lignin is a natural polyaromatic compound which is degraded by white-rot fungi using ligninolytic extracellular oxidative enzymes, it was naturally assumed that these organisms could degrade hazardous compounds using the same enzyme systems.
Pleurotus sp belongs to a subclass of white rot fungus that produces laccases,
Mn peroxidases, but not lignin peroxidase. One of the most important species,
Pleurotus pulmonarius (FR.) Quélet (P. sapidus, P. sajor-caju) was used in this study.
In the first work the removal of pentachlorophenol by Pleurotus pulmonarius
the laccase production from 32.1± 3.0 U/L in basal medium to 532 ± 3.0 U/L U/L, while no alteration in Mn peroxidase production was observed (18.0 ± 3.0 U/L). Approximately 70% of the initial PCP (25 ppm) was removed in submerged cultures under ligninolytic conditions, while not more than 20% of PCP was removed under non-ligninolytic conditions. The production of laccase appears to be important to improve the degradation of PCP by P. pulmonarius.
In the second work, the production of laccase and Mn peroxidase by P.
pulmonarius and the capability of cultures to decolourise the industrial dyes Remazol
Brilliant Blue R (RBBR), congo red, methylene blue, Poly R478 and ethyl violet were studied in solid state conditions. The condition of high initial moisture content (80-90%) strongly promoted the expression of laccase, while manganese peroxidase was produced specially in cultures developed at low initial moisture (50-70%). Cultures containing high manganese peroxidase activities were more efficient to decolourise congo red, ethyl violet and methylene blue. Cell free filtrates with high manganese peroxidase activity were more efficient to decolourise congo red, ethyl violet and methylene blue, while RBBR was equally decolourised by Mn peroxidase and laccase. No cell free extracts from P. pulmonarius was able to decolorize Poly R478.
Environmental pollutants are a serious concern world-wide. The results obtained in this study showed the potential applicability of Pleurotus pulmonarius for
the removal of aromatic compounds and wasterwater treatment of dye effluents.
Key words: Pleurotus pulmonarius, dye decolorisation, pentachlorophenol, ligninolytic enzymes, manganese peroxidase, bioremediation,
REMOVAL OF PENTACHLOROPHENOL BY Pleurotus pulmonarius IN SUBMERGED CULTURES UNDER LIGNINOLYTIC AND NON-LIGNINOLYTIC CONDITIONS
DANIELA FARANI DE SOUZA, CRISTINA GIATTI MARQUES DE SOUZA, MARIA APARECIDA FERREIRA COSTA, CINTHIA GANDOLFI BOER, MÁRCIA FERNANDES NISHIYAMA, HELOÍSA BRESSAN GONÇALVES AND ROSANE
MARINA PERALTA*
Departamento de Bioquímica, Universidade Estadual de Maringá, CEP 87015-900, Maringá– PR, Brazil.
ABSTRACT
The removal of pentachlorophenol (PCP) by Pleurotus pulmonarius grown in
submerged cultures under ligninolytic and non-ligninolytic conditions was studied in this work. Under ligninolytic conditions, the fungus was more resistant to PCP, removing 70% of PCP at an initial concentration of 25 mg/L, against a removal of 20% under non-ligninolytic conditions. PCP acted as a laccase inducer, and the existence of high activities of enzyme seems to be related to its degradation by P.
pulmonarius.
Key words: laccase, Pleurotus pulmonarius, xenobiotics, pentachlorophenol,
phenolic compounds, ligninolytic enzymes.
INTRODUCTION
most countries. In Brazil, its use as wood preservative was allowed until 2006 and it appears as one of the most common soil and wastewater pollutant in the country (Almeida et al., 2007). PCP is persistent with a half-life of up to 178 and 200 days in soil and water systems, respectively (Larson et al., 1997).
The possibility of using white-rot fungi to degrade PCP and related compounds has been focus of interest due to the presence of a ligninolytic system containing unspecific peroxidases (lignin peroxidase and Mn peroxidase) and laccase. Some fungi produce all these enzymes while other produce only one or two of them. It is generally believed that the same non-specific system responsible for lignin degradation by white-rot fungi, is also involved in the oxidation of aromatic pollutants (Lamar and Dietrich, 1990, Dritsa et al., 2007). The first studies on xenobiotic degradation by white rot fungi were carried out with Phanerochaete chrysosporium. It
was shown that this basidiomycete is able to degrade PCP and several other xenobiotic compounds in a nitrogen-limiting medium (ligninolytic condition) while degradation was reduced in high nitrogen medium (non-ligninolytic conditions) (McAllister et al., 1996; Mileski et al., 1988; Bending et al., 2002). However, there has been increasing interest in studying the capability of other white rot fungi to degrade PCP and other pollutants (Dritsa et al., 2007, Ryu et al., 2000, Marcial et al., 2005, Levin et al., 2003, Machado et al., 2005, Ullah et al., 2000a,b).
An increasing range of white rot fungi is now being investigated for bioremediation purposes (Machado et al., 2005; Alleman et al., 1995; Lamar and Dietrich, 1990, Mileski et al., 1988). Laccase appears as the main enzyme involved in the removal of xenobiotics including PCP in some white-rot species such as
Pleurotus and Coriolus (Ricotta et al., 1996, Ullah et al., 2000a, Ullah et al., 2000b,
Sedarati et al., 2003, Levin et al., 2003), while in other white-rot fungi such as
Nematoloma frowardii (Hofrichter et al., 1998, Sack et al., 1997), Phanerochaete.
chrysosporium (Moen and Hammel, 1994), Irpex lacteus (Baboravá et al., 2006) and
Bjerkandera sp (Eibes et al., 2007, Rubilar et al., 2007), the oxidation of xenobiotics is mainly due to the action of Mn peroxidase.
Laccase is described as the most important ligninolytic enzyme of P.
pulmonarius CCB19 in submerged cultures (Zilly et al., 2002). In submerged cultures
laccase by phenolic compounds is well studied in several basidiomycetes, including
Pleurotus sajor caju (Lo et al. 2001), Marasmus quercophilus (Farnet et al. 1999),
Pleurotus eryngii (Muñoz et al. 1997), and Trametes modesta (Nyanhongo et al.
2002).
Pleurotus spp are among the easiest mushrooms to cultivate (Cohen et al.,
2002). The specie most studied concerning its application in bioremediation processes is P. ostreatus. It shows high potential to degrade several organic
compounds including polycyclic aromatic hydrocarbons (PAH) and chlorophenols in liquid and solid state fermentation cultures (Bezalel et al., 1996a-c, Zeddel et al., 1993). Pleurotus spp, in contrast to P. chrysosporium, expresses the ligninolytic
system during the growth phase, and the production of ligninolytic enzymes is not inhibited by high nitrogen concentration. Other Pleurotus species have also
demonstrated potential in bioremediation processes (Rodrigues et al., 2004, Cohen et al., 2002). One of these species, Pleurotus pulmonarius (P. sajor-caju) when
cultured under submerged conditions produces laccase as the main extracellular enzyme (Souza et al., 2004, Zilly et al., 2002). In this type of cultures, it was able to decolourise textile industrial dyes (Zilly et al., 2002). Additionally, its capability to degrade atrazine in solid state cultures has also been described (Masaphy et al., 1996). In the present study, we examined the ability of Pleurotus pulmonarius to
grow in the presence of PCP in submerged cultures under ligninolytic and non-ligninolytic conditions. An attempt was done to correlate the removal of PCP with the production of laccase by the fungus.
2. MATERIALS AND METHODS
2.1. Organism
Pleurotus pulmonarius CCB19 was obtained from the São Paulo Botany
Institute Culture Collection. It was cultured on potato dextrose agar Petri dishes (PDA) for 2 weeks at 28oC. When the Petri dish was fully covered with the mycelia, mycelial plugs measuring 10 mm in diameter were made and used as inoculum for submerged cultures.
One disk from the growing edge of the mycelium on PDA plates was transferred to a 250 mL Erlenmeyer flask containing 50 ml of potato-dextrose medium. The cultures were incubated at 28 °C in a r otary shaker at 120 rpm for 5 days. Homogenized pellets from 5-day-old shaken cultures were used as inocula (corresponding to 1–2 mg dry weight per mL of culture).
2.3. Submerged ligninolytic and non-ligninolytic culture conditions
Cultures were carried out under ligninolytic and non-ligninolytic conditions. In non-ligninolytic conditions, homogenized pellets from 5 days-old shaken PD cultures (1-2 dry weight per ml of culture), were transferred to 25 mL of mineral medium (Montenecourt and Eveileigh, 1977) supplemented with glucose (10 g/L) and ammonium tartrate (0.86 g/L). To prepare the ligninolytic medium, mineral medium was firstly autoclaved with corn cob power (50 g of corn cob power plus 500 ml of mineral medium) for 15 min to extract soluble phenolic compounds. The mixture was filtered and the filtrate was then supplemented with glucose (10/L) and ammonium tartrate (0.86 g/L).
2.4. Rate of growth and PCP tolerance test
The fungus was tested for its growth rate in solid media containing pentachlorophenol (PCP). Ligninolytic and non-ligninolytic media (described above) were supplemented with agar (16 g/L) and PCP (1 to 30 mg/L) from a stock solution 1mg/mL in ethanol. Inocula (10 mm diameter) were cut with a cork borer from fully-grown PDA cultures and transferred into Petri dishes. Three replicates were performed for each PCP concentration.
2.5. Effect of PCP in submerged cultures
PCP was added in the 5th day of cultivation at a final concentration of 25 mg/L. The effect of PCP in submerged cultures was estimated by comparison of the dry biomass weight and the extracellular enzyme activities (Mn peroxidase and laccase) with control variants without PCP.
To evaluate the PCP content adsorbed on the mycelia obtained from submerged cultures, the biomass was filtered from culture, washed with distilled water, and immediately frozen at -20o C before freeze-drying. The resulting dried extract was ground to small particle-size before addition to ethanol in a 125 mL flask that was shaken at 120 rpm in an orbital shaker for 24 h. The filtrate was separated by centrifugation (15 min at 5000g) and used to quantify PCP by HPLC.
2.7. Analysis of PCP
2.8. Enzyme assays
The laccase activity was followed spectrophotometrically at 525 nm through the oxidation of syringaldazine to its quinine form, using a molar absorption coefficient of 65,000 for the product (Leonowicz and Grzywnowicz, 1981). The reaction mixture contained 1.5 mL phosphate buffer (0.1 M, pH 6.5), 0.2 mL syringaldazine (0.5 mM in ethanol solution) and 0.1 mL of culture filtrates. Mn peroxidase activity was assayed by the oxidation of 1 mM MnSO4 in 0.05 M sodium malonate , pH 4.5, in the presence of 0.1 mM H2O2. Manganic ions, Mn3+, form a complex with malonate, which absorbs at 270 nm (ξ270= 11.59 mM-1 cm-1) (Wariishi et al., 1992). Enzymic activity was expressed as international units (U) defined as the amount of enzyme required to produce 1 mol product per min.
2.9. Other chemical analysis
The total phenol content was determined colorimetrically at 750 nm, using the Folin-Ciocalteau reagent and expressed as ferulic acid equivalents. Total carbohydrates were determined by the phenol-sulfuric acid method with glucose as the standard (Dubois et al., 1956). Total protein content was estimated by the Bradford method using bovine serum albumin as the standard (Bradford, 1976). Total nitrogen was determined by the Kjeldahl standard method.
2.10. Statistical analyses
The data obtained were compared using paired t-test, and the level of significance of p<0.05 was chosen for all statistical comparisons. The data are presented as mean±SEM. The analysis was done using the statistical program pack GraphPad Prism® (Graph Pad Software, San Diego, USA).
2.11. Reagents
3. RESULTS
3.1. Growth of P. pulmonarius in submerged cultures under ligninolytic and non-ligninolytic conditions.
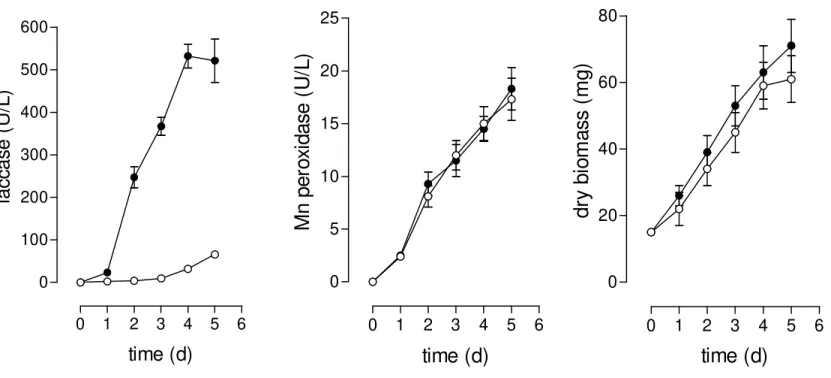
Previous results have shown that the supplementation of media with corn cob soluble phenolic compounds resulted in a considerable improvement of both, laccase production and capability of cultures to decolourise industrial dyes (Tychanowicz et al., 2004). For this reason, to obtain a ligninolytic condition in submerged culture, the same strategy was used in this work. The addition of soluble corn cob extract in the basal medium resulted in an improvement of phenolic content (as ferulic acid equivalents) from 0 (basal medium, non-ligninolytic condition) to 1.43 moles/ml (ligninolytic condition), without any significant alteration of carbon (determined as total carbohydrate and total protein contents) and nitrogen sources (determined as Kjeldahl nitrogen) (data not shown). The production of biomass and enzymes (laccase and Mn peroxidase) in both media is shown in Fig 1. A slight increase (statistically non-significant after application of t-test) in the production of biomass
was observed by the addition of soluble corn cob extract. The presence of corn cob soluble phenolic compounds improved the production of laccase more than 10 times (from 32.1±3.0 U/mL to 532±28.0 U/mL), while no significant alteration in the production of Mn peroxidase was observed (18.0±2.0 U/L).
3.2. Growth rates in solid media and PCP tolerance
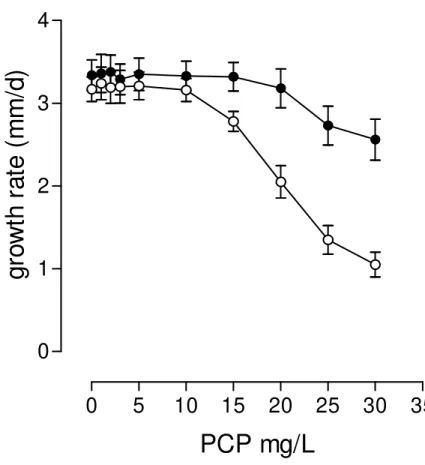
The growth rates of P. pulmonarius in Petri dishes were tested under
ligninolytic and non-ligninolytic conditions (FIG 2). The dishes were fully grown in 6-7 days. The growth rate measurements were analyzed by the least-squares method. The correlation coefficients R2 ranged from 0.96 to 0.99 in the three replicates. The growth rate of P. pulmonarius was 3.17±0.26 mm/d in non-ligninolytic medium and
3.34±0.32 mm/d in ligninolytic medium. In spite of the slight improvement at the growth rate under ligninolytic conditions, comparison of the growth in both conditions did not reveal significant differences (P>0.05). This type of cultivation was used to test the tolerance of P. pulmonarius to PCP. The ligninolytic condition improved the
whereas under the ligninolytic condition growth inhibition started at concentrations above 20 mg/L and was much less pronounced.
3.3. Effect of addition of PCP in submerged cultures under ligninolytic and non-ligninolytic conditions
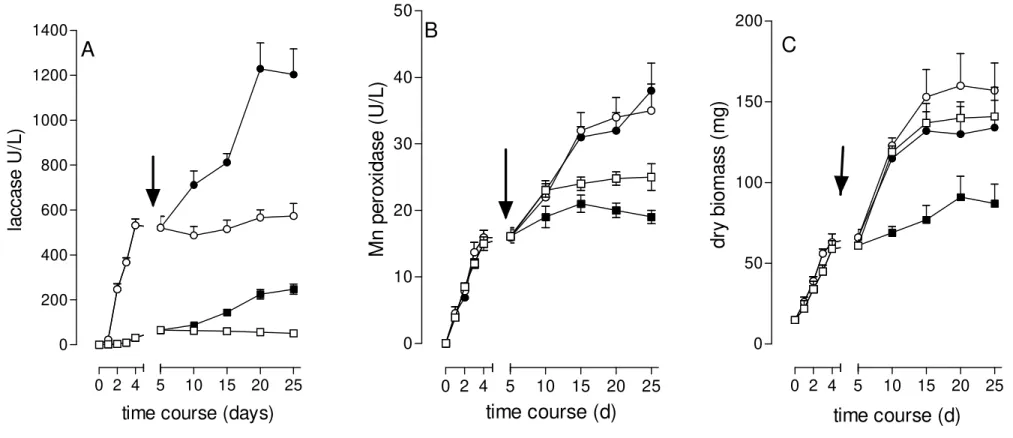
An amount of 25 mg/L of PCP (final concentration) was added to five-day-submerged cultures of P. pulmonarius developed under ligninolytic and
non-ligninolytic conditions. The addition of PCP in both types of cultures increased about 2 times the levels of laccase (from 600 U/L to 1,200 U/L) (Fig 3A), while no alteration was observed in the levels of Mn peroxidase (35 U/L) (Fig 3B). In relation to the biomass, PCP inhibited the growth of the fungus under both conditions, although the inhibition had been more evident under non-ligninolytic conditions (Fig 3C).
3.4. Evaluation of PCP removal
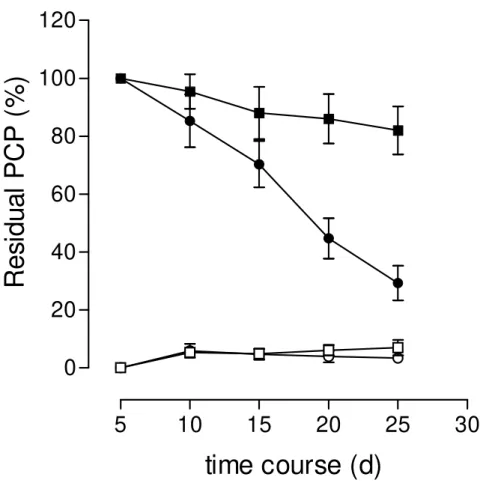
Figure 4 shows the removal of PCP by P. pulmonarius in ligninolytic and non-
ligninolytic submerged cultures. After 25 days of cultivation, around 70 and 20% of the initial PCP were removed in culture filtrates developed under ligninolytic and non-ligninolytic conditions, respectively. In both types of culture, less than 8% of PCP was found adsorbed for the fungal mycelium.
DISCUSSION
In this work, the supplementation of submerged cultures with corn cob extract had a very pronounced effect on the production of laccase by P. pulmonarius without
improvement in the levels of Mn peroxidase. In both, ligninolytic and non-ligninolytic media, maximal Mn peroxidase activities were found in 20th day cultures. These results are in agreement with those obtained in previous studies with the same strain
of P. pulmonarius, when low levels of Mn peroxidase were found in submerged
cultures (Souza et al. 2004). The laccase inducer effect of the soluble plant extract is considered to be related to its phenolic contents. Cotton stalk extract for example, acted as inducer of laccase in Pleurotus ostreatus submerged cultures (Platt et al.
Ganoderma applanatum, Trametes versicolor and Rhizoctonia solani (Ardon et al.
1996).
Our results also showed that PCP increased the production of laccase by P.
pulmonarius in submerged cultures, while the production of manganese peroxidase
was barely affected by the presence of PCP. In addition to this, the previous presence of a higher laccase activity apparently increased both the resistance of the fungus to PCP and its removal from the medium. Inductive aromatic compounds very often are toxic to fungalgrowth and metabolism, and it has been proposed that one of the possible functions of fungal laccase is the polymerization of toxic aromatic compounds (Thurston 1994). Therefore, laccase may function as a defense mechanism against oxidative stress. Some potential pollutants of the environment have been described as laccase inducers. In liquid cultures of Trametes versicolor,
the addition of several xenobiotics, including PCP at 0.5 mM, improved the laccase production (Mougin et al., 2002). In submerged cultures of T. versicolor, the addition
of 10 mg/L phenol improved 20 fold the production of laccase (Pazarhoglu et al., 2005). More recently, the efffects of paraquat dichloride (1,1′
-dimethyl-4,4′bipyridinium dichloride hydrate, methyl viologen dichloride), an effective contact
herbicide, were examined for its action on the laccase activity of Trametes versicolor
and Abortiporus biennis strains (Jaszek et al., 2006). Because of its prooxidative
character, paraquat is commonly used in laboratories as an oxidative stress conditioning factor. The addition of 25 M paraquat to T. versicolor and 20 M
paraquat to A. biennis cultures significantly stimulated the production of laccase.
CONCLUSION
Environmental pollutants are a serious concern world-wide. Many studies have shown that these persistent compounds can be degraded by the ligninolytic system of white-rot fungi. In this paper, a convenient submerged medium to study the capability of P. pulmonarius to remove PCP is proposed. Apparently, laccase is an
REFERENCES
Alleman BC, Logan BE, Gilbertson RL. 1992. Toxicity of pentachlorophenol to six species of white rot fungi as a function of chemical dose. Applied and Environmental Microbiology, 58: 4048-4050.
Almeida FV, Centeno AJ, Bisinoti MC, Jardim WF. 2007. Substâncias tóxicas persistentes (STP) no Brasil. Quim. Nova, 30, 1976-1985.
Apajalahti JHA, Salkinoja-Salonen MS. 1984. Absorption of pentachlorophenol (PCP) by bark chips and its role in microbial PCP degradation. Microbial Ecology, 10, 359-367.
Ardon O, Kerem Z, Hadar Y. 1996. Enhancement of laccase activity in liquid cultures of the ligninolytic fungus Pleurotus ostreatus by cotton stalk extract. J. Biotechnol.
51, 201–207.
Ardon O, Kerena Z, Hadar Y. 1998. Enhancement of lignin degradation and laccase activity in Pleurotus ostreatus. Can. J. Microbiol., 44, 670-680.
Baboravá P, Moder M, Baldrian P, Cajthamlová K, Cajthaml T. 2006. Purification of a new manganese peroxidase of the white-rot fungus Irpex lacteus and degradation
of polycyclic aromatic hydrocarbons by the enzyme. VChemosphere, 43, 167-182. Bending D, Friloux M, Walker A. 2002. Degradation of contrasting pesticides by white
rot fungi and its relationship with ligninolytic potential. FEMS Microbiol. Lett., 12, 59-63.
Bezalel L, Hadar Y, Fu PP, Freeman JP, Cerniglia CE. 1996b. Initial oxidation products in the metabolism of pyrene, anthracene, fluorine, and dibenzothiophene by the white-rot fungus Pleurotus ostreatus. Appl. Environ. Microbiol., 62,
25554-25559.
Bezalel L, Hadar Y, Cerniglia CE. 1996a. Mineralization of polycyclic aromatic hydrocarbons by the white-rot fungus Pleurotus ostreatus. Appl. Environ.
Microbiol., 62, 292-295.
Bradford MM. 1982. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 248-254.
Cohen R, Persky L, Hadar Y. 2002. Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol.,
58, 582-594.
Dritsa V, Rigas F, Natsis K, Marchant R. 2007. Characterization of a fungal strain isolated from a polyphenol polluted site. Bioresource Technology, 98, 1741-1747. Dubois M, Gilles KA, Hamilton JK, Rebers PA. 1956. Colorimetric method for
determination of sugars and related substances. Anal. Chem., 28, 350-356
Eibes G, Moreira MT, Feijoo G, Daugulis AJ, Lema JM. 2007. Operation of a two-phase partitioning bioreactor for the oxidation of anthracene by the enzyme manganese peroxidase Chemosphere. 66, 1744-1751.
Farnet AM, Tagger S, Petit JL. 1999. Effects of copper and aromaric inducers on the laccases of the white-rot fungus Marasmiusquercophilus. Plant Biol Pathol., 322,
499-503.
Hofrichler M, Scheibner K, Schneegab K, Fritsche W. 1998. Enzymatic combustion of aromatic and aliphatic compounds by manganese peroxidase from Nematoloma
frowardii. Appl. Environ. Microbiol., 64, 399-404
Jaszek M, Grzywnowicz K, Malarczyk E, Leonowicz, A. 2006. Enhanced extracellular laccase activity as a part of the response system of white rot fungi
Lamar RT, Dietrich DM. 1990. In situ depletion of pentachlorophenol from
contamined soil by Phanerochate spp. Applied and Environmental Microbiology,
56: 3519-3526
Larson SJ, Capel, PD, Majewski MS. 1997. Pesticides in surface waters. Ann Arbor Press, Inc, Michigan
Leonowicz A, Grzywnowicz K. 1981. Quantitative estimation of laccase forms in some white-rot fungi using syringaldazine as a substrate. Enz. Microb. Technol., v.
3, p. 55-58.
Levin l. Viale A, Forchiassin A. 2003. Degradation of organic pollutants by the white rot basidiomycete Trametes trogii. International Biodeterioration and
Biodegradation, 52, 1-5
Lo ASC, Ho YS, Buswell JA. 2001. Effect of phenolic monomers on the production of laccases by the edible mushroom Pleurotus sajor-caju and partial characterization
of a major laccase component. Mycologia, 93, 413-421.
Marcial J, Barrios-González J, Tomasini A. 2005. Effect of medium composition on pentachlorophenol removal by Amylomyces rouxii in solid-state culture. Process
Biochemistry, 41, 496-500
Masaphy S, Levanon D, Henis Y. 1996. Degradation of atrazine by the lignocellulosic fungus Pleurotus pulmonarius during solid-state fermentation. Bioresource
Technology, 56, 207-214
McAllistter KA, Lee H, Trevors JT. 1996. Microbial degradation of pentachlorophenol. Biodegradation, 7, 1-40
Mileski GJ, Bumpus JA, Jurek SD, Aust SD. 1988. Biodegradation. Appl. Environ. Microbiol., 54, 2885-2889Moen, M.A., Hammel, K.E. 1994. Lipid peroxidation by the manganese peroxidase of Phanerochaete chrysosporium is the basis for phenanthrene oxidation by the intact fungus. Appl. Environ. Microbiol., 60, 1956-1961
Moen MA, Hammel KE. 1994. Lipid peroxidation by the manganese peroxidase by
Phanerochaete chrysosporium is the basis for phenanthrene oxidation by the
intact fungus. Appl. Environ, Microbiol., 60, 1956-1961
Montecourt BS; Eveileigh DE. Preparation of mutants of Trichoderma reesei with
enhanced celulase production. Appl. Environ. Microbiol., v.34, p. 1-5, 1977.
Mougin C, Kollmann A, Jolivalt C. 2002 Enhanced production of laccase in the fungus Trametes versicolor by the addition of xenobiotics. Biotechnol Lett 24, 139–
142
Muñoz C, Guillen F, Martinez AT. 1997. Induction and characterization of lacases in the ligninolytic fungus Pleurotus eryngii. Curr. Microbiol., 34, 1-5.
Nyanhongo GS, Gomes J, Gubitz G, Zvauya R, Read JS, Steiner W. 2002. Production of laccase by a newly isolated strain of Trametes modesta. Biores.
Technol., 84, 259-263, 2002.
Pazarhoglu N, Sariisik M, Telefoncu A. 2005. Laccase: production by Trametes
versicolor and application to denim washing. Process Biochemistry, 40, 1673-1678
Platt MW, Hadar Y, Chet I. 1984. Fungal activities involved in lignocellulose degradation by Pleurotus. Appl. Microbiol. Biotechnol., 20, 150-154.
Ricotta A, Unz RF, Bollag JM. 1996. Role of laccase in the degradation of pentachlorophenol. Bull Environ Contam Toxicol, 57, 560-567
Rodrigues E, Nuero O, Guillen F, Martinez AT, Martinez MJ. 2004. Degradation of phenolic and non-phenolic aromatic pollutants by four Pleurotus species: the role
Ryu WR, Shim SH, Jang MY, Jeon YJ, Oh KK, Cho MH. 2000. Biodegradation of pentachlorophenol by white-rot fungi under ligninolytic and nonligninolytic conditions. Biotechnol. Bioprocess Eng., 5, 211-214
Rubilar O, Feijoo G, Diez C, Lu-Chau TA, Moreira MT, Lema JM. 2007. Biodegradation of pentachlorophenol in soil slurry cultures of Bjerkandera adusta and Anthracophyllum discolor. Ind. Eng. Chem. Res., 46, 6744-6751
Sack U, Hofrichter M, Fritsche W. 1997. Degradation of polycyclic aromatic hydrocarbons by manganese peroxidase of Nematoloma frowardii. FEMS
Micrbiology Letters 152, 227–334.
Sedarati MR, Keshavarz T, Leontievski AA, Evans CS. 2003. Transformation of high concentrations of chlorophenols by the white-rot basidiomycete Trametes
versicolor immobilized on nylon mesh. Electronic Journal of Biotechnology, 6
Semple KT, Reid BJ, Fermor TR. 2001. Impact of composting strategies on the treatment of soil contamined with organic pollutants. Environmental Pollution, 112, 269-283
Souza CGM, Tychanowicz GK, Souza DF, Peralta RM. 2004. Production of laccase isoforms by Pleurotus pulmonarius in response to presence of phenolic and
aromatic compounds. J. Basic Microbiol., 44, 129-136
Thurston CF. The structure and function of fungal laccases. Microbiology, 140, 19-26, 1994.
Tychanowicz GK, Zilly A, Souza CGM, Peralta, RM 2004. Decolourisation of industrial dyes by solid state cultures of Pleurotus pulmonarius. Proc. Biochem,
39, 855-859.
Ullah MA, Bedford CT, Evans CS. 2000a. Reactions of pentachlorophenol with laccase from Coriolus versicolor. Appl. Microbiol. Biotechnol., 53, 230-234
Ullah MA, Kadhim H, Rastall RA, Evans CS. 2000b. Evaluation of solid substrates for enzyme production by Coriolus versicolor, for use in bioremediation of
chlorophenols in aqueous effluents. Appl. Microbiol. Biotechnol., 54, 832-837. Wariishi H, Valli K, Gold MH. 1992. Manganese (II) oxidation by manganese
peroxidase from the basidiomycete Phanerochate chrysosporium. J. Biol. Chem.,
267, 23688-23695
Zeddel A, Majcherczyk A, Huttermann A. 1993. Degradation of polychlorinated biphenyls by white-rot fungi Pleurotus ostreatus and Trametes versicolor in a
Zilly A, Souza CGM, Peralta RM. 2002. Decolorisation of industrial dyes ba a Brazilian strain of Pleurotus pulmonarius producing laccase as the sole
0 1 2 3 4 5 6 0 100 200 300 400 500 600 time (d) la cc as e (U /L )
0 1 2 3 4 5 6 0 5 10 15 20 25
time (d)
M
n
pe
ro
xi
da
se
(
U
/L
)
0 1 2 3 4 5 6 0 20 40 60 80
time (d)
dr
y
bi
om
as
s
(m
g)
Figure 1. Production of laccase and Mn peroxidase and biomass by P. pulmonarius. The cultures were developed in ligninolytic (●) and
0
5
10 15 20 25 30 35
0
1
2
3
4
PCP mg/L
gr
ow
th
r
at
e
(m
m
/d
)
Figure 2. Growth rates of P. pulmonarius in ligninolytic (●) and non-ligninolytic (○)
0 2 4 0 200 400 600 800 1000 1200 1400
5 10 15 20 25
time course (days)
la cc as e U /L )
0 2 4 0 10 20 30 40 50
5 10 15 20 25
time course (d)
M n pe ro xi da se ( U /L )
0 2 4 0
50 100 150 200
5 10 15 20 25
time course (d)
dr y bi om as s (m g)
A
B
C
Figure 3. Effect of PCP in submerged cultures of P. pulmonarius developed under ligninolytic and non-ligninolytic conditions. The arrow
indicates the time at which PCP was introduced in the cultures at the final concentration of 25 mg/L. (●) ligninolytic cultures with PCP;
5
10
15
20
25
30
0
20
40
60
80
100
120
time course (d)
R
es
id
ua
l P
C
P
(
%
)
Figure 4. Residual PCP in culture filtrates of P.pulmonarius developed under
ligninolytic (●) and non-ligninolytic conditions (■). (□) amount of PCP adsorbed to the
mycelial mass developed under non-ligninolytic conditions; (○) amount of PCP
THE RATIO Mn PEROXIDASE/LACCASE IS HIGHLY AFFECTED BY THE INITIAL MOISTURE CONTENT ON CORN COB SOLID STATE CULTURES OF
Pleurotus pulmonarius
DANIELA FARANI DE SOUZA, CISSA KELMER BRACHT, ANA MARIA ALEXANDRINO, GISELE PEZENTE FERREIRA, MARIA APARECIDA FERREIRA
COSTA, CINTHIA GANDOLFI BOER, CRISTINA GIATTI MARQUES DE SOUZA and ROSANE MARINA PERALTA
Departamento de Bioquímica, Universidade Estadual de Maringá
ABSTRACT
The production of ligninolytic enzymes by the white-rot fungus Pleurotus pulmonarius
(FR.) Quélet and the capability of cultures to decolourise the industrial dyes Remazol Brilliant Blue R (RBBR), congo red, methylene blue and ethyl violet were studied in this work. The condition of high initial moisture content (80-90%) strongly promoted the expression of laccase, while manganese peroxidase was produced specially in cultures developed at low initial moisture (50-70%). Cultures containing high manganese peroxidase activities were more efficient to decolorize the four dyes than those containing high laccase activity. Cell free filtrates with high manganese peroxidase activity were also more efficient to decolorize methylene blue, ethyl violet and congo red, while the dye RBBR was efficiently decolorized by both enzymes, Mn peroxidase and laccase.
Key words: Pleurotus pulmonarius, ligninolytic enzymes, dye decolorization,
industrial dyes
1. INTRODUCTION
(Nigam et al., 2000). There are more than 100,000 commercially available dyes with over 7×105 ton of dye-stuff produced annually (Zollinger, 1987). Due to their chemical structure, dyes are resistant to fading on exposure to light, water and many chemicals (Poots and McKay, 1976 and McKay, 1979). Many dyes are difficult to decolourise due to their complex structure and synthetic origin. There are many structural varieties, such as, acidic, basic, disperse, azo, diazo, anthroquinone based and metal complex dyes. Decolorization of textile dye effluents does not occur when they are treated aerobically by municipal sewerage systems (Willmott et al., 1998).
Ligninolytic fungi have been studied as possible agent of biodegradation because their extracellular degradation systems are basically non-specific, a fact that allows the degradation of mixtures of refractory substances (Kirk and Farrell, 1987). The major enzymes associated with the lignin-degrading ability of white-rot fungi are lignin peroxidase (EC 1.11.1.14), manganese peroxidase (EC 1.11.1.13) and laccase (EC 1.10.3.2) (Vicuna, 2000). Some white-rot fungi produce all these enzymes while others produce only one or two of them (Leonowicz et al., 2001). Several strains of the dye- or colored material-degrading microorganisms have been reported, including species of Pleurotus, Coriolus, Trametes, Polyporus, Lentinus and Picnoporus
(Leonowicz et al., 2001, Sani et al., 1998, Kahraman and Yesilada, 1999, Das et al., 2000, Sam and Yesilada, 2001, Zilly et al., 2002). The capability of dye decolorization may be due to the activities of lignin peroxidase (Young and Yu, 1997, Ollikka et al., 1993), Mn peroxidase (Boer et al., 2004, Heinfling et al., 1998, Hatvani and Mecs, 2002), and laccase (Wong and Yu, 1999, Zilly et al., 2002, Pointing and Vrjmoed, 2000, Landelbauer et al., 2004, Rodriguez et al., 1999).
Pleurotus spp. are among the easiest mushrooms to cultivate (Cohen et al.
2002). The two most important species cultivated on an industrial scale are P.
ostreatus and P. pulmonarius (P. sajor-caju, P. sapidus). In nature they grow on
wood, usually on dead, standing trees or on fallen logs. Various substrates that contain lignin and cellulose can be used for Pleurotus cultivation, such as wood
chips, corn wheat, rice straw, cotton stalks, waste hulls, and other agricultural wastes, some of which can be recycled and upgraded for use as animal feed or for preparation of other products (Cohen et al. 2002). These substrates are frequently used to study the production of MnP and laccase by Pleurotus spp in both
2003). It mimics the natural environment of the white-rot fungi and holds tremendous potential for the production of enzymes, including laccase and MnP (Pandey et al.
1999).
Pleurotus pulmonarius (P. sajor-caju) when cultured under submerged and
solid state conditions using wheat bran as substrate produces laccase as the main extracellular enzyme (Souza et al., 2002). The capability of P. pulmonarius to
decolourise textile dyes in both types of cultures has already been described and associated with the production of laccase considering that the production of Mn peroxidase in these cultures was very low (Zilly et al., 2002, Tychanowicz et al, 2004). More recently however, it has been described that when P. pulmonarius was
cultured in wheat bran solid state medium at low initial moisture content, it produced both enzymes, Mn peroxidase and laccase, in elevated amounts (Souza et al., 2006). The objectives of this study were to compare the production of laccase and Mn peroxidase by P. pulmonarius in solid state cultures using three different substrates,
wheat bran, corn cob and pineapple peel at different initial moisture contents and to evaluate the capability of these cultures to decolorize some industrial dyes.
2. MATERIAL AND METHODS
2.1. Microorganism
Pleurotus pulmonarius CCB-19 was obtained from the Culture Collection of
the Botany Institute of São Paulo. It was cultured on potato dextrose agar (PDA) medium for 2 weeks at 28 °C. When the plates were f ully covered with the mycelia, mycelial plugs measuring 10 mm in diameter were made and used as inocula.
2.2. Enzyme production on SSC
P. pulmonarius CCB-19 was cultivated in 250 ml Erlenmeyer flasks containing
5 g of wheat bran, corn cob or pineapple peel. The following salts were added to give
a final salt concentration of (mg/g): K2HPO4, 1; MgSO4 7H2O, 0.2 and CaCl2 2 H2O,
cultures were inoculated aseptically by using three mycelial plugs obtained from the colony edge of a colony grown on PDA medium. Incubation was at 28 °C.
2.3. Enzyme extraction
Crude extract was obtained by adding 50 ml of cold water to the contents of each flasks, stirring for 1 h at 4 °C, and using fi ltration and centrifugation. The supernatant was stored at –20 °C until assay.
2.4. Dye decolorization experiments on SSC
To test the ability of cultures to decolourise industrial dyes, each dye was membrane-filtered through a 0.45 m cellulose nitrate filter and mixed with the corn cob medium, previously autoclaved, to a final concentration of 200 ppm. After 15 days, the residual dyes in the cultures were extracted firstly with 50 ml of water followed by 50 ml of a mixture of methanol:acetone:water (1:1:1). Dye disappearance was determined spectrophotometrically by monitoring the absorbance at the wavelength of maximum absorbance for each dye. In control cultures, either dye or the fungus (abiotic control) was omitted. To calculate the residual dye in the cultures, the total dye extracted with water and organic mixture in the abiotic control was considered as 100%. The amount of adsorbed dye on corn cob medium after growth of the fungus was always less than 10%.
2.5. Dye decolourisation by culture filtrates
2.6. Enzyme assays
All enzyme assays were performed at 40 °C. Laccase activity was followed spectrophotometrically at 525 nm, by the oxidation of syringaldazine to its quinone form, using a molar absorptivity of 65,000 for the product (Leonowicz and Grzywnowicz 1981). The reaction mixture contained 1.5 ml phosphate buffer (0.1 M, pH 6.5), 0.2 ml syringaldazine (0.5 mM in ethanol solution), and 0.1 ml of culture filtrates. The MnP activity was assayed by the oxidation of 1 mM MnSO4 dissolved in 0.05 M sodium malonate, pH 4.5, with 0.1 mM H2O2. The absorption of the complex resulting from this reaction was measured at 270 nm (ε270 = 11.59 mM–1cm–1) (Wariishi et al. 1997).
2.7. Dyes
The following dyes were used in this work: one anthracene derivative, Remazol brilliant blue (RBBR); one triphenylmethane dye, Ethyl violet; one heterocyclic dye, Methylene blue; one polymeric dye, Poly R-478 and one azo dye, Congo red
2.8. Statistical analysis
The data obtained were compared using paired t-test, and the level of
significance of p<0.05 was chosen for all statistical comparisons. The data are presented as mean ± SEM. The analysis was done using the statistical program pack GraphPad Prim® (Graph Pad Software, San Diego, USA).
2.9. Chemicals
3. RESULTS
3.1. Effect of substrate and initial moisture content (IMC) in the production of P.
pulmonarius enzymes
The production of ligninolytic enzymes by P. pulmonarius CCB-19 was
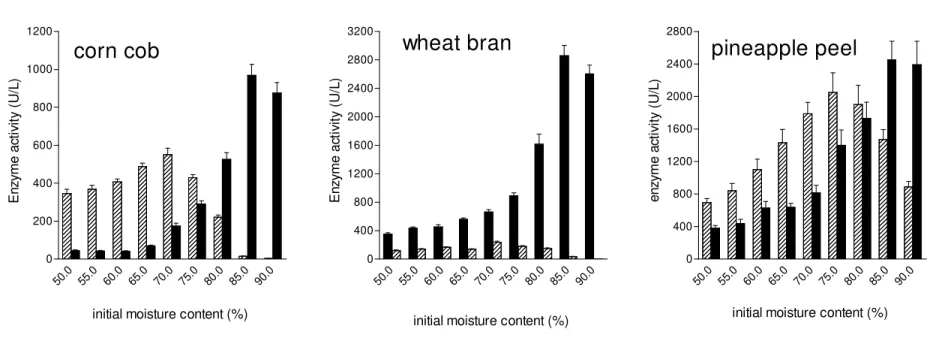
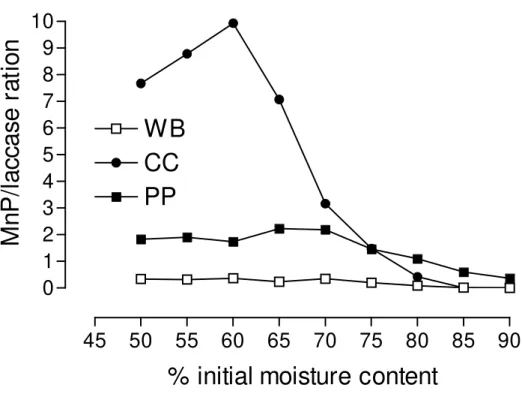
determined using three different substrates at different initial moisture contents (Fig. 1). Laccase was the main ligninolytic enzyme produced by the fungus in wheat bran cultures and its production was positively affected by increases in initial moisture content (Fig. 1B). In pineapple peel cultures both enzymes were produced at high amounts, being high initial moisture content (80-90%) the best condition to produce laccase, and low initial moisture content (70-75%), the best condition to produce Mn peroxidase. The substrate where the initial moisture content had the strongest effect in the production of enzymes was corn cob (Fig 1A). Initial moisture content of 85-90% was the best condition to produce laccase and very low Mn peroxidase activity was detected in these filtrates. The best initial moisture contents to produce Mn peroxidase changed from 50 to 65%, a condition where the production of laccase was very low (Fig 1A). The impact of initial moisture content on the relative production of both enzymes in these cultures can be best observed in Fig 2. The Mn peroxidase/laccase ratio changed from 9.93 (at IMC of 60%) to less than 0.01 (at IMC of 90%) in corn cob cultures. The lowest MnP/laccase ratio was found in wheat bran cultures (ranged from 0.37 to 0.002) followed by pineapple peel cultures (ranged from 2.23 to 0.37).
3.2. Effect of initial moisture content in capability to decolorize industrial dyes by corn cob cultures of P. pulmonarius
The industrial dyes used in this work were selected on the basis of their stability over a wide range of pH (3-11), thermostability, stability under culture conditions in non-inoculated flasks and they were representative for each dye chemical category (anthracene derivative, azo, heterocyclic, polymeric and triphenylmethane dyes).
than the cultures developed at 85% of initial moisture content (p<0.05). The dye Poly R478 was barely decolorized by cultures in both moisture content cultures. Alcoholic extracts from mycelia and corn cob showed that less than 10% of the dyes were adsorbed by the mixture of fungi plus corn cob. RBBR was equally decolourized by cultures developed at initial moisture contents of 60 and 85%.
3.3. Capability of crude dialyzed cell free extracts from P. pulmonarius solid state cultures decolorize industrial dyes with high and low MnP/lcc ratio
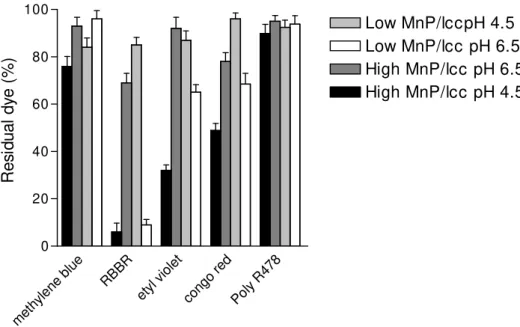
To test the ability of crude-dialysed cell free extracts to decolourise the industrial dyes used in this study, the extracts were incubated with 100 ppm of dyes under two different conditions, to obtain the best condition for Mn peroxidase activity (50 mM malonate buffer pH 4.5 with 1 mM MnSO4 and 0.1 mM H2O2 ) and the best condition for laccase activity (50 mM phosphate buffer, pH 6.5). The results are shown in Figure 3. Both cell free extracts (from 60% and 85% initial moisture content cultures) efficiently decolourised RBBR. No cell free extracts were able to decolourize Poly R478, while the cell free extracts obtained from cultures developed with initial moisture content of 60% were more efficent to decolourize the dyes methylene blue, ethyl Violet and congo red.
DISCUSSION
Many white rot fungi have been intensively studied in connection with their ligninolytic enzyme production and their decolorization ability (Boer et al., 2004, Pointing and Vrjmoed, 2000, Kasinath et al., 2003, Chagas and Durrant, 2001, Jarosz-Wilkolazka et al., 2002). However, most studies on dye decolorization have been carried out using liquid or solid cultures on agar plates, which do not reflect the natural living conditions (i.e. in wood and other lignocellulosic substrates) of white-rot fungi. Solid state culture was chosen here because it mimics the natural environment of the white-rot fungi and high levels of ligninolytic enzymes are generally produced in this type of cultivation. The list of different substrates used for the cultivation of
Pleurotus sp is long, including several agricultural materials, such as wheat bran,
and Mn peroxidases (Rosales et al., 2002, Songulashvili et al., 2006, Alexandrino et al., 2007).
In this work, pineapple peel, a yet not used substrate in this type of study was introduced and tested as substrate for P. pulmonarius. In pineapple peel solid state
cultures both, laccase and Mn peroxidase were produced at levels higher than in corn cob and wheat bran cultures. A previous study has demonstrated the ability of
P. pulmonarius CCB19 to grow and produce laccase as the main ligninolytic enzyme
on SSC using wheat bran with a moisture content of 75% as the substrate (Souza et
al. 2002). In the present work, we showed that the cultivation of P. pulmonarius
under solid state conditions using corn cob and pineapple peel as substrates resulted in a convenient condition to produce also high amounts of Mn peroxidase. By varying only the initial moisture content, the use of corn cob as substrate allowed the obtainment of cell free extracts rich either in laccase either in Mn peroxidase. In addition to this, the low amounts of natural colored pigments in this material, permitted the use of its cell free extracts in experiments of decolorization without any additional treatments, because the color of the extracts do not interfere with the determination of residual dyes. From the results obtained in this work, it is possible to conclude that Mn peroxidase was the main enzyme responsible for the decolourisation of congo red, ethyl violet and methylene blue by P. pulmonarius. Our
data suggest that both enzymes laccase and Mn peroxidase from P. pulmonarius are
equally efficient in decolorize RBBR. To detect the real potential of enzymes to decolorize industrial dyes, it is necessary to obtain isolated fractions of these enzymes. The main laccase from P.pulmonarius was recently purified (Souza et al.,
2003). Purification of Mn peroxidase is in progress in the laboratory.
ACKNOWLEDGEMENTS
REFERENCES
Alexandrino, A.M., Faria, H.G., Souza, C.G.M., Peralta, R.M. 2007. Reutilisation of orange waste for production of lignocellulolytic enzymes by Pleurotus ostreatus.
Cienc. Tecnol. Aliment., 27, 364-368
Banat, I.M., Nigam, P., Singh, D. and Marchant, R., 1996. Microbial decolorization of textile-dye-containing effluents: a review. Bioresour. Technol. 58, 17–227.
Boer, C.G., Obici, L., Souza, C.G.M., Peralta, R.M. 2004. Decolorization of synthetic dyes by solid state cultures of Lentinula (Lentinus) edodes producing manganese
peroxidase as the main ligninolytic enzyme. Biores. Technol., 94, 107-112
Chagas, E.P., Durrant, L.R. 2001. Decolorization of azo dyes by Phanerochaete
chrysosporium and Pleurotus sajor-caju. Enz. Microbiol. Technol. 29, 473–477.
Cohen, R., Persky, L., Hadar, Y., 2002. Biotechnological applications and potential of wooddegrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol.,
58, 582–594.
Das, N., Chakraborty, T.K., Mukherjee, M. 2000. Purification and characterization of laccase 1 from Pleurotus florida. Folia Microbiol., 45, 447-451
Gervais, P. and Molin, P., 2003. The role of water in solid-state fermentation. Biochem. Eng. J., 13, 85–101.
Hatvani, N., Mecs, I. 2002. Effect of the nutrient composition on dye decolourisation and extracellular enzyme production by Lentinus edodes on solid medium. Enz. Microbiol. Technol., 30, 381-386
Heinfling, A., Martinez, M.J., Martinez, A.T., Bergbauer, M., Szewzyl, U. 1998. Transformation of industrial dyes by manganese peroxidases from Bjerkandera
adusta and Pleurotus eryngii in a manganese-independent reaction. Appl.
Environ. Microbiol., 64, 2788-2793.
Jarosz-Wilkołazka, A., Rdest-Kochmańska, J., Malarczyk, E., Wardas, W.,
Leonowicz, A. 2002. Fungi and their ability to decolorize azo and anthraquinonic dyes. Enzyme Microbiol. Technol. 30, 566–572.
Juang, R.S., Tseng, R.L., Wu, F.C. and Lin, S.J., 1996. Use of chitin and chitosan in lobster shell wastes for colour removal from aqueous solutions. J. Environ. Sci. Health A 31, 325–338.
Kasinath, A., Novotný, C., Svobodová, K., Patel, K.C. and Sasek, V., 2003. Decolorisation of synthetic dyes by Irpex lacteus in liquid cultures and packed-bed
bioreactor. Enzyme Microbiol. Technol., 32, 167–173.
Kirk, T.K., Farrell, R. 1987. Enzymatic “combustion”: the microbial degradation of lignin. Ann. Rev. Microbiol., 41, 465-505
Landelbauer, A., Maute, O., Kessler, R.W., Erlacher, A., Gubitz, G.M. 2004. Study of dye decolorization in an immobilized laccase enzyme-reactor using on line spectroscopy. Biotechnol Bioeng., 87, 552-563
Leonowicz, A. and Grzywnowicz, K., 1981. Quantitative estimation of laccase forms in some white rot fungi using syringaldazine as a substrate. Enz. Microb. Technol., 3, 55–58.
Leonowicz, A., Cho, N.-S., Luterek, J., Wilkolazka, A., Wojtas-Wasilewska, M., Matuszewska, A., Hofrichter, M., Wesenberg, D., Rogalski, J. 2001. Fungal laccase: properties and activity on lignin. J. Bas. Microbiol., 41, 185-227.
McKay, G., 1979. Waste colour removal from textile effluents. Am. Dyes. Rep. 68,
29–36
Mishra, G. and Tripathy, M., 1993. A critical review of the treatments for decolourization of textile effluent. Colourage, 40, 35–38.
Moo-Young, M., Moreira, R. P., Tengerdy, R. P., 1983. Principles of solid state fermentation. In: J. E. Smith, D. R. Berry, B. Kristiansen (Eds.), The Filamentous Fungi, vol. 4, Fungal Technology,
Nigam, P., Armour, G., Banat, I.M., Singh, D. and Marchant, R., 2000. Physical removal of textile dyes and solid state fermentation of dye-adsorbed agricultural residues. Bioresour. Technol. 72, 219–226.
Ollikka, P., Alhonmaki, K., Leppanen, V-L., Glumoff, T.R., Suominnen, I. 1993. Decolorization of azo, triphenylmethane, heterocyclic, and polymeric dyes by lignin peroxidase isoenzymes from Phanerochaete chrysosporium. Appl. Environ.
Microbiol., 59, 4010-4016
Pandey, A., Selvakumar, P., Soccol, C. R., Nigam P., 1999. Solid state fermentation for the production of industrial enzymes. Curr. Sci., 77, 149–162.
Pointing, S. B., Vrjmoed, L. L. P., 2000. Decolorization of azo and triphenylmethane dyes by Picnoporus sanguineus producing laccase as the sole phenoloxidase.
World J. Microbiol., 16, 317– 318.
Rodriguez, E., Pickard, M. A., Vazquez-Duhalt, R., 1999. Industrial dye decolorization by laccases from ligninolytic fungi. Cur. Microbiol., 38, 27–32.
Rosale, E., Rodriguez Couto, S., Sanroman, A. 2002. New uses of food waste: application to laccase production by Trametes hirsuta. Biotechnol. Lett., 24,
701-704
Sam, M., Yesilada, O. 2001. Decolorization of orange II dye by white-rot fungi. Folia Microbiol., 46, 143-146
Sani, R.K., Azmi, W., Banerjee, U.C. 1998. Comparison of static and shake culture in the decolorization of textile dyes and dye effluents by Phanerochaete
chrysosporium. Folia Microbiol., 43, 85-88
Songulashvili, G., Elisashvili, V., Wasser, S., Nevo, E., Hadar, Y. 2006. Laccase and manganese peroxidase activities of Phellinus robustus and Ganoderma
adspersum grown on food industry wastes in submerged fermentation.
Biotechnol. Lett., 28, 1425-1429
Souza, C. G. M., Zilly, A., Peralta, R. M., 2002. Production of laccase as the sole phenoloxidase by a Brazilian strain of Pleurotus pulmonarius. J. Bas. Microbiol.,
42, 83–90.
Souza, C.G.M, Peralta, R.M. 2003. Purification and characterization of the main laccase produced by the white-rot fungus Pleurotus pulmonarius on wheat bran
solid state médium. J. Bas. Microbiol., 43, 278-286
Vicuna, R. 2000. Lignininolysis. A very peculiar microbial process. Mol. Biotechnol., 14, 173-176
Wariishi, H., Valli, K., Gold, M. H., 1992. Manganese (II) oxidation by manganese peroxidase from the basidiomycete Phanerochaete chrysosporium. J. Biol. Chem.
267, 23688–23695.
Willmott, N., Guthrie, J. and Nelson, G., 1998. The biotechnology approach to colour removal from textile effluent. JSDC 114, 38–41
Wong, Y., Yu, J. 1999. Laccase-catalyzed decolorization of synthetic dyes. Water Research, 36, 3512-3520
Young, l., Yu, L. 1997. Ligninase-catalyzed decolorization of sybthetic dyes. Wat. Res., 5, 1187-1193
50.0 55.0 60.0 65.0 70.0 75.0 80.0 85.0 90.0 0 200 400 600 800 1000 1200
initial moisture content (%)
E nz ym e ac tiv ity ( U /L )
50.0 55.0 60.0 65.0 70.0 75.0 80.0 85.0 90.0
0 400 800 1200 1600 2000 2400 2800 3200
initial moisture content (%)
E nz ym e ac tiv ity ( U /L )
50.0 55.0 60.0 65.0 70.0 75.0 80.0 85.0 90.0
0 400 800 1200 1600 2000 2400 2800
initial moisture content (%)
en zy m e ac tiv ity ( U /L )
corn cob
wheat bran
pineapple peel
Figure 1. Effect of initial moisture content in the production of laccase and Mn peroxidase by P. pulmonarius in solid state cultures. The
45 50 55 60 65 70 75 80 85 90
0
1
2
3
4
5
6
7
8
9
10
WB
CC
PP
% initial moisture content
M
nP
/la
cc
as
e
ra
tio
n
Figure 2. Effect of initial moisture content in the ratio Mn peroxidase/laccase of P.
Table 1. Degree of decolorization of industrial dyes by solid state cultures of P.
pulmonarius at two initial moisture contents.
Synthetic dye common name Λ
(nm)
Residual dye after 15 days of cultivation 60% IMC cultures
MnP
85% IMC cultures Laccase Anthracene derivative dye
RBBR
595 3.7±2.8(a) 4.4±2.2(a)
Azo dyes Congo red
497 22.0±5.0(a) 47.3±5.6(b)
Heterocyclic dye methylene blue
665 52.1±5.7(a) 67.9±7.3(b)
Triphenylmethane dye ethyl violet
596 39.5±6.2(a) 62.5±2.3(b)
Polymeric dye Poly R478
530 83.8±6.8(a) 91.8±7.2(a)
methy lene
blue RBBR
etyl violet
cong o re
d
Poly R478
0 20 40 60 80 100
Low MnP/lccpH 4.5 Low MnP/lcc pH 6.5 High MnP/lcc pH 6.5 High MnP/lcc pH 4.5
R
es
id
ua
l d
ye
(
%
)
Figure 3. In vitro decolorization of industrial dyes by crude dialyzed cell free extracts
from P. pulmonarius solid state cultures. The dyes were added to the reaction
medium to give a final concentration of 100 ppm. The mixtures were maintained at 40º C for 2 h. The reaction mixture with low MnP/lcc ratio contained 2.1 U/L of manganese peroxidase activity and 97.0 U/L of laccase activity. The reaction mixture with high MnP/lcc ratio contained 81.2 U/L of manganese peroxidase activity and 8.0 U/L of laccase activity.