Influência do ambiente na higidez de larvas de
engraulidídeos coletadas na região de Santos (SP)
São Paulo 2007
Dissertação apresentada ao Instituto Oceanográfico da Universidade de São Paulo, como parte dos requisitos para obtenção do título de Mestre em Ciências, área de Oceanografia Biológica
Orientadora: Profa. Dra. June Ferraz Dias
PDF processed with CutePDF evaluation editionwww.CutePDF.com
Influência do ambiente na higidez de larvas de
engraulidídeos coletadas na região de Santos (SP)
Carla Bertolucci Fiadi
Dissertação apresentada ao Instituto Oceanográfico da Universidade de São Paulo, como parte dos requisitos para obtenção do título de Mestre em
Ciências, área de Oceanografia Biológica
Julgada em ___/___/___
_____________________________________ _______________ Prof(a). Dr(a). Conceito
_____________________________________ _______________ Prof(a). Dr(a). Conceito
LISTA DE TABELAS.. ...i
LISTA DE FIGURAS ... ii
AGRADECIMENTOS ... iv
RESUMO ... vii
ABSTRACT...viii
1. INTRODUÇÃO ... 1
1.1. PANORAMA GERAL ... 1
1.2. HIGIDEZ DAS LARVAS DE PEIXES ... 2
1.3. INFLUÊNCIA DO AMBIENTE ... 6
2. 0BJETIVOS... 8
3. MATERIAL E MÉTODOS ... 9
3.1. HIDROGRAFIA... 9
3.2. COLETA DAS LARVAS...11
3.3. TAXONOMIA DAS LARVAS DE ENGRAULIDÍDEOS...12
3.4. PROCESSAMENTO DAS LARVAS ...15
3.5. ANÁLISE DOS DADOS...18
4. RESULTADOS ...21
4.1. HIDROGRAFIA...21
4.2. MORFOMETRIA...24
4.3. CONDIÇÃO APARENTE ...26
4.4. PROTEÍNAS HIDROSSOLÚVEIS ...27
5. DISCUSSÃO ...31
5.1. HIDROGRAFIA...31
5.2. HIGIDEZ DAS LARVAS ...33
5.3. INFLUÊNCIA DO AMBIENTE ...38
6. CONCLUSÕES...43
7. CONSIDERAÇÕES FINAIS ...44
Tabela 1. Valores de taxa de crescimento (mm/dia), de comprimento das larvas (C) (mm), com nota sobre comprimento total (CT) ou padrão (CP), para engraulidídeos, retirados de trabalhos realizados em diferentes regiões.
Tabela 2. Número de exemplares utilizados em cada medida: comprimento padrão (CP), comprimento total (CT), altura da cabeça (AC), altura do corpo na região da nadadeira peitoral (AP), altura do corpo na região do ânus (AA), altura da cabeça/diâmetro do olho (AC/DO), e em cada análise: fator de condição (K), quantificação de proteína e relação proteína/DNA; separados por estação para setembro de 2005.
Tabela 3. Número de exemplares utilizados em cada medida: comprimento padrão (CP), comprimento total (CT), altura da cabeça (AC), altura do corpo na região da nadadeira peitoral (AP), altura do corpo na região do ânus (AA), altura da cabeça/diâmetro do olho (AC/DO) e em cada análise: fator de condição (K), quantificação de proteína e relação proteína/DNA; separados por estação para março de 2006.
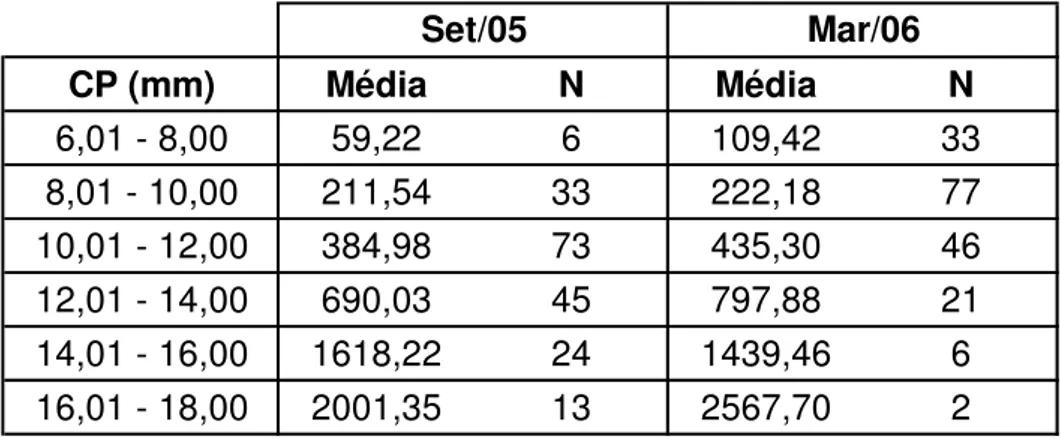
Tabela 4. Valores médios de quantidade de proteína para cada classe de comprimento padrão (CP) e número de exemplares (N) de engraulidídeos coletados na plataforma continental ao largo de Santos, SP.
Tabela 5. Valores médios da relação proteína/DNA para cada classe de comprimento padrão (CP) e número de exemplares (N) de engraulidídeos coletados na plataforma continental ao largo de Santos, SP.
61
62
62
63
Figura 1. Foto indicando a redução na altura da cabeça (AC) de
larvas de Clupea harengus, devido à inanição.
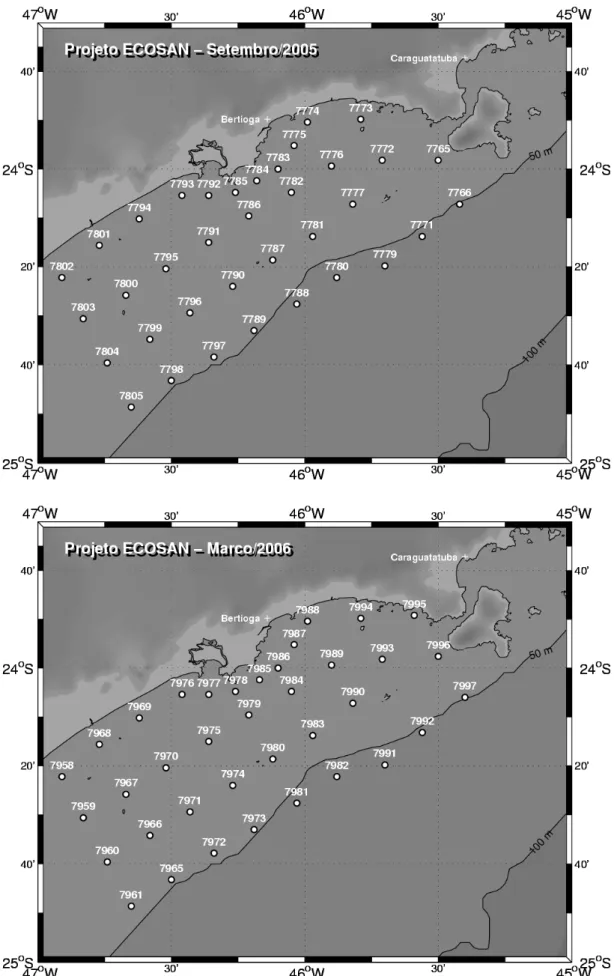
Figura 2. Mapa da região de estudo com as estações amostradas nas duas coletas.
Figura 3. Larva de anchoita com indicação das medidas tomadas – CT (comprimento total), CP (comprimento padrão), DO (diâmetro do olho), AC (altura da cabeça), AP (altura do corpo na região da nadadeira peitoral) e AA (altura do corpo na região do ânus).
Figura 4. Protocolo utilizado na quantificação de DNA.
Figura 5. Protocolo utilizado na quantificação de proteínas.
Figura 6a. Valores de altura da cabeça (AC), altura do corpo na região da nadadeira peitoral (AP) e altura do corpo na região do ânus (AA) plotados em função do comprimento padrão (CP).
Figura 6b. Valores de quantidade de proteína, proteína/DNA e altura da cabeça/diâmetro do olho (AC/DO) plotados em função do comprimento padrão (CP).
Figura 7. Diagramas TS para setembro de 2005 e março de 2006.
Figura 8a. Distribuição horizontal de temperatura e salinidade, a 5m de profundidade e no fundo, para setembro de 2005.
Figura 8b. Distribuição horizontal de temperatura e salinidade, a 5m de profundidade e no fundo, para março de 2006.
Figura 9. Mapa da região de estudo com as radiais utilizadas nas seções verticais.
Figura 10a. Distribuição vertical de temperatura e salinidade, em 3 radiais diferentes, para setembro de 2005.
Figura 10b. Distribuição vertical de temperatura e salinidade, em 3 radiais diferentes, para março de 2006.
Figura 11. Distribuição horizontal de densidade, a 5m de profundidade e no fundo, para setembro de 2005 e março de 2006.
64
65
66
67
68
69
70
71
72
73
74
75
76
Figura 13. Freqüência de larvas em cada classe de comprimento padrão (CP).
Figura 14. Relação entre massa e comprimento padrão (CP).
Figura 15. Valores médios e desvios padrão do fator de condição (K) para cada classe de comprimento padrão (CP).
Figura 16. Valores médios e desvios padrão de altura da cabeça (AC), altura do corpo na região da nadadeira peitoral (AP) e altura do corpo na região do ânus (AA) para cada classe de comprimento padrão (CP).
Figura 17. Freqüência de larvas frágeis em cada classe de comprimento padrão (CP).
Figura 18. Freqüência de larvas frágeis em cada estação.
Figura 19. Mapa evidenciando as estações contendo mais do que 50% de larvas frágeis e sinalizando onde foi encontrada diferença significativa.
Figura 20. Valores médios e desvios padrão de quantidade de proteína para cada classe de comprimento padrão (CP).
Figura 21. Valores médios e desvios padrão de proteína/DNA para cada classe de comprimento padrão (CP).
Figura 22. Freqüência de larvas com valores de quantidade de proteína abaixo da média.
Figura 23. Freqüência de larvas com valores de proteína/DNA abaixo da média.
Figura 24. Mapa evidenciando as estações contendo mais do que 50% de larvas com quantidade de proteína abaixo da média e sinalizando onde foi encontrada diferença significativa.
Figura 25. Mapa evidenciando as estações contendo mais do que 50% de larvas com valores de proteína/DNA abaixo da média e sinalizando onde foi encontrada diferença significativa.
79
80
81
82
83
84
85
86
86
87
88
89
Agradeço à June pela orientação, pelas oportunidades, pela convivência,
pelo ambiente agradável que ajudou e permitiu ser criado no Laboratório de
Ecologia Reprodutiva e do Recrutamento de Organismos Marinhos (vulgo
LERROM), por me salvar e incentivar quando na reta final achei que tudo
estava perdido e que a dissertação não seria entregue. Tenho grande
admiração e carinho por você e sinto falta dos tempos que a gente podia sentar
para abrir peixe ou macerar larva.
A todos que já passaram pelo LERROM agradeço a companhia, os
momentos de aprendizado e de descontração. Em especial, agradeço à Helcy
que sempre me ajudou, desde os primeiros dias de estágio até agora durante o
meu mestrado e que se dispôs diversas vezes a resolver problemas e tornar
tudo mais prático e fácil para que eu pudesse desenvolver meu trabalho
tranqüilamente. Pela valiosa ajuda nos momentos finais de elaboração dessa
dissertação, agradeço à Isa e à Thassya, devo uma pra vocês, hein?! E
agradeço ao Frango por ser o Frango... e por ter me ajudado a estudar pra
passar na prova de ingresso pro mestrado.
À Tati por ter me ensinado tantas coisas sobre os elasmos e por ter sido
tão boa companhia em todos os momentos, se eu tivesse te dado ouvido e feito
o mestrado com os machos de cação-viola, muita dor de cabeça poderia ser
evitada... Saudades de você, amiga.
Quando o computador travava, quando a máquina de xerox pifava,
Sem o auxílio do prof. Daniel Lemos e dos alunos do Laboratório de
Aqüicultura Marinha, a quantificação de proteínas não teria sido realizada,
agradeço a oportunidade oferecida e o investimento de tempo e material para
realização dessa importante etapa do trabalho.
As informações referentes à distribuição de clorofila foram fornecidas
pelo Laboratório de Fitoplâncton e Produção Primária, agradeço à Flávia e à
profa. Sônia Gianesella por terem compartilhado essas informações. Agradeço
também ao Kenji do Laboratório de Produção Secundária por ter
disponibilizado a versão "quase" final de sua monografia.
Ao prof. Paulo Sumida e ao pessoal do Laboratório de Dinâmica Bêntica
pelo fornecimento contínuo de gelo.
Ao prof. Ilson e ao pessoal do LaDO pelas conversas durante as
inúmeras vezes que invadi o laboratório na hora do almoço.
Agradeço ao CNPq pela bolsa concedida.
A todo pessoal da biblioteca, da secretaria da pós e do DOB.
À minha família por sempre ter me apoiado e acreditado em mim. Em
especial à minha mãe que cuidou de mim até eu casar, e ainda cuida quando
tenho que passar uns dias em São Paulo, ao meu pai com quem me pareço
talvez mais do que gostaria, aos meus irmãozinhos (os dois de sangue e a que
eu escolhi pra ser minha irmãzinha) dos quais sinto uma falta enorme quando
não estão por perto, admiro muito vocês, e à minha vó por sempre dar bons
gráficos das análises hidrográficas e estava sempre pronto a responder minhas
dúvidas e a ensinar coisas novas. Foi quem me ajudou nos momentos difíceis,
enxugou minhas lágrimas quando o desespero batia, me acalmou quando o
nervosismo tomou conta, se alegrou com cada nova conquista e sempre me
O principal objetivo desse trabalho foi analisar a influência das condições
ambientais na higidez das larvas de engraulidídeos da plataforma continental
ao largo de Santos (24o49’S – 23o49’S e 46o55’W – 45o24’W). As coletas foram
realizadas em dois cruzeiros oceanográficos (setembro de 2005 e março de
2006). Verificou-se diferença nas condições hidrográficas entre as duas
campanhas de coletas, sendo que em setembro de 2005 houve maior
influência de descarga de águas estuarinas na plataforma, enquanto em março
de 2006 houve intrusão da Água Central do Atlântico Sul. As duas situações
geraram estratificação vertical e favoreceram a entrada de nutrientes no
sistema, o que gera enriquecimento da zona eufótica e beneficia as cadeias
tróficas locais. Para avaliar a condição de higidez das larvas, foram utilizados
indicadores morfológicos (relação massa-comprimento, fator de condição,
altura do corpo da larva, relação entre altura da cabeça e diâmetro do olho) e
bioquímicos (quantidade de proteína e relação proteína/DNA). Os indicadores
apontaram melhor condição relativa, para as larvas coletadas em março de
2006, que pode ser conseqüência da ocorrência de uma intensa estratificação
vertical na coluna de água e de uma maior estabilidade gravitacional em março
de 2006, proporcionando maior concentração de nutrientes e ocasionando
aumento na densidade e melhora na condição das populações planctônicas.
Palavras-chave: engraulidídeos, larvas de peixes, higidez, plataforma
The aim of this study was to analyze influence of environmental conditions on
health of engraulidid larvae of the continental shelf off Santos (24º49´S –
23º49´S e 46º55´W – 45º24´W). Samples were collected during two
oceanographic cruises (September 2005 and March 2006). Differences among
hydrographic conditions were verified between surveys: in September there was
a major influence of the estuarine water discharge in the inner shelf, whereas in
March there was an intrusion of the South Atlantic Central Water (SACW) in the
region. The two conditions generated vertical stratification and improved the
pelagic food web, in which fish larvae participate. Morphological (weight-length
relation, condition factor, larvae body height, head height-eye diameter relation)
and biochemical (protein quantity and protein/DNA relation) indicators were
used to evaluate the larvae health condition. Results showed better relative
condition of larvae collected in March 2006 that can be a consequence of the
intense water column vertical stratification and gravitational stability in March
2006, providing higher nutrients concentration, increasing density of organisms
and improving condition of planktonic populations.
1. INTRODUÇÃO
1.1. PANORAMA GERAL
A região sudeste ocupa lugar de destaque na produção pesqueira
nacional, sendo que, a região da baixada santista apresenta-se como um dos
ecossistemas marinhos mais produtivos da costa brasileira. O desembarque
pesqueiro tem sido reduzido, porém ainda bastante expressivo, e a atividade
pesqueira artesanal apresenta importante papel sócio-econômico (Bertozzi,
2002).
A região também apresenta indiscutível importância no que diz respeito
ao comércio externo do país. Entre o final de 2002 e o final de 2006, o Porto de
Santos foi responsável por 27% do que foi comercializado pelo Brasil
(exportação e importação) (Porto de Santos, 2007).
A pressão pesqueira, a movimentação de embarcações devido ao porto
e a acelerada urbanização que vem ocorrendo na área, podem influenciar o
ecossistema marinho, alterar características fisiológicas dos organismos e
induzir modificações ecológicas das populações e comunidades que habitam a
região.
A complexidade do ambiente em questão mostra-se ainda maior pela
presença do estuário de Santos. Entretanto, apesar da importância e
complexidade da região, poucos estudos foram realizados no que concerne
aos aspectos ecológicos do ambiente marinho.
O projeto multidisciplinar intitulado “A influência do complexo estuarino
visa ampliar o conhecimento a respeito do estuário, da plataforma continental e
da interação entre esses dois ambientes. Dentro desse projeto, uma das
preocupações foi estudar alguns aspectos ecológicos da ictiofauna da região.
A dinâmica populacional dos peixes é influenciada direta e indiretamente
pela ação antrópica, porém as alterações causadas pela crescente urbanização
e exploração dos recursos marinhos não são as únicas responsáveis pelas
flutuações no estoque, nos quais ocorre uma variação populacional natural que
necessita de maior compreensão. O entendimento dessas flutuações passa
pelo entendimento da dinâmica larval das espécies de teleósteos. Falhas no
recrutamento, ou seja, no processo de entrada de uma classe etária na vida
juvenil ou adulta ou em um estoque passível de ser pescado, irão afetar a
captura anual na maioria dos recursos pesqueiros (Rothschild, 1986; Kimura et
al., 2000).
1.2. HIGIDEZ DAS LARVAS DE PEIXES
Lasker (1987) afirma que os estudos voltados ao início do ciclo de vida
dos peixes não têm sua importância limitada ao ambiente acadêmico, mas se
estendem à sociedade em geral por serem relevantes para as atividades de
pesca e aqüicultura, pois se sabe que as maiores taxas de mortalidade
ocorrem no período larval. Essa acentuada mortalidade está ligada a dois
principais fenômenos que ocorrem durante a fase larval: predação e inanição.
A alimentação desfavorável, assim como fatores hidrográficos e
patologias podem interferir no crescimento das larvas, e baixas taxas de
crescimento podem prolongar a duração dos estágios larvais, aumentando o
disso, Sclafani et al. (1997) realizaram experimentos indicando que larvas em
inanição apresentam capacidade de osmorregulação prejudicada aumentando
sua densidade em meio hipertônico, o que facilita seu afundamento; esse
processo pode transportar as larvas para águas ainda menos propícias ao seu
desenvolvimento.
Desse modo, estudos da condição nutricional e das taxas de
crescimento e mortalidade das larvas vêm sendo desenvolvidos com o objetivo
de elucidar os mecanismos que regem o recrutamento.
Para o estudo da condição nutricional de larvas são utilizados métodos
que podem ser agrupados em três principais categorias:
• Morfometria – análise de relações entre determinadas medidas
tomadas das larvas;
• Histologia – análise de preparações com cortes de larvas, buscando
verificar possíveis alterações em estruturas dos tecidos que podem
ser afetados pela inanição;
• Bioquímica – análise, principalmente, de lipídios, proteínas, atividade
tripsolítica e relação RNA-DNA
Ehrlich et al. (1976) estudou mudanças morfológicas e histológicas
durante o crescimento e inanição em Clupea harengus (arenque) e
Pleuronectes platessa (solha). Nesse trabalho, entre outros resultados, foi
verificado que a condição de inanição acarretou a redução no tamanho das
células epiteliais do trato digestório das duas espécies; e em C. harengus
houve encolhimento da região da cabeça evidenciado pelo aumento da relação
Para Engraulis mordax, O’Connel (1976) analisou, com base na
histologia, alterações na estrutura dos tecidos, elaborando uma escala para
classificar cada uma das alterações. Esse método permitiu bons resultados
para separação das larvas bem nutridas daquelas em inanição, através da
análise multivariada.
Theilacker (1978) focou seu estudo em modificações de características
morfológicas e histológicas devido à inanição, utilizando larvas de Trachurus
symmetricus. Foi encontrada diferença entre larvas bem nutridas e em inanição
na estrutura do cérebro, fígado, pâncreas, trato digestório e musculatura.
Quanto à morfometria, foi constatado que não existe uma medida que, sozinha,
possa ser usada para separar larvas em inanição das demais; porém, ao
analisar relações entre as medidas fica possível realizar tal agrupamento. A
autora também afirma que critérios morfológicos são mais práticos de serem
utilizados, pois requerem menos tempo e não necessitam de técnicas especiais
de preservação, porém, é ressaltado que análises histológicas podem ser mais
acuradas para classificação individual das larvas. Além disso, quando uma
larva sofre privação alimentar ocorrem processos caracterizados por alterações
importantes, do ponto de vista da observação celular e tecidual, ao longo do
tempo, que podem culminar com a necrose celular. A partir do momento em
que os processos irreversíveis se iniciam, atinge-se o chamado ponto de
não-retorno (“point of no return”), em que não há mais possibilidades de
recuperação, sendo que somente o acompanhamento histológico identifica
indubitavelmente a inanição ( .
Powell & Chester (1985) também utilizaram métodos morfométricos
utilizaram métodos histológicos para analisar a condição nutricional de larvas
do gênero Engraulis, coletadas no ambiente.
Além desses estudos sobre condição nutricional, algumas
características morfológicas da larva, podem indicar seu estado de higidez
como danos encontrados na musculatura e trato digestório, aspecto e
coloração da musculatura e/ou cabeça, indicando fragilidade (Dias et al., 2004).
A condição de saúde da larva é crucial para seu desenvolvimento, mas a
utilização de uma abordagem ecológica no estudo dessa condição para uma
determinada população ou comunidade pode avaliar épocas e/ou regiões
favoráveis para desenvolvimento mais adequado das larvas, bem como,
fornecer dados importantes para a estimativa do recrutamento das espécies
estudadas.
A abordagem ecológica, que visa entender as relações entre os seres
vivos e entre estes e o ambiente em que vivem, exige, no caso de organismos
planctônicos marinhos, um entendimento das condições hidrográficas da região
de estudo. Esse fato se deve à própria natureza do plâncton, isto é, sua
capacidade limitada de locomoção, fazendo com que esses organismos sejam
levados pelas correntes existentes no oceano. Desse modo, o ictioplâncton
está sujeito às condições ambientais nas quais se encontra, sendo incapaz de
deslocamentos de maior escala em busca de alimento.
O estudo da hidrografia também auxilia no entendimento dos fenômenos
que podem gerar um acréscimo ou decréscimo de nutrientes disponíveis,
facilitando a compreensão das condições existentes na região para a época
1.3. INFLUÊNCIA DO AMBIENTE
As características hidrográficas interferem direta e indiretamente nos
fatores bióticos, de forma que, o entendimento de diversos aspectos da
ecologia de organismos marinhos passa pela análise de dados abióticos.
A Plataforma Continental Sudeste (PCSE) apresenta massas de água
que, geralmente, são resultado da mistura entre a Água Tropical, quente e
salina (T > 20oC e S > 36,4), Água Central do Atlântico Sul, mais fria (T < 20oC
e S < 36,4) e Água Costeira, menos salina (S < 34): a Água Tropical é
transportada para o sul-sudoeste na camada superficial da Corrente do Brasil
(CB) próximo à quebra de plataforma; a Água Central do Atlântico Sul é
transportada na mesma direção, porém, na camada inferior da CB, enquanto a
Água Costeira é encontrada na região mais próxima à costa e resulta da
mistura da descarga continental de águas interiores com as águas da
plataforma continental (Castro et al. 2006).
Quanto à produtividade primária das águas citadas, Gaeta & Brandini
(2006) afirmam que, no geral, a Água Costeira permanece mais produtiva
durante todo o ano devido ao aporte de nutrientes de origem continental, mas a
Água Tropical possui baixa produtividade em função da escassez de nutrientes
e a Água Central do Atlântico Sul, apesar da abundância de nutrientes,
encontra-se abaixo da zona eufótica. Os autores ainda esclarecem que
fenômenos como ressurgência, ondas internas e vórtices ciclônicos podem
ocasionar a fertilização da zona eufótica com nutrientes provindos da Água
Central do Atlântico Sul.
Considerando as afirmações acima, podemos identificar duas
fluvial, que traz material oriundo do continente para o oceano, enriquecendo as
regiões próximas à desembocadura do rio; e a ressurgência costeira, quando
águas mais profundas e ricas em nutrientes afloram na região próxima à costa
ou na zona eufótica, podendo ocasionar volumoso aumento na produção
primária marinha. Esses dois processos podem ocorrer em uma mesma região,
coexistindo ou não, temporalmente.
Os fatores que proporcionam um enriquecimento da coluna d'água, além
daqueles que permitem uma maior probabilidade de encontro da larva com sua
presa, costumam ser determinantes no recrutamento de teleósteos. Allain et al.
(2001), utilizando um modelo hidrodinâmico 3D, verificaram a influência da
ressurgência costeira, da estratificação vertical e a extensão da pluma fluvial no
recrutamento de Engraulis encrasicolus na Baía de Biscaia, demonstrando que
os dois primeiros processos atuam mais fortemente nas variações do
recrutamento da espécie em questão.
Já no trabalho de Vasconcellos et al. (1998), realizado na costa sul do
Brasil, os autores encontraram uma situação de maior sucesso de alimentação,
para larvas de Engraulis anchoita, no inverno e acreditam que tal fato se deve à
contribuição das águas vindas do continente e da presença de águas frias de
origem subantártica na região. Novamente o efeito da estratificação vertical da
coluna d'água é apontado como componente importante para o
2. 0BJETIVOS
O objetivo central deste trabalho é analisar a influência do ambiente na
higidez das larvas de engraulídideos da plataforma continental adjacente ao
estuário de Santos, além de verificar a ocorrência de variação espacial e/ou
temporal nas larvas com diferentes situações de condição.
Os objetivos específicos são:
• Inferir a condição de higidez das larvas através de características
morfológicas e de quantificação de proteínas;
• Analisar as condições hidrográficas através de seções e de
distribuições horizontais de temperatura, salinidade e densidade;
• Avaliar a variação espacial dos resultados obtidos, através de
possíveis diferenças entre as estações de uma mesma coleta;
• Avaliar a variação temporal dos resultados obtidos, através de
possíveis diferenças entre as coletas;
Hipótese de trabalho: Larvas de engraulidídeos estão em melhor condição
relativa em situações hidrográficas de maior estabilidade gravitacional e
3. MATERIAL E MÉTODOS
As coletas foram realizadas a bordo do N/Oc. Prof. W. Besnard em duas
campanhas de coleta: setembro de 2005 (set/05) e março de 2006 (mar/06). A
duração dos cruzeiros foi oito dias na primeira campanha (19 a 26/09/05) e
cinco dias na segunda campanha (12 a 16/03/06), na região entre 24°49’S e
23°49’S e 46°55’W e 45°24’W. Um mapa da região de estudo contendo as
estações de coleta pode ser visto na figura 2. O presente estudo faz parte do
ECOSAN (“A influência do complexo estuarino da baixada santista na região da
plataforma continental adjacente”), projeto integrado, multi- e interdisciplinar,
sobre a influência, remota e local, do continente e da baía/estuário de Santos
na plataforma continental adjacente à baixada santista. Dentro do projeto, as
diferentes áreas da Oceanografia vêm desenvolvendo estudos, que objetivam
um maior entendimento da dinâmica do ecossistema costeiro da região.
3.1. HIDROGRAFIA
Para obtenção dos perfis verticais de temperatura e salinidade, foi
utilizado um CTD (conductivity, temperature and depth sonde), sendo que a
salinidade é obtida através dos dados de condutividade.
O tratamento dos dados obtidos foi feito em duas etapas:
1) remoção de picos (“spikes”) – realizada utilizando-se o software de
pós-processamento fornecido pelo fabricante do CTD (FSI – Falmouth Scientific,
INC.1). Esse processo remove os valores discrepantes, geralmente originados
por falha de comunicação do equipamento, através da comparação com
intervalos pré-estabelecidos de valores aceitáveis para temperatura e
condutividade para o perfil como um todo e a cada 30 m de profundidade.
2) média em caixas (“bin averaging”) – após a remoção dos picos, foi feita a
promediação vertical dos dados em intervalos equi-espaçados de
profundidade; a freqüência de amostragem adotada foi de 4 Hz e, como o
intervalo de profundidade foi estabelecido a cada 1 m, há no mínimo quatro
valores por caixa de profundidade, já que a velocidade de descida do CTD,
durante o perfilamento, foi de aproximadamente 1 m s-1.
Ao final desse tratamento, os perfis verticais apresentam-se alisados e
os valores de temperatura e salinidade equi-espaçados verticalmente.
Para obtenção das seções verticais de temperatura, salinidade e
densidade, foi elaborada uma grade bi-dimensional na qual os dados foram
interpolados linearmente. Esse mesmo procedimento foi adotado na
elaboração das distribuições horizontais (Emery & Thompson, 2001).
O método linear de interpolação foi escolhido por sua simplicidade, uma
vez que assume como sendo linear a variação espacial da propriedade
amostrada. Em função desta premissa, podem ocorrer descontinuidades no
campo interpolado. A fim de atenuá-las, mas preservando os padrões das
feições presentes, foi aplicado pontualmente um filtro espacial gaussiano do
3.2. COLETA DAS LARVAS
A coleta do ictioplâncton foi realizada com rede bongô operando com
duas malhas: 500 e 300 µm, seguindo a metodologia descrita em Smith &
Richardson (1977). O tempo total de arrasto variou de acordo com a
profundidade local do ponto amostrado, sendo realizada uma amostragem
oblíqua, de maneira que, a rede descia até a profundidade máxima
estabelecida (a 5 m da superfície de fundo ou no limite de 200 m), e subia após
30 segundos de arrasto na profundidade máxima. Se a profundidade local era
menor que 60 m, a rede subia e descia duas vezes sem sair da água para
aumentar o volume de água filtrado. Como a rede opera aberta durante todo o
processo, em estações mais profundas, um maior fluxo de água é filtrado,
durante a descida e subida.
Como a coleta é oblíqua e a rede não apresenta dispositivos de
fechamento, toda a coluna de água é amostrada, sem a possibilidade de
estudar a distribuição vertical dos organismos coletados.
Quando o produto da coleta da rede fina apresentava um mínimo de 5
larvas visíveis, todo o material coletado pelas duas redes era imediatamente
colocado no gelo e levado ao laboratório de bordo para triagem. Para evitar
autólise, as larvas foram triadas em, no máximo, 15 minutos e imediatamente
acondicionadas em tubos criogênicos, que foram depositados em nitrogênio
3.3. TAXONOMIA DAS LARVAS DE ENGRAULIDÍDEOS
As amostras foram levadas ao laboratório na sede do Instituto
Oceanográfico, descongeladas, mas mantidas em gelo e, com auxílio de
estereomicroscópio, foi feita a identificação das larvas em nível de família.
De acordo com Katsuragawa et al. (2006), são reconhecidas doze
espécies de engraulidídeos na costa sudeste do Brasil e, excetuando-se
Engraulis anchoita que apresenta distribuição mais ao largo (Nakatani, 1982;
Asano et al., 1991; Bakun & Parrish, 1991; Matsuura et al., 1991), os
engraulídideos possuem hábito costeiro, com ocorrência em águas de baixa
salinidade. Para a costa sudeste e sul do Brasil, excetuando-se a anchoíta que
vem sendo exaustivamente estudada, larvas de engraulidídeos costeiros foram
pouco estudadas sob diferentes aspectos: Cetengraulis edentulus sobre
estágios iniciais (Kraus & Bonecker, 1994); Anchoa tricolor sobre morfologia do
desenvolvimento (Koblitz, 1990; Pinheiro et al., 1996); A. tricolor e Anchoa parva
sobre caracteres-diagnóstico e épocas de desova (Godefroid et al., 1999;
Hofstaetter et al., 2002; Hofstaetter et al., 2004). Trabalhos costeiros sobre
ictioplâncton indicam a predominância das larvas de E. anchoita em estações
oceanográficas mais ao largo, mas mantêm outros engraulidídeos no nível de
Família (Katsuragawa et al., 1993; Katsuragawa & Dias, 1997).
Embora a identificação das larvas tenha sido baseada em referências
clássicas, como Moser et al. (1984) e Leis & Trinsk (1989), bem como recentes
(Farooqi et al., 2006), e em caracteres como contagem de miômeros ou
vértebras, número de raios de nadadeiras, padrões de pigmentação ou posição
relativa das nadadeiras, houve grande sobreposição dos valores dos
nadadeiras, tornando baixa a resolução taxonômica desses caracteres para a
identificação das larvas coletadas. Assim, não foi possível a identificação de
gêneros ou espécies dos engraulidídeos que ocorreram na região de estudo.
Na região de Santos, que inclui baía, costa, praias e estuários, está
relatada a ocorrência de nove espécies: Anchoa filifera, Anchoa januaria,
Anchoa marinii, Anchoa spinifera, Anchoa tricolor, Anchovia clupeoides,
Anchoviella brevirostris, Anchoviella lepidentostole, Lycengraulis grossidens
(Paiva Filho, 1982; Paiva Filho et al., 1987; Ribeiro Neto, 1989; Dias, 2007b).
Este fato dificulta ainda mais uma tomada de decisão sobre a taxonomia larval
com base nos caracteres listados acima.
Com o intuito de verificar se havia semelhanças ecológicas que não se
restringissem apenas à guilda trófica entre as larvas da família, foram
levantados dados na literatura sobre taxas de crescimento das larvas de
Engraulididae, provenientes de diferentes locais e de faixas de comprimento
diversas, para que se pudesse trabalhar com maior confiabilidade no nível de
Família, uma vez que os indicadores de condição ligados ao metabolismo
(quantidade de proteínas e de DNA) pudessem ser comparáveis em nível
ecológico e não apenas taxonômico. A preocupação foi não gerar erros de
interpretação na análise da condição larval, uma vez que valores muito
discrepantes de taxas de crescimento para larvas de diferentes espécies
sugeririam que essas diferenças não estariam relacionadas à condição
nutricional do grupo estudado, mas à estratégia de vida e diferenças
intra-específicas no desenvolvimento das diferentes espécies.
Na tabela 1 são encontrados valores de taxas de crescimento de larvas
deixa claro que a taxa de crescimento varia de acordo com o local e/ou
condições às quais as larvas foram submetidas, mais do que variam de espécie
para espécie. Este fato pode ser explicado pela dependência que a taxa de
crescimento tem de uma série de fatores ligados às condições ambientais a
que essas larvas são submetidas, bem como da fase de desenvolvimento em
que estas se encontram (Suthers, 1998).
Como conseqüência, diferentemente de estudos desenvolvidos com
larvas de Engraulis anchoita, espécie que ocorre apenas mais ao largo, não foi
possível identificar as larvas coletadas nos cruzeiros oceanográficos a serem
usados neste estudo. Porém, julgou-se plenamente possível usar uma
abordagem ecológica para as larvas da Família, uma vez que a redução da
variabilidade intraespecífica na condição larval é difícil de se obter, mesmo em
laboratório, pois não há razões para se esperar que os indivíduos, mesmo que
de mesma espécie, respondam de maneira similar à disponibilidade de
alimento (Ferron & Leggett, 1994). Além disso, Ramírez et al. (2001) discutem
que, para alguns clupeóides, a morfologia larval muda acentuadamente apenas
quando estas atingem a fase de metamorfose e transição para a fase juvenil.
Além dos engraulidídeos, foram capturados, somente em março de
2006, alguns clupeídeos, cuja identificação é mais rotineira (Sardinella
brasiliensis e Harengula jaguana), porém em número muito baixo, o que
inviabilizou a abrangência do estudo a outros pequenos pelágicos. Desta
forma, ficou decidido que os esforços deveriam concentrar-se apenas nos
3.4. PROCESSAMENTO DAS LARVAS
Após a identificação das larvas, com auxílio de estereomicroscópio e
uma escala micrometrada acoplada, foi realizada a tomada dos seguintes
dados das larvas, ilustrados pela figura 3:
- comprimento total (CT) - da ponta do focinho até o final da nadadeira caudal
ou da membrana embrionária;
- comprimento padrão (CP) - da ponta do focinho até a extremidade da
notocorda; - altura da cabeça na região nucal (AC) – da nuca até a região
ventral;
- altura do corpo na região da nadadeira peitoral (AP) – do dorso até o ventre;
- altura do corpo na região do ânus (AA) – do dorso até o ventre, excluindo-se o
trato digestório, cujas dimensões podem variar em função da presença ou não
de bolo alimentar;
- diâmetro do olho (DO) – equivalente à medida vertical de uma borda à outra,
passando-se pelo centro (não foi medido o tamanho da órbita).
Foram escolhidos dois tipos de indicadores de condição, com diferentes
níveis de resposta à origem e disponibilidade de alimento, bem como a fatores
abióticos: indicadores organismais morfológicos, de resposta mais lenta, e
indicadores bioquímicos, que respondem mais rapidamente a agentes
estressores ambientais.
Indicadores organismais morfológicos
Foram selecionados os seguintes indicadores: relação
massa-comprimento, altura da cabeça, relação entre altura da cabeça e diâmetro do
região do ânus, além da condição aparente das larvas e do fator de condição
de Foulton. Foram também estimados os comprimentos totais de larvas que
apresentavam danos na nadadeira caudal, a partir da equação comprimento
padrão-comprimento total.
A relação entre altura do corpo e a da cabeça têm sido consideradas
boas discriminantes das larvas em condições precárias (Ehrlich et al., 1976;
Theilacker, 1978, 1986; Powell & Chester, 1985).
A condição aparente das larvas foi estimada a partir de observações
objetivas a respeito do estado de higidez. Foram considerados: 1) presença de
danos não provenientes de manipulação ou congelamento; 2) aspecto da
musculatura (coesão dos miômeros e coloração); 3) aspecto da cabeça
(formato, integridade da mandíbula, presença dos olhos); 4) integridade e
ligação do trato digestório com o corpo. Algumas dessas características foram
usadas para separar larvas coletadas vivas daquelas que já estavam mortas
antes da coleta (Dias et al., 2004). Quando possível, foi medida a massa das
larvas. Larvas consideradas demasiadamente frágeis ou com problemas devido
ao congelamento não tiveram suas massas medidas, para evitar dano ainda
maior em suas estruturas.
Como algumas medidas dependiam de manipulação e mudança de
posição das larvas sobre a placa de Petri, nem sempre era possível alocar
essas larvas da maneira correta e, como conseqüência, o número de larvas
das quais foram tomadas as medidas de AC, AP, AA e DO variou e foi sempre
consideravelmente inferior ao total de larvas amostradas.
A partir de massa úmida e do comprimento total estimado, foi calculado
oceanográficos. O valor de K pode expressar variações no estado fisiológico
dos peixes (Vazzoler, 1996) e foi utilizado por Neilson et al. (1986) para
descrever a condição de larvas de bacalhau do Atlântico. O fator de condição
de Foulton K é expresso pela relação:
K = (Mt / CT
3)10
4em que: Mt é a massa úmida total (g)
CT = comprimento total (mm).
Como o número de larvas das quais foi possível medir o CT foi bastante
reduzido, foi necessário estimar os valores através de um ajuste, a equação
utilizada para descrever a relação entre comprimento padrão e comprimento
total é CT = 1,1763 CP - 0,6434 (R2 = 0,9956), e foi usada para a conversão de
um comprimento pelo outro, para o cálculo do K. Essa equação foi obtida
através da relação entre CT e CP das larvas cujas duas medidas puderam ser
tomadas.
Indicadores bioquímicos
Após a tomada de dados, cada larva foi processada, individualmente,
para quantificação de DNA seguindo protocolo baseado em Caldarone et al.
(2001) com modificações (Fig. 4).
A quantificação de proteínas foi realizada, seguindo protocolo
apresentado na figura 5. A metodologia utilizada é baseada no método de
Bradford (Bradford, 1976) e permite a quantificação de proteínas hidrossolúveis
em microquantidades. Essa quantificação se dá através da utilização de
alterada de marrom para azul e seu máximo de absorbância passa de 465 nm
para 595 nm. As soluções padrões utilizadas foram obtidas através da diluição
de albumina bovina, nas seguintes concentrações: 25, 125, 250, 500, 750,
1000, 1500 e 2000 g/ml. Para cada leitura realizada, uma série dessas
diluições foi utilizada, obtendo-se uma curva de calibração seguindo o padrão:
Y = (A – D) / (1 + (X/C)B) + D
Um exemplo dos valores envolvidos foi retirado de uma das leituras: A =
0,479; B = 1,342; C = 2250,214; D = 2,849; com R2 = 0,949.
A quantidade absoluta de proteínas, bem como base para estimativas da
taxa de crescimento instantâneo, tem sido utilizada como medida da condição
larval, com aplicabilidade em trabalhos sobre cultivo de larvas em laboratório
(Fukuda et al., 2001; Kono et al., 2003; Shoji & Tanaka, 2004), bem como em
trabalhos de campo (Ramírez et al., 2001).
3.5. ANÁLISE DOS DADOS
Uma vez que já foi reportado encolhimento da cabeça em casos de
inanição (Ehrlich 1976), bem como diminuições em medidas que indicam a
altura das larvas (Powell & Chester, 1985), os valores de AC, AP e AA foram
utilizados como indicativos de condição de higidez. De maneira geral, índices
morfométricos são úteis para comparar condição larval entre estações do ano
ou anos, sendo menos sensível para eventos de curta duração (dias, semana)
(Suthers, 1998).
Para avaliar corretamente os dados obtidos, alguns cuidados foram
tomados. As variáveis que apresentaram ligação clara com comprimento
CP. Foi o caso de altura da cabeça (AC), altura do corpo na região da
nadadeira peitoral (AP), altura do corpo na região do ânus (AA) (Fig. 6a),
quantidade de proteína e proteína/DNA (Fig. 6b).
A relação entre AC e diâmetro do olho (DO), que foi obtida dividindo-se o
valor de AC pelo valor de DO para cada larva, não apresentou relação clara
com CP (Fig. 6b), de modo que, foram comparados os valores médios de cada
época de coleta, sem a necessidade de análises por classes de comprimento.
A quantidade de proteínas totais também foi utilizada, pois índices como
os de estocagem (quantidade de carbohidratos, proteínas ou lipídios) podem
revelar respostas a alterações na dieta ou mudanças de habitat (Suthers,
1998). Além disso, ao contrário de outros vertebrados, peixes ou larvas de
peixes com déficit alimentar têm seu metabolismo reduzido e não só produzem
menos proteínas como podem degradar algumas que já possuem, o que
interfere na massa e na condição dos indivíduos (Ferron & Leggett, 1994).
Uma indicação ainda mais precisa é a relação proteína/DNA, uma vez
que a quantidade de DNA, por ser fixa em cada célula, pode representar a
quantidade de células no organismo, assim a relação proteína/DNA é um
indicativo do metabolismo celular.
Para avaliar as diferenças dos valores de quantidade de proteína e
proteína/DNA, entre as estações e entre cruzeiros oceanográficos, foi adotada
a seguinte estratégia: foram empregadas classes de CP com intervalos de 1
mm, sendo necessário juntar os dados das duas coletas, obtendo-se médias
mais consistentes para cada classe de CP; o valor de cada larva foi comparado
ao da média referente à sua classe de CP e foram anotadas as larvas que
estação, a freqüência absoluta e relativa de larvas com valores abaixo da
média da quantidade de proteína ou de proteína/DNA para sua classe de CP.
Para avaliar as diferenças encontradas entre as estações de coleta, foi
utilizado o
χ
2 com a correção de Yates:
em que: Oi = freqüência observada
Ei = freqüência esperada.
4. RESULTADOS
4.1. HIDROGRAFIA
As principais características da distribuição espacial da temperatura e
da salinidade, observadas durante as campanhas de coleta, podem fornecer
dados adicionais sobre a influência dessas propriedades físicas da água do
mar sobre os fenômenos ecológicos.
Nos diagramas TS (Fig. 7) observa-se diferença entre as duas coletas,
sendo que em setembro de 2005 existe maior variação na salinidade, enquanto
em março de 2006 existe maior variação na temperatura. Os valores mínimos e
máximos de salinidade foram 30,78 e 36,29 para setembro de 2005 e, para
março de 2006 foram 33,89 e 36,26. Já a temperatura variou entre 18,96 oC e
21,64oC em setembro de 2005 e entre 16,19 oC e 28,06 oC em março de 2006.
De maneira geral, os diagramas sugerem a presença de águas de baixa
salinidade nos dois períodos, o que já seria esperado em região costeira
especialmente, por sofrer influência do estuário de Santos, que contribui para a
manutenção de águas menos salinas. Porém, em março de 2006, pode-se
observar a nítida presença de águas mais frias na região; além disso, a
salinidade variou menos, sugerindo menor aporte de águas interiores.
Ainda com relação aos diagramas TS, cabe destacar a presença mais
marcante da Água Central do Atlântico Sul em março, identificada pelos valores
de temperatura < 19 ºC e salinidade < 36,4; essa massa d'água praticamente
não se faz presente na região em setembro. A presença da Água Costeira, que
é marcante na região, identificada pelos valores de salinidade abaixo de 34.
Em nenhum dos dois cruzeiros, podemos notar águas com salinidade > 36,4
como é seguramente característico da Água Tropical.
Para que se pudesse entender a diferença apresentada entre as duas
épocas de coleta, foram analisadas as distribuições horizontais de temperatura
e salinidade para 5 m de profundidade e para o fundo (Fig. 8). Constatam-se
duas situações hidrográficas distintas entre as duas épocas. Em setembro de
2005, na ocasião do cruzeiro, fica evidente a existência de uma maior descarga
de água menos salina na plataforma continental interna, procedente da região
do estuário. Ao contrário, em março de 2006 esta contribuição do estuário não
ocorre, mas há a entrada de água mais fria pelo fundo, proveniente da porção
mais externa da plataforma continental.
Nas distribuições horizontais, para setembro de 2005, ainda é possível
notar maior variação entre os valores de salinidade encontrados em 5 m de
profundidade e os valores do fundo, o que não ocorre com a temperatura. Já
em março de 2006 ocorre a situação inversa, ou seja, uma grande variação
entre os valores da temperatura entre os 5 m e o fundo, mas pouca variação no
que diz respeito à salinidade.
Foram realizadas ainda, seções verticais de temperatura e salinidade em
três radiais, sendo a radial 1 mais ao norte, a radial 2 mais próxima ao estuário
e a radial 3 mais ao sul (Fig. 9). Nessas seções, fica mais evidente a ocorrência
do gradiente de salinidade em setembro de 2005, mais intenso nas duas
primeiras radiais e menos intenso na radial 3 (mais ao sul), sugerindo um
aporte de água proveniente do estuário. Além disso, é possível notar a
menos salinas e menos densas entrando na plataforma pelas camadas
superiores da coluna de água, confirmando a origem estuarina desse aporte de
água.
Em situação diferente, na coleta de março de 2006, se destaca o
acentuado gradiente de temperatura, mais intenso na radial 1 (mais ao norte),
que decresce à medida que analisamos as radiais mais ao sul. Apesar de não
haver um gradiente marcante de salinidade em março de 2006 é possível
observar uma contribuição de água menos salina vinda da região mais costeira
na radial 2 (Fig. 10b).
Assim, é possível afirmar que existe estratificação vertical nas duas
épocas amostradas, porém com diferenças quanto a sua origem: em setembro
de 2005 há estratificação em função da salinidade, e em março de 2006 em
função da temperatura. Para avaliar se as situações apresentadas refletiram
em uma estratificação mais ou menos intensa em relação à densidade na
coluna d'água, foram realizadas distribuições horizontais e seções verticais de
densidade. A densidade aqui representada é o valor da densidade total,
subtraída a máxima densidade da água pura (1000 kg/m3)
É possível notar uma maior variação na densidade superficial (5 m de
profundidade) em setembro de 2005 (Fig. 11), porém é em março de 2006 que
ocorre uma maior variação no fundo, e uma maior diferença entre os valores de
5 m de profundidade e do fundo. Analisando as seções verticais observa-se
estratificação nas duas épocas, porém, mais intensa em março de 2006 (Fig.
12). Dessa forma, pode-se concluir que tanto o efeito da salinidade (em
distribuição da densidade, ocasionando situações de estratificação na coluna
d'água.
É sabido que, tanto o aporte de água oriunda do continente quanto a
penetração de águas frias pelas camadas inferiores da coluna de água
ocasionam o enriquecimento dessas camadas. Como mostrado, essas
situações hidrográficas também podem proporcionar a manutenção de uma
coluna de água estável e estratificada, sendo que o conjunto dos dois
fenômenos pode propiciar melhores condições ecológicas para as larvas de
peixes, bem como para todo o zooplâncton em geral.
4.2. MORFOMETRIA
Em setembro de 2005 foram avaliadas 216 larvas, das quais apenas 51
puderam ser medidas quanto à massa. Já em março, das 202 larvas
analisadas, 120 tiveram suas massas medidas. As tabelas 2 e 3 apresentam o
número de larvas utilizadas neste estudo por estação hidrográfica e por
cruzeiro.
A distribuição de freqüência de larvas em cada classe de comprimento
padrão (CP) pode ser vista na figura 13. Houve maior freqüência de
exemplares na classe 10,01 - 12,00 mm em setembro de 2005 e na classe 8,01
– 10,00 mm em março de 2006. Nas maiores classes de CP (16,01 – 18,00 mm
e >18,00 mm) existem poucos representantes em março de 2006. É possível
verificar, através do deslocamento da moda em direção aos valores mais
baixos de CP, que em março de 2006 foram coletados indivíduos menores do
A relação entre massa e CP (Fig. 14) mostra uma maior inclinação da
curva ajustada em setembro de 2005, indicando um maior incremento em
massa para um mesmo valor de CP, porém, o número de exemplares utilizados
nessa análise é bastante reduzido devido à dificuldade já relatada
anteriormente. Além disso, a nuvem de pontos concentrada nos comprimentos
entre 5 e 10 mm para março de 2006 força a menor inclinação, deixando
dúvidas sobre uma comparação direta desses resultados.
Também foi calculado o fator de condição (K) das larvas, sendo que a
média e desvio padrão para cada classe de CP podem ser vistos na figura 15.
Nas menores classes (6,01 - 8,00 e 8,01 - 10,00 mm) não houve larvas com
massa medida em setembro de 2005, mas nas demais classes as maiores
médias ocorreram em março de 2006, sendo que na classe de 14,01 - 16,00
mm a diferença entre os valores médios de K nas duas épocas de coleta é
mais acentuada, havendo uma tendência de aumento com o comprimento.
Tanto a relação entre massa e comprimento, quanto o fator de condição,
são comumente utilizados como indicativos de higidez tanto de larvas como de
adultos de peixes.
Algumas medidas obtidas das larvas também podem ser consideradas
como indicativos da condição de higidez como a altura da cabeça (AC), altura
do corpo na região da nadadeira peitoral (AP) e altura do corpo na região do
ânus (AA). Pode-se observar que os valores médios para essas variáveis
apresentaram-se maiores em março de 2006 para todas as medidas citadas e
em todas as classes de tamanho (Fig. 16).
A classe de CP de 6,01 - 8,00 mm não pôde ser representada na análise
para que a medida fosse tomada corretamente. Já para AP e AA essa classe
de CP mostra uma diferença mais acentuada entre as duas épocas, inclusive,
não havendo sobreposição das barras de desvio padrão.
O valor médio do resultado da divisão AC/DO para setembro de 2005 foi
1,77 com o desvio padrão de 0,14, enquanto em março de 2006 o valor médio
foi 1,85 com desvio padrão de 0,12.
De maneira geral, para os indicadores analisados, pode-se observar
que, além da tendência das variáveis aumentarem seus parâmetros com o
comprimento das larvas, os valores médios obtidos para setembro de 2005 e
para março de 2006 ficam mais afastados na medida em que larvas de maior
tamanho são analisadas, pelo aumento dos valores obtidos para o mês de
março. Desta forma, reconhece-se um padrão para a condição larval: melhores
condições relativas foram identificadas para as larvas coletadas em março de
2006, em comparação com aquelas de setembro de 2005.
4.3. CONDIÇÃO APARENTE
A partir dos itens levados em consideração para a avaliação da condição
aparente, larvas avaliadas como frágeis representaram 23,6% do total de
larvas avaliadas em setembro de 2005 e 13,8% do total de larvas para março
de 2006. A freqüência de larvas frágeis em cada classe de comprimento
padrão (Fig. 17) mostra, para as duas épocas amostradas, que a incidência de
larvas frágeis é maior nos exemplares com menor comprimento. Para setembro
de 2005 esse decréscimo da freqüência de larvas frágeis na medida em que
Também foram realizadas análises buscando-se verificar se existem
diferenças, no que se refere à fragilidade das larvas, entre as estações de
coleta. Na figura 18 encontram-se as distribuições de freqüências de larvas
frágeis para cada uma das estações. De maneira geral, em ambos os cruzeiros
ocorreram larvas com sinais de fragilidade em diferentes proporções, mas em
ambos os cruzeiros houve predominância de larvas sem sinais de fragilidade
aparente. Em termos espaciais, é possível visualizar a localização das
estações em que está representada a ocorrência de mais que 50% de larvas
frágeis (assinaladas) e as que apresentaram 50% ou menos de larvas frágeis
(não assinaladas), além de verificar quais possuem uma diferença significativa
entre larvas frágeis e não-frágeis (Fig. 19). As estações 7771, 7775, 7776,
7782, 7784, 7785, 7790 e 7791 em setembro de 2005 e, em março de 2006, as
estações 7967, 7977, 7978, 7986, 7988, 7995 e 7997 apresentaram diferença
significativa, através do teste χ2, no número de larvas com condição aparente
mais frágil.
4.4. PROTEÍNAS HIDROSSOLÚVEIS
Foram estimados os valores médios de quantidade de proteína por larva
e representados para classes de CP para as duas épocas de coleta (Fig. 20).
Com exceção da classe de 14,01-16,00 mm, os valores médios
apresentaram-se mais elevados em março de 2006, em comparação com os de apresentaram-setembro de
2005, para cada uma das classes de comprimento, sugerindo melhores
Os valores médios da relação proteína/DNA também foram analisados,
expressos em classes de CP, para as duas épocas de coleta, e corroboram os
anteriores, com valores mais altos para as larvas de março de 2006 (Fig. 21).
Nas tabelas 4 e 5 estão expressos os valores médios citados acima,
para quantidade de proteína e relação proteína/DNA respectivamente, além do
número de exemplares em cada classe de CP. É possível verificar que o valor
médio de proteína para a classe de 6,01 - 8,00 mm quase dobra em março de
2006 se comparado ao de setembro de 2005. Para a relação proteína/DNA
essa diferença encontra-se ainda maior para a mesma classe de CP, quase
triplicando seu valor em março de 2006.
Os resultados obtidos para as avaliações da quantidade de proteínas
vão de encontro ao padrão já observado na morfometria: houve diferenças nos
resultados dos indicadores de higidez das larvas entre as épocas de coleta.
Além disso, no caso específico das análises sobre proteínas, larvas das
menores classes de tamanho também apresentaram diferenças acentuadas
entre as épocas de coleta.
Em termos espaço-temporais, foi verificada, para cada estação de
coleta, a freqüência de larvas com valores de quantidade de proteína (Fig. 22)
e proteína/DNA (Fig. 23) abaixo da média estimada para cada classe de
comprimento. Em setembro de 2005 as estações oceanográficas com menores
índices de larvas com quantidade de proteína abaixo da média foram as de
número 7771, 7775, 7784, 7785 e 7789, portanto com maior quantidade de
larvas em melhores condições em termos percentuais. Considerando-se o
mesmo critério, as estações 7967, 7987, 7988 e 7995 são as que apresentam
à relação proteína/DNA, em setembro de 2005, a estação 7771 foi a única a
apresentar menos do que 50% de larvas com valores abaixo da média,
enquanto as estações 7775, 7785 e 7789 apresentaram exatamente 50% de
larvas nessas condições. Em março de 2006, as estações 7967, 7977, 7987,
7988, 7993 e 7995 apresentaram menos do que 50% de larvas com valores
abaixo da média, mostrando que um número maior de lavas por estação
oceanográfica apresentam melhores condições nesse período (Fig. 23).
É possível observar a localização das estações contendo maior
freqüência de larvas com baixos valores de quantidade de proteína e a
existência de desvio significativo, através do teste
χ
2, na figura 24. Emsetembro de 2005, houve diferença significativa nas estações 7790 e 7797 com
predomínio de larvas com quantidade de proteína abaixo da média e, na
estação 7771 com predomínio de larvas acima da média; já em março de 2006,
houve diferença significativa nas estações 7978, 7986 e 7997 com maior
freqüência de larvas abaixo da média e nas estações 7967 e 7988 com maioria
de larvas acima da média (Fig. 24).
Para a análise de proteína/DNA é possível observar algumas diferenças
em relação à análise anterior. Em setembro de 2005 existe um maior número
de estações apresentando diferença significativa (7780, 7782, 7790, 7791,
7797 e 7804) e todas elas com predomínio de larvas com quantidade de
proteína/DNA abaixo da média; já para março de 2006 a estação 7986, que
apresentou diferença significativa na análise de quantidade de proteínas, não
Não é possível identificar um padrão na distribuição espacial das
estações com maiores porcentagens de larvas com quantidade de proteína
abaixo da média: tanto na região mais costeira e próxima da boca da baía de
Santos, quanto mais ao largo, ocorrem estações oceanográficas com larvas em
5. DISCUSSÃO
5.1. HIDROGRAFIA
As escalas temporais e espaciais dos processos hidrográficos que
ocorrem na plataforma continental podem variar enormemente. As águas
encontradas na plataforma continental são bastante energéticas e, como as
condições termohalinas podem sofrer modificações rapidamente, influenciadas
por processos hidrodinâmicos e de interação oceano-atmosfera, as condições
observadas em um cruzeiro oceanográfico, com duração de alguns dias,
dependem de uma série de processos físicos que ocorrem em escalas
temporais e espaciais diversas (Castro, 1996). O cenário apresentado nesse
trabalho é fruto de coletas realizadas de forma pontual, em estações de coletas
pré-estabelecidas e nos intervalos de tempo que duraram os cruzeiros. Em
conseqüência disso, não se podem acessar informações referentes aos
processos que estavam em vigência nos momentos anteriores às coletas.
As condições hidrográficas encontradas em março de 2006 vão de
encontro com vasta literatura sobre Oceanografia, reportando o fenômeno de
ressurgência na Plataforma Continental Sudeste do Brasil (Ikeda et al., 1974;
Miranda, 1982; Matsuura, 1986; Campos et al., 2000; Rezende, 2003; Mattos et
al., 2004; entre outros), especialmente no verão. Esse fenômeno é mais
intenso em regiões ao norte da área de estudo, especialmente ao largo do
cabo Frio (RJ). Assim, a entrada de águas mais frias na plataforma adjacente
ao estuário de Santos pelas camadas mais ao fundo, provenientes da porção
Em setembro de 2005 é possível notar uma maior influência de águas
estuarinas sobre a plataforma continental, ocasionando um gradiente de
salinidade bem demarcado, principalmente, próximo à baía de Santos. Não foi
possível encontrar literatura relatando a variabilidade temporal das descargas
do estuário de Santos sobre a plataforma.
Em ambas as situações, forçantes atmosféricas influenciam diretamente
os processos em questão. No caso da intrusão da Água Central do Atlântico
Sul, a ação do vento é um dos fenômenos determinantes, enquanto a
precipitação é agente importante na descarga estuarina (Castro & Miranda,
1998; Lopes, 2006; Lopes et al., 2006). Assim, mudanças nas condições
atmosféricas, que podem ocorrer em questão de dias ou até mesmo de
algumas horas, locais ou remotas, possivelmente irão alterar a ocorrência e
intensidade de processos como os observados aqui.
Foi observada estratificação vertical da coluna d’água para a área de
estudo, tanto em setembro de 2005 como em março de 2006. Através da
análise das seções de densidade fica clara a existência não só da
estratificação quanto de certa estabilidade gravitacional na coluna d'água,
principalmente para março de 2006, quando os gradientes de densidade são
mais acentuados. Ainda em março de 2006 é possível observar que tais
gradientes de densidade são dominantemente decorrentes dos próprios
gradientes de temperatura. É interessante notar que na radial 3, nos 15
primeiros metros de profundidade, as feições estão mais relacionadas à
variação na salinidade, uma vez que a temperatura variou pouco nessa faixa
específica de profundidade, mas dos 15 metros para o fundo a temperatura
situação diferente: como a temperatura quase não variou, os gradientes de
salinidade é que originam a estratificação no campo de densidade.
É importante ressaltar esse efeito da salinidade na distribuição da
densidade, uma vez que grande parte dos trabalhos em Oceanografia Biológica
apenas considera o perfil termal para acessar informações sobre estratificação
da coluna d'água. Como pode ser visto nesse estudo, uma descarga de água
menos salina na região costeira pode gerar uma estratificação vertical que não
depende de variações na temperatura.
5.2. HIGIDEZ DAS LARVAS
Os resultados sobre a relação entre massa e comprimento padrão pode
gerar a falsa impressão de que as larvas coletadas em setembro de 2005
apresentam-se em melhor condição do que as larvas coletadas em março de
2006. O que de fato ocorreu foi um excesso de larvas frágeis em setembro de
2005 que não puderam ter suas massas medidas, o que tornou a inclinação da
curva ajustada mais acentuada do que estaria se fossem incluídas essas larvas
frágeis.
Essa impossibilidade de medição da massa das larvas mais frágeis e/ou
danificadas gera um vício de amostragem, uma vez que as larvas que teriam
menores valores de massa para dado comprimento não estão representadas.
Ainda, como as larvas mais frágeis são mais freqüentes nos menores valores
de CP, há uma subamostragem, em relação ao número de indivíduos para a
medida da massa, das larvas menores. Assim, optou-se pela não utilização