1413-8670/© 2012 Elsevier Editora Ltda. All rights reserved.
www.elsevier.com/locate/bjid
The Brazilian Journal of
INFECTIOUS DISEASES
A r t i c L E i n f o
Article history:
received 15 August 2011 Accepted 17 november 2011
Keywords: Probiotic
Lactobacillus rhamnosus Herpes simplex virus type 1
A b s t r A c t
Background: Due to the emergence of drug resistance in herpes simplex virus type 1 (HsV-1), researchers are trying to find other methods for treating herpes simplex virus type 1 infections. Probiotic bacteria are effective in macrophage activation and may have antiviral activities.
Objective: this study aimed at verifying the direct effect of Lactobacillus rhamnosus, a probiotic bacterium, in comparison with Escherichia coli, a non-probiotic one, on HsV-1 infection, and determining its effect on macrophage activation for in vitro elimination of HsV-1 infection. Methods: the above bacteria were introduced into HsV-1 infected Vero cells, and their effects were examined using both Mtt and plaque assay. to determine macrophage activation against in vitro HsV-1 infection, J774 cells were exposed to these bacteria; then, macrophage viability was examined with the Mtt method, and tumor necrosis factor alpha (tnf-α), interferon-gamma (ifn-γ), and nitric oxide (no) assessments were performed using the ELisA method.
Results: A significant increased viability of macrophages was observed (p < 0.05) in the presence of Lactobacillus rhamnosus before and after HsV-1 infection when compared with Escherichia coli as a non-probiotic bacterium. However, tumor necrosis factor α concentration produced by Escherichia coli-treated J774 cells was significantly higher than Lactobacillus rhamnosus-treated J774 cells (p < 0.05). interferon-gamma and no production were not different in the groups treated with Escherichia coli or with Lactobacillus rhamnosus.
Conclusion: the results of this study indicate that Lactobacillus rhamnosus enhances macrophage viability for HsV-1 elimination and activation against HsV-1 more effectively, when compared with non-probiotic Escherichia coli. it also seems that receptor occupation of macrophage sites decreases HsV-1 infectivity by both of the studied bacteria.
© 2012 Elsevier Editora Ltda. All rights reserved.
Original Article
In vitro study of the effect of a probiotic bacterium
Lactobacillus rhamnosus against herpes simplex virus type 1
Soghra Khani
a, Mohammad Motamedifar
a*, Hossein Golmoghaddam
b,
Hamideh Mahmoodzadeh Hosseini
c, Zahra Hashemizadeh
aaDepartment of Bacteriology and Virology, Shiraz HIV/AIDS Research Center (SHARC), Shiraz University of Medical Sciences, Shiraz, Iran bDepartment of Immunology, Shiraz University of Medical Sciences, Shiraz, Iran
cSchool of Pharmacy, Students’ Research Committee,Tabriz University of Medical Sciences, Tabriz, Iran
*Corresponding author at: Department of bacteriology and Virology, shiraz HiV/AiDs research center (sHArc), shiraz University of Medical sciences, shiraz, iran
Introduction
Herpes simplex virus-1 (HsV-1) can cause two types of infection. the initial infection, most often asymptomatic, involves the epithelial cells and can be latent in the sensory nerve ganglia. in a few infected individuals, the latent viruses are activated under different stress conditions, for instance sunshine,
hormonal changes, fever, appearing as cold sore.1 HsV-1 is the
agent of many diseases such as gingivostomatitis, sore throat (pharyngitis), herpetic whitlow, keratoconjunctivitis, herpes encephalitis and meningitis. in immunosuppressed patients, it
can be a chronic destructive process.1-3 both specific and
non-specific defense mechanisms against HsV-1 are important. Host genetic background, macrophages, nK cells, and t-cells and
antibodies are important factors in the defense against HsV-1.1,2
Macrophage activation can also reduce infection by the virus.4
Anti-viral drugs including acyclovir, vidarabine, famciclovir, valacyclovir, pencyclovir are widely used in the treatment of
HsV-1 infection.1,2 nowadays, the occurrence of mutations in viral
DnA polymerase and thymidine kinase has led to an increase
in drug resistance to nucleoside analogs such as acyclovir.4,5 to
find appropriate alternatives, researchers suggest seeking new
chemical and plant sources of therapy for HsV-1 infections.6-10
considering the role of specific and non-specific host defenses against HsV-1, some therapies can work to strengthen and activate the innate and specific immune
system components against HsV-1.11 coordinating operations
of macrophages in anti-herpetic response occur during the first hour of virus invasion. the first defensive line is production and secretion of cytokines such as type i interferon (ifn) and
tumor necrosis factor (tnf). subsequently, iL-12 and ifn-γ
are produced, thereby resulting in natural killer cell and macrophage activation. the production of active oxygen radicals (ros) and of no are auxiliary mechanisms to improve anti-viral
response.12-15 Macrophages have receptors for molecules such
as lipopolysaccharide, mannose, and unmethylated cPG repeats
of bacteria.16 signals resulting from the combination of these
factors increase the secretion of a wide range of cytokines, which strengthen the innate immune system and activate the specific immune system, especially t cells, against pathogens
such as viruses17,18 Among them, inf-γ launches various
activities in macrophages, including antimicrobial activity, causing eradication of intracellular pathogens. Macrophage activity is one of the first steps in preventing virus proliferation (HiV, HsV, VZV) in infected cells. Also, antigen processing and presentation to th1 lymphocytes by macrophages induce
cell-mediated immunity.15-18 no induction of the macrophages in
an inflammatory response is also an innate immune defense
system.19 no has different tasks such as killing infected cells,
tumor cells, and parasitic pathogens. Antiviral effect of no in
various viral infections has also been documented.20
Probiotic bacteria such as Lactobacillus and Bifidobacterium,
which typically ferment dairy foods in human and animal digestive tracts, can balance the immune system and increase the specific and non-specific immune responses. they are important as immunomodulatory bacteria in the body’s defense against
pathogens such as viruses (HiV, VZV).21,22 Various studies have
shown that Lactobacillus rhamnosus (strain GG), a species of probiotic
bacteria, has beneficial effects in treating rotavirus gastroenteritis
in infants.23,24 several studies from in vitro systems, animal
models, and humans, suggests that probiotics can enhance both
specific and nonspecific immune responses.25,26
in a study, it was demonstrated that consumption of lactic acid bacteria in guinea pigs has anti-influenza effects. Also,
taking 1 × 109 cfu/mL of lactic acid bacteria decreases genital
herpes complications caused by HsV-2.27 in another study,
the effect of probiotic bacteria for prevention and treatment of viral diarrhea caused by rotavirus, astrovirus, calicivirus, enteric adenovirus and coronavirus was reviewed, and the helpful role of probiotic bacteria in limiting viral diarrhea
was documented.26 in an in vitro study, it was demonstrated
that probiotic bacteria increase the antiviral response against
vesicular stomatitis virus (VsV) by activating macrophages.11
so, the emergence of antiviral drug resistance observed in HsV-1 makes it necessary to find other alternatives for treating HsV-1 infection. considering the important role of macrophages in eliminating viral infections and the effective role of probiotic bacteria in the removal of viral infections (including HsV), this study aimed to investigate: 1) the direct
effect of Lactobacillus rhamnosus, a probiotic bacterium, on
HsV-1; and 2) its effect on macrophage activation for in vitro
elimination of HsV-1 infection. in this way, the possible use of probiotics to inhibit HsV-1 infection would be scientifically proven as an alternative treatment.
Methods
Bacteria and cytotoxic assays
Lyophilized standard strains of Lactobacilli rhamnosus (Ptcc
1637) and E. coli (Ptcc 25923) were provided by the iranian
research organization for science and technology, tehran,
iran. Lactobacilli rhamnosus was cultured to log phase growth
in L-cystein HcL supplemented by Man rogosa sharp (Mrs) broth (Himedia – india) under anaerobic conditions at 37°c, overnight. in order to isolate single bacterial colonies, the harvested broth was transferred to Mrs agar (Himedia – india) and incubated under the described conditions for 24-36 hours.
the maximum number of bacteria introduced into the antiviral assays was determined as the highest number of bacteria that was not cytotoxic to the macrophage cell line J774 (data not shown).
the value was determined to be 1 × 108 cfU/mL of each strain.
to prepare 1 × 108 cfU/mL of viable bacteria for stimulation
experiments, isolated colonies were suspended in Pbs. the optical density (oD) was measured by spectrophotometer at 530 nm. colony concentration was determined via internal Mcfarland standard oD comparison.
the percentage of adherent bacteria was determined by co-incubation of bacteria with the macrophage cell line for
90 minutes, at 37°c, in a humidified atmosphere of 5% co2.
order to prepare inactivated bacterial suspension, the culture
was frozen at 1 × 108 cfU/mL count and thawed in boiling
water for 3 cycles. Also, E. coli was grown to log phase growth
in EMb agar (Merck – Germany) at 37°c overnight.11
Virus
HsV-1 was isolated from the lip lesions of a patient and was confirmed by neutralization test using guinea pig anti-HsV-1 serum (niH – UsA) and monoclonal anti-HsV-1 antibodies
against HsV glycoproteins D and G.28
HsV-1 was grown to a titer of 106/mL in Vero cell line, in the
maintenance medium of Dulbeccos modified Eagles growth medium (DMEM, sigma) supplemented with 2% fetal bovine serum (Gibco – Germany), 0.14% (v/v) sodium bicarbonate, 100 U/mL penicillin, 100 μg/mL streptomycin sulphate, and 0.25 μg/mL amphotericin b. the virus was stored at −70°c until used. for the antiviral assay, 100 μL of the virus with a plaque-forming unit of 50% per mL was used.
Cells
the murine monocyte/macrophage cell line J774 cells were obtained from the Pasteur institute, tehran, iran. they were grown until confluence in rPMi 1640 media (Gibco – Australasia) containing 10% fetal bovine serum (fbs), and supplemented with 100 μg/mL streptomycin and 100 iU/mL penicillin in tissue culture flasks (nunc – Denmark), at 37°c,
in an atmosphere of 5% co2.
Also, confluent Vero African green monkey kidney cells (razi institute, Hesarak – iran) were prepared in DMEM, supplemented with 8% fetal calf serum. for Vero cells maintenance, DMEM medium was used supplemented with only 2% fetal calf serum. in order to evaluate cell viability, the trypan blue dye (Merck –
Germany) exclusion method was performed.29
Plaque assays
Possible inhibitory effect of the studied bacteria on HsV-1 was investigated using plaque reduction assay, as previously
described.10 Phosphate buffered saline (Pbs)-washed confluent
Vero cells in 24-well plates (nunc – Denmark) were treated with the maintenance medium containing plates (nunc – Denmark). they were treated first with the maintenance
medium, containing 1 × 108 cfU/mL of live or bacterial debris
suspension of E. coli and L. rhamnosus, for 90 min, and then
with 50 PfU/100μL HsV-1, for 40 min. in another series of the experiments under the described conditions, the cells were first exposed to the virus and then to the bacteria; they were also simultaneously exposed to the bacteria and virus. After removing these treatments, the monolayers were infected with 50 PfU/mL of HsV-1 for one hour. subsequently, the monolayers were overlaid with 1% carboxymethyl cellulose (cMc) in the maintenance medium, and were incubated at
37°c with 5% co2 for four days. controls for each series of the
experiment included uninfected cell monolayers, and virus-infected treated or untreated cells with the bacteria. the virus plaques formed on the cells were fixed with methanol for
10 min, and stained with 0.5% crystal violet solution.28
MTT assay
the Mtt assay (3-4, 5 dimethyl thiazol- 2 yl)-2, 5- diphenyl tetrazolium bromide) (sigma – UsA) was used to assess the living cells according to measured active mitochondrial metabolic function. the assay was able to indicate potential beneficial
effects of antiviral agents.29 briefly, after the incubation
period (24 hours at 37°c in presence of 5% co2) on a 96-well
plate containing 90 μL of serial dilution of cell suspension
with the cell concentration of 1 × 105, 5 × 104, 2.5 × 104,
1.25 × 104, 6,250 cell/mL, the wells were washed. then, 10μL
of ready Mtt dye was added. the plate was incubated at 37°c for 3-4 hours. After this period, the contents were centrifuged for 10 min at 300 rpm, the supernatant was removed, and the cells were suspended in 100 μL DMso. the optical density of
each well was determined at 570 nm, using an ELisA reader.30
Assays of the effect of J774 cell culture supernatant treated with L. rhamnosus on HSV-1 infected Vero cell
1 × 105 cell/mL of J774 cells were incubated in a 24-wells
plate, at 37°c, in the presence of 5% co2 for 24 hours;
1 × 108 cfU/ mL living and/or bacterial debris suspension was
added to a separate well plate. Prior to the Pbs washing, the cells were remained at 37°c for 90 min to adhere the bacteria to the cells. the plate was incubated at 37°c in the presence
of 5% co2 for another 48 hours until no and other cytokines
were released. in the next stage, 90 μL of the supernatant of J774 cells treated with bacteria were transferred to the plate containing 50 PfU/100 μL HsV-1 infected Vero cells. After 45 min, the percentage of viable cells was measured using the Mtt test, as described above. the wells containing non-infected Vero cells and free supernatant of J774 cells treated with bacteria were considered as controls.
Assays of the effect of L. rhamnosus on J774 cell line activation against HSV-1
24-hour-incubated plates of 1 × 105 cell/mL of J774 cells were
inoculated with 1 × 108 cfU/mL living and/or bacterial debris
suspension for 90 min, and then infected with 50 PfU/100 HsV-1 for 40 min.
in another series of experiments, the cells were infected with 50 PfU/100 μL HsV-1 for 40 min, and then inoculated with
1 × 108 cfU/mL living and/or bacterial debris suspension for
90 min. the plates were incubated at 37°c in the presence of
5% co2 for 24 hours. the percentage of metabolically active
cells was determined via Mtt test, as described. the well containing neither virus nor bacterium was considered as the negative control. Prior to performing the Mtt test, the
supernatant of all the cells was collected to measure no, infγ,
and tnfα levels.
NO measurement
to measure no concentration, levels of released no were
measured using the Griess reaction.31 100 μL of supernatant
from incubated 1 × 108 cell/mL of J774 cells only, J774 cells
and inactivated E. coli, at 37°c in the presence of 5% co2 for 48 hours, were added to the ELisA plate separately, with 100 μL of modified Griess reagent (Merck – Germany). After 10 min incubation at room temperature, the absorbance at 450 nm was measured. the concentration of no was calculated
using nano2 standard curve.
INFγ and TNFα levels determination
infγ and tnfα levels were detected using a commercial
enzyme-linked immunosorbent assay (ELisA) kit (bender Medsystem – UsA) according to the manufacturer’s instructions.
100 μL of supernatant from incubated 1×108 cell/mL of J774
cells only, J774 cells treated with L. rhamnosus, inactivated
L. rhamnosus, E. coli and inactivated E. coli at 37°c in the presence
of 5% co2 for 48 hours were added to the ELisA plate separately.
Statistical analysis
All statistical analyses were performed using the statistical Package for social sciences (sPss) version 11.5 software (chicago, iL, UsA). the difference between the two groups (L. rhamnosus and E. coli) was tested using the Mann-Whitney test. the J774 cells only were used as a control. p-values less than or equal to 0.05 were considered as statistically significant.
Results
L. rhamnosus significantly enhanced HSV-1 infected Vero cells viability
compared to HsV-1 infected control cells, L. rhamnosus
significantly increased cell viability of the HsV-1 infected Vero cells either pre or post HsV-1 infection [(49 ± 1.1%, 47.6 ± 1.5%,
respectively) (41.7 ± 1.1%) (p ≤ 0.05 )]. E. coli showed a similar
effect on the elevation of Vero cells viability (fig. 1).
L. rhamnosus reduced HSV-1 plaques on Vero cells
compared to HsV-1-infected Vero cells (83.3 ± 2.08) as the control, significantly lower numbers of plaques were detected
in the Vero cell culture treated with L. rhamnosus either
pre or post HsV-1 infection (68.1 ± 1, 71 ± 2, respectively). A statistically similar reduction of HsV-1 plaques was observed
in the Vero cells exposed to E. coli. (table 1).
Experiment conditions Mean numbers of plaques ± SD
Vero cell monolayer 0
Vero + HsV-1 83.3 ± 2.08
Vero + HsV-1 + L. rhamnosus 71 ± 2.0 Vero + HsV-1 + E. coli 73 ± 3.0
Vero + L. rhamnosus 0
Vero + E. coli 0
Vero + L. rhamnosus + HsV-1 68 ± 1.0 Vero + E. coli + HsV-1 69 ± 0.57
Table 1 - Comparison of number of plaques of Vero cells infected with HSV-1 in different experiments
Supernatant of treated macrophage J774 culture with
L. rhamnosus and E. coli significantly increased Vero cell viability
As shown in fig. 2, the supernatant of pre-treated macrophage
J774 cells with live and/or inactive L. rhamnosus significantly
increased the Vero cell viability in comparison to cells infected with the virus only (74.9 ± 0.95 – 51 ± 2.9) (p ≤ 0.05). However,
E. coli treatment caused a lower Vero cell viability, but the difference was not significant (p ≥ 0.05).
Fig. 1 - Comparison of Vero cells viability percentages under different conditions using the MTT test.
Fig. 2 - Comparison of cell viability percentages of HSV-1 infected Vero cells affected by J774 culture supernatant
treated with L. rhamnosus and E. coli suspensions.
A, Vero cells; B, Vero cells+HSV-1; C, Vero cells+HSV-1+
J774 supernatant treated with live L. rhamnosus; D, Vero
cells+HSV-1+ J774 supernatant treated with live E. coli;
E, Vero cells+HSV-1+ J774 supernatant treated with
inactive L. rhamnosus; F, Vero cells+HSV-1+ J774
supernatant treated with inactive E. coli; G, Vero cells+ J774
supernatant.
optical density (570 nm)
% cell via
bility
V
er
o cell
V
er
o cell + HsV
-1
V
er
o cell + HsV
-1 +
E.
coli
V
er
o cell +
E.
coli
V
er
o cell +
E.
coli
+ HsV
-1 +
L.
rhamnosus
V
er
o cell +
E.
coli
+ HsV
-1
V
er
o cell +
L.
rhamnosus
V
er
o cell +
L.
rhamnosus
+ HsV
-1
120
100
80
60
40
20
0
0.71 0.29 0.452 0.451 0.348 0.328 0.338 0.33
0.77 0.39 0.58 0.53 0.52 0.51 0.76
optical density (570nm) 120
100 80 60 40 20 0
% cell via
bility
A 100
b 51
c
74.9 D
68.3 E
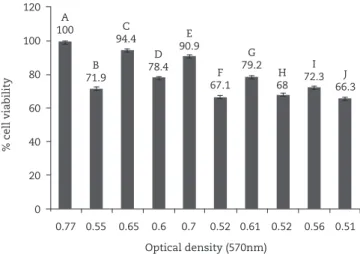
L. rhamnosus caused increased viability of macrophage J774 cells infected with HSV-1
As shown in fig. 3, a significantly increased viability of J774
cells was observed after L. rhamnosus treatment of J774 cells
pre and post-infection with HsV-1 versus non-treated J774 cells and E. coli-treated J774 cells (p ≤ 0.05).
Pro-inflammatory cytokines and NO production were stimulated in the presence of bacteria
co-incubation of J774 cells with suspension of either live
or inactive L. rhamnosus or E. coli stimulated macrophages
to produce no, tnf-α and ifn-γ (table 2). E. coli caused
significantly higher tnf-α production compared to
L. rhamnosus (p = 0.019). A remarkable induction of tnf-α
production was observed by J774 cells exposed to live E. coli
suspension pre and post infection with HsV-1, as well as
to inactive E. coli (p = 0.027). such effect was not seen with
L. rhamnosus. Live or inactive suspensions of L. rhamnosus or
E. coli did not stimulate no or ifn-γ secretion in HsV-1 infected J774 cells.
Discussion
clinical interventional studies have identified those specific probiotics that have antiviral effects via shortening of the infectious period or decreasing the risk of certain viral infection. the best finding was the beneficial influence of
probiotics on viral diarrhea in infants and children.32
in a study on anti HsV-2 effect of selected vaginal
Lactobacillus strains (L. brevis cD2, L. salivarius fV2, L.
plantarum fV9), conti et al. demonstrated the inhibitory effect of living bacterial cells on HsV-2, through several mechanisms, including interference with early steps of virus replication such as binding/entry, and the production of lactic acid and hydrogen peroxide, with a direct antiviral
effect.33
there are also two separate reports demonstrating the protective effects of various lactic acid bacteria on
VsV-infected porcine epithelial and macrophage cell lines.34,11
With regard to the proven antiviral effect of probiotic strains and to researchers’ attempts to create an appropriate protocol to treat or prevent HsV-1 infection, we decided to test a probiotic bacterium as a candidate for such a purpose.
in this study we evaluated the in vitro effects of
L. rhamnosus, a probiotic bacterium, directly on HsV-1 infections, and its influence on stimulation of no release and
tnf-α and ifn-γ production as pro-inflammatory cytokines
following L. rhamnosus activation of murine monocyte/
macrophage J774 cell line.
our results clearly demonstrated that L. rhamnosus exposure
enhanced the viability of J774 cell line up to 90%, which is remarkable in comparison to the results reported by ivec et
Experiment conditions NO (μg/mL) Mean± SD
IFNγ (pg/mL) Mean ± SD
TNF-α (pg/mL) Mean ± SD
J774 0 7.1 ± 0.57 17.3 ± 2.08
J774 + HsV-1 0.11 ± 0.015 14.5 ± 0.73 286 ± 4.6
J774 + L. rhamnosus 0.14 ± 0.01 14.1 ± 1.52 205 ± 2.88
J774 + E. coli 0.12 ± 0.02 16.1 ± 1.52 452 ± 2.51
J774 + L. rhamnosus + HsV-1 0.13 ± 0.01 16.6 ± 1.52 236 ± 2.51
J774 + HsV-1 + L. rhamnosus 0.11 ± 0.01 16 ± 1.15 276 ± 2.3
J744 + E. coli + HsV-1 0.11 ± 0.01 14.1 ± 1.52 323 ± 3.0
J774 + HsV-1 + E. coli 0.12 ± 0.01 15.1 ± 1.52 372 ± 3.0
J774 + inactive L. rhamnosus + HsV-1 0.11 ± 0.01 13.1 ± 1.52 227 ± 2.64
J774 + inactive E. coli + HsV-1 0.11 ± 0.01 13 ± 1.0 368 ± 3.05
Table 2 - The levels of TNF-α, NO and IFN-γ measured by ELISA in different conditions on this study
Fig. 3 - J774 cells viability treated with Lb .rhamnosus and
E. coli suspensions; A, J774; B, J774 + HSV-1; C, J774 +
Lb .rhamnosus; D, J774 + E. coli; E, J774 + L.rhamnosus +
HSV-1; F, J744 + E. coli + HSV-1; G, J774 + HSV-1 +
L. rhamnosus; H, J774 + HSV-1 + E. coli; I, J774 + inactive
L. rhamnosus + HSV-1; J, J774 + inactive E. coli + HSV-1.
A 100
b 71.9
c 94.4
D 78.4
E 90.9
f 67.1
G 79.2
H 68
i 72.3 J
66.3 120
100
80
60
40
20
0
% cell via
bility
0.77 0.55 0.65 0.6 0.7 0.52 0.61 0.52 0.56 0.51
al. (30% viability of VsV infected 3D4/21 cell line). the cause of viability increase can be due to difference of cell line, VsV infectious capacity versus HsV-1, or probiotic species between the two studies. Also, we detected more protective effect of live bacteria than inactive bacteria on the exposure of viral infected macrophages, a finding consistent with ivec et al.’s report. A mechanism involved in the protective effect of probiotic bacteria against virus particles is competition for cell surface
attachment. this has already been proven in vitro, and reported
as a feasible mechanism for the protection of cells in the case
of VsV infection.6,11
Preservation of a cpG motif in bacterial DnA during inactivation of bacteria has been reported to suppress the
apoptotic pathways and enhance the cell viability.11,35
our study demonstrated the higher viability of HsV-1
infected macrophage J774 cells treated with L. rhamnosus
when compared to E. coli treatment, although the reason is
unclear. one explanation of lower cells viability in the case of
E. coli exposure compared to L. rhamnosus can be related to
increased tnf-α production induced by E. coli, which causes
more cell apoptosis. We showed more tnf-α production induced
by E. coli exposure pre- and post-HsV-1 infection of the cells
compared to L. rhamnosus. the presence of lipopolysaccharide in
the cell wall of E. coli is a potential stimulator for tnf release.36
increased viability of infected Vero cells by a supernatant of
J774 cells exposed to L. rhamnosus might result from no, tnf-α
and ifn-γ production, as pro-inflammatory cytokines.11,15 this
suppressed HsV-1 proliferation. no release is another antiviral mechanism, which has also been previously reported. in fact, significant increase of no release was reported in co-incubated macrophage exposed to probiotic bacteria in the presence of
ifn-γ; however, ivec et al. stated that ifn-γ is not an important
factor in the stimulation of no release.11,37
in our study, J774 cells co-incubated with live or inactive
L. rhamnosus or E. coli were capable of inducing tnf-α, ifn-γ
and no production. concentration of ifn-γ and its dependent
mediator, no, is lower than those of a previous report. However, in that report, probiotic bacteria were applied with
exogenous ifn-γ, so it can be concluded that the simultaneous
presence of ifn-γ with probiotic bacteria might be necessary
for the production of high levels of no.37 in the case of
inactive bacteria, cell wall components such as peptidoglycan,
lipoteichoic acid and cpG motifs can induce no, tnf-α and
ifn-γ production by cells; however, their levels were lower than
those induced by live bacterial exposure.11
in conclusion, L. rhamnosus was able to induce several
antiviral effects against HsV-1, which might be through different mechanisms including competing with virus for cell surface attachment, increasing macrophage viability, following stimulation of pro-inflammatory responses. thus, our results
suggest future in vivo evaluation of L. rhamnosus for increasing
the immune defense in the cells as a probable candidate for HsV-1 infection co-therapy.
Acknowledgements
this article is taken from the Msc thesis of soghra Khani for Medical Microbiology and was financially supported by shiraz
University of Medical sciences Grant no. 4008. We also wish to thank Mr. Pedram taleazadeh shirazi for his technical assistance.
Conflict of interest
All authors declare to have no conflict of interest.
r E f E r E n c E s
1. Geo fb, carroll Kc, butel Js, Morse sA. Herpesviruses. in: Jawetz, Melnick and Adelberg`s. Medical Microbiology. 24th
ed. McGraw-Hill companies UsA; 2007. p. 428-37. 2. Kapikian A, Hoshino Y, chanock r. rotaviruses. in: Knipe
D, Howley P, editors. fields Virology. 5 ed. Philadelphia: Lippincott Williams & Wilkins; 2007. p. 2479-2601. 3. Whitley rJ. Herpes simplex virus infection. semin Pediatr
infect Dis. 2002;13(1):6-11.
4. biswas s, tiley Ls, Zimmermann H, et al. Mutations close to functional motif iV in HsV-1 UL5 helicase that confer resistance to HsV helicase-primase inhibitors, variously affect virus growth rate and pathogenicity. Antiviral res. 2008;80(1):81-5.
5. frobert E, thouvenot D, Lina b, et al. Genotyping diagnosis of acyclovir resistant herpes simplex virus. J Pathol biol. 2007;55(10):504-11.
6. Pietka-ottlik M, Wójtowicz-Młochowska H, Kołodziejczyk K, et al. new organoselenium compounds active against pathogenic bacteria, fungi and viruses. chem Pharm bull. 2008;56(10):1423-27.
7. Wang H, Vincent ooi E, Ang oJr Po. Antiviral activities of extracts from Hong Kong seaweeds. J Zhejiang Univ sci b. 2008;9(12):969-76.
8. Kratz JM, Andrighetti-fröhner cr, Kolling DJ, et al. Anti-HsV-1 and anti-HiV-1 activity of gallic acid and pentyl gallate. Mem inst oswaldo cruz. 2008;103(5):437-42. 9. rajbhandari M, Mentel r, Jha PK, et al. Antiviral activity of
some plants used in nepalese traditional medicine. Evid based complement Alternat Med. 2009;6(4):517-22.
10. Koch c, reichling J, Kehm r, et al. Efficacy of anise oil, dwarf-pine oil and chamomile oil against thymidine-kinase-positive and thymidine-kinase-negative herpesviruses . J Pharm Pharmacol. 2008;60(11):1545-50.
11. ivec M, botic t, Koren s, et al. interactions of macrophages with probiotic bacteria lead to increased antiviral
response against vesicular stomatitis virus. Antiviral res. 2007;75(3):266-74.
12. trinchieri G. interleukin-12 and the regulation of innate resistance and adaptive immunity. nat rev immunol. 2003;3:133-46.
13. Denis M. Human monocytes/macrophages: no or no no? J Leukoc biol. 1994;55(5):682-4.
14. MacMicking J, Xie QW, nathan c. nitric oxide and
macrophage function. Annu rev immunol. 1997;15:323-509. 15. Ellermann-Eriksen s. Macrophages and cytokines in the early
defence against herpes simplex virus. Virol J. 2005;2:59-89 16. Miettinen M, Lehtonen A, Julkunen i, Matikainen s.
28. Motamedifar M, nekooeian AA, Moatari A. the effect of hydroalcoholic extract of olive leaves against herpes simplex virus type 1. iran J Med sci. 2007;32:222-6.
29. Leary JJ, Wittrock r, sarisky rt, et al. susceptibilities of herpes simplex viruses to penciclovir and acyclovir in eight cell lines. Antimicrob Agents chemother. 2002;46(3):762-8. 30. bergamini A, Perno cf, capozzi M, et al. tetrazolium-based
colorimetric assay for quantificationof HiV-1-induced cytopathogenicity in monocyte-macrophages exposed tomacrophage-colony-stimulating factor. J Virol Methods. 1992;40(3):275–86.
31. Green Lc, Wagner DA, Glogowski J, et al. Analysis of nitrate, nitrite, and [15n] nitrate in biological fluids. Anal biochem. 1982;126(1):131-8.
32. szajewska H, Mrukowicz JZ. Probiotics in the treatment and prevention of acute infectious diarrhea in infants and children: a systematic review of published randomized, double-blind, placebo-controlled trials. J Pediatr Gastroenterol nutr. 2001;33(suppl 2):17-25.
33. conti c, Malaciino c, Mastromarino P. inhibition of herpes simplex virus type 2 by vaginal Lactobacilli. J Physiol Pharmacol. 2009;60(suppl 6):19-26.
34. botic t, Klingberg tD, Weingartl H, et al. A novel eukaryotic cell culture model to study antiviral activity of potential probiotic bacteria. int J food Microbiol. 2007;115(2):227-34. 35. Jozsef L, Khreiss t, filep JG. cpG motifs in bacterial DnA
delay apoptosis of neutrophil granulocytes. fAsEb J. 2004;18(14):1776–8.
36. Abbas AK, Lichtman AH, Pillai s. cellular And Molecular immunology. Elsevier, Philadelphia,.6th ed. 2007.
37. Korhonen r, Korpela r, saxelin M, et al. induction of nitric oxide synthesis by Probiotic Lactobacillus rhamnosus GG in J774 Macrophages and Human t84 intestinal Epithelial cells. inflammation. 2001;25(4):223-32.
17. Morita H, He f, fuse t, et al. cytokine production by the murine macrophage cell line J774.1 after exposure to lactobacilli. biosci biotechnol biochem. 2002;66(9):1963-6. 18. Gessani s, belardelli f, Pecorelli A, et al. bacterial
lipopolisaccharide and gamma interferon induce transcription of beta interferon mrnA and interferon secretion in murine macrophages. J Virol. 1989;63(6):2785-9. 19. Kidd P. th1/th2 balance: the hypothesis, its limitations,
and implications for health and disease. Altern Med rev. 2003;8(3):223-246.
20. Kim DW, cho sb, Yun cH, et al. induction of cytokines and nitric oxide in murine macrophages stimulated with enzymatically digested lactobacillus strains. J Microbiol. 2007;45(5):373-8.
21. Martin L cross. immunoregulation by probiotic Lactobacilli: pro-th1 signals and their relevance to human health. clin Applied immunol. 2002;3(3):115-25.
22. Acheson DW, Luccioli s. Microbial-gut interactions in health and disease. Mucosal immune responses. best Pract res clin Gastroenterol. 2004;18(2):387-404.
23. oberhelman rA, Gilman rH, sheen P, et al. A placebo-controlled trial of Lactobacillus GG to prevent diarrhea in undernourished Peruvian children. J Pediatr. 1999;134(1):15-20.
24. Parvez s, Malik KA, Ah Kang s, Kim HY. Probiotics and their fermented food products are beneficial for health. J Appl Microbiol. 2006;100(6):1171-85.
25. rizzello V, bonaccorsi i, Dongarrà ML, et al. role of natural killer and dendritic cell crosstalk in immunomodulation by commensal bacteria probiotics. J biomed and biotechnol. 2011;473097.
26. takeda K, suzuki t, shimada si, et al. interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei shirota. clin and Experimental immunol. 2006;146:109–115.