Claudia Figueiredo Pereira
Licenciada em Bioquímica
Carbon monoxide, autophagy and cytoprotection
in response to cerebral ischemia
Dissertação para obtenção do Grau de Mestre em Genética
Molecular e Biomedicina
Orientador: Helena Luísa Araújo Vieira,
Investigadora Principal, Centro de Doenças Crónicas,
Faculdade de Ciências, Universidade Nova de Lisboa
Júri:
Presidente: Prof. Dra Margarida Castro Caldas
Arguente(s): Dra Raffaella
Vogal(ais): Dra Helena Luísa Araújo Vieira
Claudia Figueiredo Pereira
Licenciada em Bioquímica
Carbon monoxide, autophagy and cytoprotection in
response to cerebral ischemia
Orientador: Helena Luísa Araújo Vieira, PhD, PI, CEDOC FCM/UNL
Carbon monoxide, autophagy and cytoprotection in response to cerebral
ischemia
Copyright Claudia Figueiredo Pereira, FCT/UNL, UNL
A Faculdade de Ciências e Tecnologia e a Universidade Nova de Lisboa têm o direito, perpétuo e sem limites geográficos, de arquivar e publicar esta dissertação através de exemplares impressos reproduzidos em papel ou de forma digital, ou por qualquer outro meio conhecido ou que venha a ser inventado, e de a divulgar através de repositórios científicos e de admitir a sua cópia e distribuição com objectivos educacionais ou de investigação, não comerciais, desde que seja dado crédito ao autor e editor.
vii
Acknowledgments
I would like to express my sincere gratitude to all the people that direct and indirectly helped during this thesis. I would also like to acknowledge to the good conditions of the institute were I worked: CEDOC.To my supervisor, Dr. Helena Vieira, for the teaching, the patience, the good moments of scientific divagation, for all dedication, support and the constant messages of courage never forgotten. Thank you for teaching me again how to love science. In the personal field, it is a great honor to meet people like you, unique.
To Dr. Roberto Motterlini and Dr. Roberta Foresti, for the excellent example of hard-working. For the opportunity for meeting other scientific universe, by the way you welcomed me, making me feel as part of the picture. For the conversations, the advices, through the open door. Thank you.
To the financial support provide by e-COST, (COST Action BM1005), allowing me a new scientific experience in France.
To CO-team, Claudia, Sofia and Sara. To Claudia and Sofia, thank you for being the great person that you are, for all the wonderful moments along this year, letting me growth up by my own but always near. For the advices, for just hear me, for stopping your work to help me, for “pra quê?” and “num bai
dar”. You are “big” persons.
To CEDOC colleagues, it was a great pleasure to share laboratory with you all. The Cilia Regulation and Disease Lab: Petra, Catarina, Pedro, Barbara and Susana. In special, to João, André, Catarina, Petra, Pedro, Neuza and Telmo, for the good moments, for the sharings, a handful of smiles and laughter, “almoços”, “Gémeos”, “Jantaradas”, “gostos musicais”and “loucuras”. To Pedro for the friendship along this two last years, for being a good surprise, for “boleias” and “disparates”. To Telmo for all the informatics help and for Neuza to the good mood.
To equipe 3 of INSERM U995, Ben, Jane-Lowis, Sarah, Nicolas, Romain, Jeremy, Lolita and Coraly, it was a great pleasure to meet you all, to have a possibility to know other world of science. To Romain, merci pour tout, for the teaching how to live life, “fais des bêtises, mais fais-les avec enthousiasme”. À minha “Familia” que apesar de longe estamos sempre perto, pelos esforços de encontros, por terem contribuido para aquilo que sou hoje.
Para a Lídia um muito obrigada por estares sempre presente, por esta amizade/irmandade que já dura há tanto tempo, basicamente por seres tu.
A minha eterna gratidão aos meus pais, por todo o esforço, pelas palavras de apoio, conselhos. À minha mãe por ser esse ser fantástico que me ensina a lutar todos os dias sem nunca baixar os braços. Obrigada por seres o pilar da nossa família. Ao meu pai, pelo feitiozinho, pelos ensinamentos
viii
de “enginhocas”, por me ensinar que tudo tem solução e nem sempre o caminho mais fácil é o melhor.
Aos meus Avôs, os meus dois exemplos de vida, nos vossos braços sinto-me segura, obrigada. Ao Téta, por seres tu, como já há uns anos disse, por tudo e por nada, por estares sempre lá, por acreditares em mim, por me fazeres sentir capaz de alcançar o mundo, pelas surpresas, por seres o meu Pintas e a minha Personagem.
…This Master thesis was only possible due to you all. In other words thank you very much, for nothing and for all.
ix
...
xi
Abstract
There is an increasing need for promoting neuroprotection against cerebral ischemia, which is the main cause of brain damage in adults. Astrocytes are the most abundant cells inboard the central nervous system (CNS), being known as key glial cell for promoting neuronal survival and homeostasis. It is more established in nowdays that astrocytic dysfunction contributes to neurodegenerative processes. Although, carbon monoxide is a well renown as a lethal and toxic gas due to its high affinity to hemoglobin, CO exerts anti-apoptotic, anti-inflammatory and anti-proliferative functions. Recent studies showed likewise that CO induces autophagy, promoting therefore cytoprotective and anti-inflammatory effects. Autophagy is a major catabolic pathway, known as an autodigestive process that degrades cellular organelles and proteins, playing an important role in cellular homeostasis during environmental stress.Due to the great interest on the signaling and cytoprotective actions of CO, novel strategies have been put forward to exploit the potential therapeutic effects of this gaseous molecule. One of these approaches consist on the development of CO-releasing molecules (CO-RMs), compounds that deliver small quantities of CO to tissues and first identified by the group of Motterlini and co-workers. The aim of this Master thesis was to study the action of CORM-A1, a boron-containing compound that spontaneously releases CO, against cell death in primary culture of astrocytes. In particular, we examined the role of CORM-A1 in autophagy, mitophagy and cell metabolism.
Here, we demonstrated that CORM-A1 promotes the induction of autophagy in primary culture of astrocytes. Furthermore, autophagy is directly involved in the cytoprotective effect of CORM-A1 against cell death. In some preliminary experiments we have shown that CORM-A1 also induced mitophagy, while autophagy and inhibition of cell death promoted by CORM-A1 seem to occur under hypoxia (5% of oxygen). This master thesis has addressed several important questions on the role of CO in astrocyte function but also opened to many other important questions on the mechanism of action of CO. For instance, future work must be undertaken in order to explore whether CO-mediated induction of reactive oxygen species (ROS), which play an important role in cell signaling, which are the factors directly involved in mitophagy and the cross-talk between apoptosis and modulation of autophagy.
xiii
Resumo
Isquemia cerebral é a principal causa de danos cerebrais em adultos, levantado assim uma necessidade crescente para a promoção da neuroprotecção. Os astrócitos são as células mais abundantes no interior do sistema nervoso central (SNC), estando envolvidas na homeostasia e sobrevivência neuronal. Um das causas já estabelecidas para patologias neurodegenerativas está relacionado com a disfunção destas células.O monóxido de carbono (CO) é geralmente conhecido como um gás tóxico letal devido à sua elevada afinidade com a hemoglobina, no entanto já se sabe que o CO apresenta propriedades benéficas para o organismo, entre elas anti-apoptóticos e anti-inflamatória e anti-proliferativa, para além destas propriedades, estudos recentes mostraram também que CO induz autofagia. Autofagia, é um processo degradação de organelos celulares e proteínas denaficadas, exercendo um papel importante na homeostase celular.
Em linha com o conhecimento já existente, o objetivo deste trabalho foi estudar a acção de CO contra a morte celular em cultura primária de astrócitos, em particular o papel da CORM-A1 na indução de autofagia, mitophagy e metabolismo celular. CORM-A1 é uma nova fonte de CO, designada como “CO-releasing molecules”, desenvolvida por Motterlini e colegas.
Neste trabalho demonstramos que CORM-A1 induz autofagia em astrócitos. Além disso, autofagia está directamente envolvida no efeito citoprotector de CORM-A1 contra a morte celular, bem como a indução de mitofagia. Para além disto, verificamos que efeito da CORM-A1 em hipoxia continua a ser citoprotector.
Em suma, durante esta dissertação de mestrado muitas “janelas” de investigação foram abertas, deixando caminho para futuras abordagens.
xv
Index
ACKNOWLEDGMENTS ... VII
... ... IX
ABSTRACT ... XI
RESUMO ... XIII
INDEX ... XV
FIGURE INDEX ... XVII
TABLE INDEX ... XIX
ABBREVIATIONS ... XXI
I.
INTRODUCTION ...1
CENTRAL NERVOUS SYSTEM ...3
1
The role of astrocytes in central nervous system ... 4 1.1
CARBON MONOXIDE ...7
2
Heme oxygenase (HO) ... 7 2.1
Biological functions of carbon monoxide ... 8 2.2
Carbon monoxide in the CNS ... 9 2.2.1 CO Releasing molecules ... 10 2.3
AUTOPHAGY ... 11
3
Autophagic Flux ... 11 3.1 Mitophagy ... 13 3.2 BNIP3 ... 14 3.2.1PINK1 and Parkin ... 14 3.2.2
xvi
AIM ... 19
4
III.
MATERIAL AND METHODS ... 21
Material, solutions and medium ... 23 4.1
Medium and culture material ... 23 4.1.1
Preparation of Buffer solutions ... 23 4.1.2
Preparation of CORM-A1 ... 24 4.1.3
Antibodies ... 24 4.1.4
Primary Cultures of Astrocytes: isolation and maintenance ... 25 4.2
Role of CORM-A1 in primary culture of astrocytes ... 26 4.3
Inducers and inhibitors of cell death and autophagy ... 26 4.3.1
Induction of autophagy ... 27 4.3.2
Induction of cell death and assessment of cell viability ... 28 4.3.3
Western Blot ... 30 4.4
Imunofluorescente Microscopy ... 31 4.5
Quantitative PCR - Measure of mitochondrial DNA ... 31 4.6 Statistical Analyses ... 32 4.7
IV.
RESULTS ... 33
RESULTS ... 35
5
CORM-A1 prevents cell death in primary culture of astrocytes ... 35 5.1.1
CORM-A1 promotes autophagy induction. ... 37 5.1.2
Autophagy involvement in the cytoprotective role of CORM-A1 ... 39 5.1.3
The action of CORM-A1 in mitophagy ... 40 5.2
CORM-A1 apparently promotes mitophagy activation by BNIP3 expression... 40 5.2.1
The role of CORM-A1 in mitophagy ... 41 5.2.2
The role of CORM-A1 in mitochondrial biogenesis ... 43 5.2.3
The role of oxygen in CORM-A1 action ... 43 5.3
Cytoprotective effect of CORM-A1 in astrocytes is maintained at low concentration of oxygen, 5%O2 . 44
5.3.1
V.
DISCUSSION ... 47
xvii
Figure Index
Figure 1.1 – Schematic representation of several brain cell populations. ... 3
Figure 1.2 – Representative scheme of communication between neurons and astrocytes- trypartide synaps. ... 6
Figure 2.1- Endogenous pathway production of carbon monoxides.. ... 7
Figure 2.2 - Chronologic discover of CORM'S.. ... 10
Figure 3.1- Autophagic flux. ... 12
Figure 3.2 - Mitophagy process by activation of PINK1 and Parkin induction. ... 15
Figure 4.1 - Autophagic Flux... 26
Figure 4.2- Scheme of autophagy induction in primary culture of astrocytes along time. ... 28
Figure 4.3 - Scheme of the primary culture of astrocytes treatment for cell viability measurement; ... 29
Figure 4.4 – Example fo gates used to evaluate the cell viability of cells. ... 29
Figure 4.5 - Example of Standard curve, used for protein quantification.. ... 30
Figure 5.1 – Purity of primary culture of astrocytes.. ... 36
Figure 5.2 – CORM-A1 confers protection against cell death.. ... 37
Figure 5.3 - CORM-A1 induces autophagy in astrocytes. ... 38
Figure 5.4 - CORM-A1 confers protection against apoptosis by inducing autophagy. ... 39
Figure 5.5 - CORM-A1 seem to be involved in mitophagy induction in astrocytes.. ... 41
Figure 5.6 – Optimization of the assay to measure co-localization of autophagy/mitophagy in astrocyte primary cultures.. ... 42
Figure 5.7 – Effect of CORM-A1 on cytochrome c oxidase activity and mitochondria biogenesis. ... 43
Figure 5.8 – CORM-A1 confers protection against apoptosis by inducing autophagy at low levels of oxygen. ... 45
xix
Table Index
Table 4.1- Reagents used culture cells, for the extraction of astrocytes ... 23
Table 4.2 - Reagents utilized in the technique of Western Bolt. ... 23
Table 4.3 - Description of conditions and features of the used antibodies. ... 24
Table 4.4 - Description of cells cultured in different plates with different concentration for the different techniques. ... 25
Table 4.5 - Inducers and Inhibitors of autophagy and cell death inducers ... 27
Table 4.6 - Protocol for gel preparation at 12 and 15% of acrylamide. ... 30
xxi
Abbreviations
• ATP- Adenosine triphosphate; • AXO – Alexander disease; • Ca+2 – calcium cation;
• cAMP- Cyclic adenosine monophosphate; • cGMP- Cyclic guanosine monophosphate; • CO – Carbon monoxide;
• COHb – Carboxyhemoglobin;
• CORM’s - Carbon monoxide releasing molecules; • CNS – Central nervous system;
• DiOC – 3,3′-dihexyloxacarbocyanine iodide;
• DMEM - Dulbecco's modified eagle medium; • dMfn – Factor mitufusion;
• DNA - Deoxyribonucleic acid;
• EBSS – Earle’s balancel salt solution; • EDTA - Ethylenediamine tetraacetic acid; • FBS – Fetal bovine serum;
• GABA – Neurotransmitter glutamate; • GC – Guanilate cyclase;
• HCQ – Hydroxycloroquine; • HO- Hemoxygenase; • IF – Imunofluorescense;
• MMP- Mitochondrial membrane permeabilization; • PBS – Phosphate buffer saline;
• PE – Phosphatidylethanolamine; • PI – Propidium iodide;
• PI3K- phosphatidylinositol 3-kinase; • PNS- Peripheral Nervous System;
• Q-PCR – Quantitative polymerase chain reaction; • RT – Room temperature;
• WB – Western blot; • WM- Wortmannin;
• ROS – Reactive species of oxygen; • t-BHP - tert-Butylhydroperoxide;
I.
Introduction
State of the art
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
3
Central nervous system
1
The nervous system coordinates the actions and transmits signals in the body and it consists of two main parts: the central nervous system (CNS) and the peripheral nervous system (PNS). The central nervous system is the processing center and consists of two main organs, spinal cord and brain. It collects, integrates and sends information to the peripheral nervous system. Both organs, brain and spinal cord, are protected by three layers of connective tissue named as meninges (Roberts et al. 2004).
At the cellular level, nervous system is defined by the presence of nerve cells, also known by neurons, and special types of supporting cells, known as glial cells (figure 1.1). The neurons are accountable for functions that are unique to the nervous system, while the glial cells are non-neuronal cells that primarily support and protect the neurons (Bordal et al. 2010).
Figure 1.1 – Schematic representation of several brain cell populations. Addapted from.(Halassa, Fellin et al. 2007; Halassa, Fellin et al. 2009)
Neurons are post-mitotic and highly polarized cells, meaning that they develop, in the course of maturation, distinct subcellular domains responsible for different functions. Morphologically three major regions compose neurons: cell body, dendrites and axon. The cell body, or soma, contains the nucleus and the major cytoplasmic organelles. Multiple short dendrites are extensions of the receiving surface and arise from perikaryon, ramifying over a certain volume of gray matter. Dendrites can differ in size and shape, depending on the neuronal type. Finally, the axon conducts nerve impulses to other neurons or to muscle cells, which is the most extended part of the neuron. Each neuron might have multiple dendrites, but just one axon (Roberts et al. 2004).
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
4
Neuronal cells are classified into two broad groups, projection neurons and interneurons. Projection neurons are characterized by their capacity to respond to stimuli with an electrical discharge, a nerve impulse, and, further, by their fast conduction of the nerve impulse over long distances. This capacity is allocated to axon that is particularly built to conduct the nerve impulse from cell body to other cells. Therefore a signal can be transmitted in millisecond along the body, the CNS or between them. Interneurons mediate cooperation among neurons that rely grouped together (Roberts et al. 2004). Between all the cell types in the body, neurons are the most dependent on oxygen and nutrient supply, since the specialization on the transmission of information does not allow the existence of energy reservoirs. In accordance, brain receives about 15% of the cardiac output at rest (Roberts et
al. 2004; Bordal et al. 2010). Moreover, just few minutes of blood flow interruption may cause neuronal
cell death, occurring during cerebral ischemic stroke, which is the third cause of death in west countries.
The CNS is composed by three types of glial cells, oligodendrocytes, astrocytes and microglia. In 1859, Rudolph Virchow, studied this set of cells and coined as neuroglia, or “nerve glue”. This name derives from the notion that glial cells served as a kind of glue, an inactive connective tissue holding neurons together in the CNS. Glial cells are the major cells present in the brain and indispensable for neuronal functioning (Roberts et al. 2004; Vilhardt et al. 2005).
Oligodendrocytes are the main responsible for the formation of myelin sheath in the CNS and are involved in a short number of processes. The main one is during brain development as responsibles for myelin production. The other cell type that is present in CNS is microglia, which originates from mesoderm and is known as the macrophages of nervous tissue (Vilhardt et al. 2005). Some studies indicate that microglia constitute 5 to 20% of all the glial cells and are distributed throughout all parts of the CNS. These cells have a particularly function in CNS that is the role of cleaning the extracellular medium, by “scanning” the environment for foreign material and sick or dead cellular elements (Roberts et al. 2004; Bordal et al. 2010). Finally the third glial cells are astrocytes, which are discussed in the next section.
The role of astrocytes in central nervous system
1.1
Astrocytes are the most abundant cells inboard the CNS, outnumbering neurons in a ratio of 10:1, constituting about 20-50% of the total human brain volume (Sagduyu K.et al. 2002; Roberts et al.
2004; Izhikevich et al. 2007). They are known for being a heterogeneous cell population based on their morphology, function and expression of different sets of receptors, transporters, ions channels and neurotransmitters (Son, Jeong et al. 2005; Sofroniew and Vinters et al. 2010; Allaman, Bélanger
et al. 2011).
In the beginning of Neurobiology field, they were the most poorly understood neural cells, thought for simply being the cells that occupy the spaces between neurons. However astrocytes revealed to be
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
5
involved in many important functions in the brain: modulation of metabolism, neurotransmitters and homeostatic functions, establishing and maintaining the essential nature of interactions between neurons (Sagduyu K.et al. 2002; Son, Jeong et al. 2005). Based on in vivo and in vitro approaches, itis already recognized that astrocytes produce and release growth factors for the modulation of distinct neuronal subpopulations at the level of: morphology, proliferation, differentiation and survival. Additionally, astrocytes also act on the control of development and function of astrocytes and oligodendrocytes. The ability of astrocytes in producing and responding to growth factor and cytokines is the major mechanism underlying the development and regenerative capacity of CNS (Roberts et al. 2004).
Since the last decade, astrocytes have been categorized in two subtypes, protoplasmic or fibrous, based on their unique cytoarchitectural and phenotypic features which allow them to respond to changes in the microenvironment (Sofroniew; Vinters et al. 2010). Protoplasmic astrocytes are found throughout all gray matter, whereas the fibrous astrocytes are only found in white matter. The
protoplasmic astrocytes exhibit a morphology of several stem branches, while fibrous astrocytes have
longer branches and 50 to 60 long fiber-like processes, but both make extensive contacts with intraparenchymal blood vessels via specialized processes called endfeets. These endfeets express glucose transporters of the Glut-1 type and are a possible site of glucose uptake (Sagduyu K. et al. 2002). In this sense astrocytes have been shown to have an important role in neurovascular and neurometabolic coupling. Likewise, for enabling the dynamic coupling of cerebral blood flow with energy demand, the astrocytes release vasoactive substance (Allaman, Bélanger et al. 2011).
At the metabolic level, astrocytes are the only cell types in the brain that comprise the energy storage molecule glycogen, and that respond to glutamatergic activation by increasing their rate of glucose utilization and releasing lactate in the extracellular space, which might, in turn, be used by neurons to sustain their energy demands (Sagduyu K. et al. 2002). Other homeostatic functions have been demonstrated, including: (i) ion and water homeostasis, (ii) defense, against oxidative stress, (iii) scar formation and tissue repair, (iv) modulation of synaptic activity via the release of gliotransmitters, and (iv) synapse formation and remodeling (Sofroniew; Vinters et al. 2010; Allaman, Bélanger et al. 2011). Astrocytes are powerfully coupled together by gap junctions, containing aqueous pores, which are permeable to ions and other molecules with low molecular weight (Sagduyu K. et al. 2002). A set of biological important molecules, comprising nucleotides, small peptides, cAMP, sugars, amino acids, inositol triphosphate (IP3) and Ca2+ have accesses by gap junctions (Sofroniew; Vinters et al. 2010). Since the last years the role of astrocytes in the brain has raised a special interest for investigation, these star-shaped cells that lack axons not only supports neuronal activity, but can also modulate the neurotransmission by modulating synapses. This process is defined as the “tripartide synapse” and depending on intracellular levels of Ca+2, astrocytes release gliotransmitters (e.g. glutamate) – in a process termed gliotransmission - that have feedback actions on neurons (figure 1.2). The term “tripartite synapse” refers to a concept in synaptic physiology based on the demonstration of the existence of bidirectional communication between astrocytes and neurons (Halassa, Fellin et al. 2007; Halassa, Fellin et al. 2007; Halassa, Fellin et al. 2009). Glutamate is the most common amino acid
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
6
and neurotransmitter present in brain tissue, which is an excitatory neurotransmitter. Glutamate can be generated from neuronal metabolism of glucose and glutamine is its principal precursor of synaptically released glutamate. For instance, glutamate, which is uptaken by astrocytes, which convert glutamate into glutamine and then release it to the extracellular space, while glutamine is uptaken by neurons, restarting the cycle generating again glutamate and GABA, which are potent excitatory and inhibitory neurotransmitters, respectively (Sagduyu K.et al. 2002; Roberts et al. 2004).
Figure 1.2 – Representative scheme of communication between neurons and astrocytes- trypartide synaps. Adapted (Halassa, Fellin et al. 2007)
Considering the pivotal role of astrocytes in the brain and the strong cooperation between neurons and astrocytes, direct evidences point to an important role of astrocytes in several pathologies, either through the loss of normal function or the gain of defective functions. A striking example of one disorder caused by a primary dysfunction of astrocytes is Alexander disease (AXD), the first identified human neurological pathology (Allaman, Bélanger et al. 2011). Consequently, in contrast to the classically accepted paradigm that brain function results exclusively from neuronal activity, there is an emerging view, in which brain function actually arises from the coordinated activity of a network comprising both neurons and glia cells (Sagduyu K. et al. 2002; Roberts et al. 2004).
As stated above, the role of astrocytes in brain functioning is of extreme importance. Although many advances have been made in this area, the full understanding of astrocytic intracellular events is still unknown. Consequently in this master thesis we have a great interest in studying astrocytes due to their crucial role on brain homeostasis and neuronal functioning.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
7
Carbon monoxide
2
Carbon monoxide (CO) is a diatomic colorless small molecule, invisible, chemically inert, nonirritant, and odorless gas (Idriss, Blann et al. 2008). It is, commonly known as a lethal gas and toxic air pollutant, however, CO is an endogenously produced gaseoustrasmitter. The chemical composition of CO was identified in the 18th century. In 1857, Claude Bernard demonstrated that CO could cause asphyxia, by reversely moving oxygen from hemoglobin and forming carboxyhemoglobin (COHb). During the subsequent years, CO toxicity has been lengthily studied (Ozaki, Kimura et al. 2012). Only after almost one century, in 1949, it was demonstrated that CO is endogenously produced through its identification in exhaled human air (Sjöstrand et al. 1949). In 1968 heme oxygenase (HO) was identified as the enzyme that catalyzes heme and endogenously produces CO (Ozaki, Kimura et
al. 2012). CO is generated by oxidation of the heme group, in particular the α-methene bridge carbon atom of the heme porphyrin ring, being a catabolic byproduct of hemoglobin in the body under abnormal conditions (figure 2.1) (Ozaki, Kimura et al. 2012; Ryter ; Choi et al. 2013).
Figure 2.1- Endogenous pathway production of carbon monoxides (Ozaki, Kimura et al. 2012). Heme group is converted in biliverdin, ferriciron and carbon monoxide by the action of heme oxygenase. Biliverdin is after converted in bilirubin in the presence of biliverdin reductase.
Heme oxygenase (HO)
2.1
Heme oxygenase is the rate-limiting enzyme, responsible for metabolizing heme molecules into biliverdin, ferrous iron, and CO (figure 3). HO activity and expression is induced by several different stress stimuli, such as heavy metals or reactive oxygen species (ROS), and is involved in both physiological and pathological processes (Ozaki, Kimura et al. 2012). Tenhunen and colleagues were the first to propose the existence of three isozymes, HO-1, HO-2 and HO-3 (Idriss, Blann et al. 2008). HO-1 and HO-2 have a clear and significant biological role, while HO-3 has a substantially lower catalytic activity and is less studied (Ozaki, Kimura et al. 2012). Although HO-1 and HO-2 catalyze the same reaction, they differ in expression patterns, HO-1 expression is induced in response to a numerous stimulus, while HO-2 is constitutively expressed (Motterlini, Clark et al. 2002). The inducible
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
8
HO-1, is a ubiquitous heat shock protein (HSP32), which is particularly protective safeguarding the cell against diverse stress-related conditions (such as redox). HO-1 is upregulated in response to oxidative stress, hyperthermia, hypoxia and proinflammatory stimuli in different tissues and has been shown to exert potent cytoprotective and antiapoptotic properties. The common feature between these inducers is their ability to produce ROS at low levels, and thus with signaling functions, suggesting that HO-1 provides potent cytoprotective effect (Idriss, Blann et al. 2008). While, HO-2 is abundant in brain, testis, and liver, and is responsible for particularly high HO activity in these organs during the steady conditions (Ryter; Choi et al. 2013).
One of the products originated by heme group degradation is biliverdin, which is rapidly converted in bilirubin by the action of biliverdin reductase. These two molecules are natural antioxidants, usually are present in serum in high levels and are also considered to be responsible for the HO-induced cytoprotection. (Idriss, Blann et al. 2008).
It is well known that byproducts generated during the heme catabolism are described as having potential protective role. Nevertheless, in several types of injury the protection has been attributed mainly to CO.
Biological functions of carbon monoxide
2.2
Firstly, CO was described as a putative neural messenger and is now recognized as a signaling molecule exerting essential regulatory roles in a variety of physiological and pathological process in cardiovascular, nervous and immune systems. Some CO’s cytoprotective effects are: anti-inflammatory, anti-apoptotic, anti-atherogenic and anti-proliferative (Motterlini et al. 2010).
Motterlini and colleagues shown that in isolated aortic tissue CO induced vessel relaxation and prevent coronary vasoconstriction as well as acute hypertension in vivo (Motterlini, Clark et al. 2002; Clark, Naughton et al. 2003). The CO biological effects concerning the inflammatory processes seem to involve the stimulation of soluble guanylate cyclase (sGC) and cyclic guanosine monophosphate (cGMP) production, but it can also include modulation of MAPK activation and of calcium-dependent potassium channel activity, in smooth muscle models (Ryter, Alam et al. 2006).
Some evidence suggests that endogenous CO is a neurotransmitter in CNS. In 1993, Verma and colleagues demonstrated, in primary olfactory neuron culture, the action of CO as endogenous neuronal transmitter regulating cyclic guanosine monophosphate (cGMP) and guanylate cyclase (GC) (Verma, Hirsch et al. 1993; Ozaki, Kimura et al. 2012). Brouard et al in 2000 demonstrated for the first time the anti-apoptotic property of the system HO-1/CO in endothelial cells. Since then, it has been shown that CO confers resistance against cell death following ischemia-reperfusion trauma, in many different models, namely lung, heart, kidney, liver and brain (Ryter, Alam et al. 2006; Queiroga, Almeida et al. 2012).
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
9
Carbon monoxide in the CNS
2.2.1
In 2007, Chora et al. described that CO reduced neuroinflammation in experimental autoimmune encephalomyelitis, a model of multiple sclerosis. Few years after, in 2009, Zeynalov showed that in mice brain CO reduced injury after transient middle cerebral artery occlusion. In 2012, Mahan et al. showed that CO prevented cerebral injury resulting from cardiac bypass procedures using deep hypothermic circulatory arrest. In the same year, Yabluchanskiy and colleagues demostrated that a CO-releasing molecule (CORM-3) promotes neuroprotection or neurotoxicity after intracerebral hemorrhage depending on the time of administration (Smith, Mann et al. 2011).
However, in the brain environment, the CO cytoprotective-associated mechanisms have been poorly described. For that reason, along the last years our laboratory has developed a great interest for studying the effect of CO in the prevention of neuronal and astrocytic cell death and its role in mitochondria. Therefore, in 2008, Vieira and colleagues first showed that the preconditioning of murine primary cerebellar granule cells with exogenous CO prevented neuronal apoptosis induced by excitotoxicity and oxidative stress (Vieira, Queiroga et al. 2008). Likewise, CO prevents neuronal death in a perinatal model of cerebral ischemia, by increasing Bcl-2 expression, preventing the release of cytochrome c from the mitochondria and inhibiting caspase-3 activation (Queiroga, Tomasi et al. 2012).
Because astrocytes are crucial cell for cerebral homeostasis and correct functioning, our group has also focused on CO cytoprotection in this neural cell type. In 2010, Queiroga and colleagues demonstrated that CO prevented apoptosis in primary culture of astrocytes by directly inhibiting mitochondrial membrane permeabilization (MMP), with oxidized glutathione and ROS as signaling factor (Queiroga, Almeida et al. 2010). Likewise, CO also prevented astrocytic cell death by improving cell metabolism, namely by: increasing ATP production, increasing cytochrome c oxidase enzymatic specific activity and stimulating mitochondrial biogenesis, in a Bcl-2 dependent mode of action (Almeida, Queiroga et al. 2012).
Up to the date all generated data in our lab was performed using CO gas or CO-saturated solutions. We are now interested in using CO releasing molecules (CORM) that will be described in the next section.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
10
CO Releasing molecules
2.3
Since the last decade, CO has received great attention as a biological regulator that can have an important role as a therapeutic tool. The main example is the development of CO-releasing molecules (CORMs) (figure 2.2), which are organic and organometallic compounds, with the capacity of delivering CO in a time and tissue-specific manner, allowing a significant reduction in carboxyhaemglobin toxicity (Queiroga, Almeida et al. 2012). It is already known the capacity of CORMs to mimic the effect of the gaseous CO, including vessel relaxation, protection against organ ischemia-reperfusion injury, prevention of organ rejection after transplantation, inhibition of inflammatory response and anti-apoptotic properties, highlighting the efficiency of CO transport by these molecules (Motterlini, Clark et al. 2002; Motterlini et al. 2010).
Figure 2.2 - Chronologic discover of CORM'S. Schematic diagram showing the types of bioactive CORM’s identified until this moment, CORM’s are compounds that contain a heavy metal such as nickel, cobalt, or iron surrounded by carbonyl (CO) groups as coordinated ligands. Figure adapted from (Motterlini, Sawle et al. 2005).
In 2003 the first water-soluble CORM, CORM-3 (tricarbonylchloro(glycinato) ruthenium (II)), was described, which was obtained by coordinating the amino acid glycine onto the metal center. CORM-3 is fully soluble in water and presents in its structure a metal carbonyl complex with a half-time of about 1minute in physiological buffers. One of the most recently described CORMs is CORM-A1 (sodium boranocarbonate), like CORM-3, it is fully soluble in water, but does not possess a transition metal in its structure and is able to release CO with a half-life of 21 minutes being slower releaser under physiological conditions, (37ºC and pH 7,4) (Motterlini, Sawle et al. 2005).
The advantage of using this type of molecules is the capability of these small organic and organometallic compounds to deliver CO in a timely and tissue-specific manner, allowing a reduction in the carboxyhaemoglobin formation and toxicity. Opening therefore new windows of opportunity to use CO in clinical applications.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
11
Autophagy
3
Throughout a possible lethal stress, cells respond with rapid metabolic changes to protect themselves against potential damage. This is orchestrated by a multifaceted cellular program, which involves the action of various stress response pathways. One of the key pathways that arbitrates stress-induced metabolic adaptation and control of cell damage is macroautophagy, also known as autophagy (Kroemer, Mariño et al. 2010).
More than 40 years ago Christian de Duve, described a catabolic processes conserved from lower to higher eukaryotes, which is named as autophagy, this word remotes from the Greek and means “selfeating” (Gomes; Scorrano ; Rabinowitz; White et al. 2010). Autophagy is an intracellular degradation system that carries cytoplasmic constituents to the lysosome. There are three types of autophagy; macroautophagy, microautophagy and chaperone mediated autophagy (Mizushima, Levine et al. 2008).
Autophagy is a protective mechanisms that allows cells to survive in response to multiple stressors, such as; starvation, hypoxia, ROS and damaged organelles, strengthening organism defense (Levine; Kroemer et al. 2008; Mizushima et al. 2008). Autophagy is involved in diverse human processes, such as development, longevity, immunity, cancer and neurodegenerative diseases (Kroemer, Mariño et al. 2010; Rabinowitz; White et al. 2010).
Furthermore diverse studies suggest that the lifetime of Drosophila and Caenorhadditis elegans and even mice can be significantly increased by stimulation of autophagy. In the last years autophagy reveals as a protective process (Hansen; Johansen et al. 2011).
Autophagic Flux
3.1
Autophagic flux refers to all steps of the complete process of autophagy (figure 3.1). Autophagic process initiates with the formation of isolate membrane named phagophore (a and b). This phagophore sequesters and engulfs a portion of cytoplasm to form mature autophagosomes (c) the fusion of this structure with a lysosome forms the autolysosome (d), in which occurs the degradation of the engulfed content by the action of acidic lysosomal hydrolases (e) (Mizushima, Levine et al. 2008; Mizushima, Yoshimori et al. 2010). Thus, these three main steps (figure 5- steps a,c,d) allow cellular elimination of damaged or harmful components through catabolism and cellular recycling, which maintain nutrient and energy homeostasis (Xie; Klionsky et al. 2007; Hansen; Johansen et al. 2011). In the absence of stress, autophagy suits a housekeeping function, providing a routine “garbage disposal” to the cells. This function is particularly important to quiescent and terminally differentiated cells, in which the damaged organelles are not disposed by the cell moment of replication (Rabinowitz; White et al. 2010).
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
12
Autophagy is induced by an extensive range of stimulus (Rabinowitz; White et al. 2010). The most usual trigger of autophagy is nutrient deprivation also known as starvation, but other stimuli are involved including damaged organelles, misfolded proteins, DNA damaged, hypoxia, redox stress and mitochondrial damage (Mizushima et al. 2007; Kroemer, Mariño et al. 2010).
Figure 3.1- Autophagic flux (Xie and Klionsky 2007). a,b) Cytosolic material is sequestered by an expanding membrane named as phagophore. c) Resulting in the formation of double-membrane vesicle, autophagosome. d) The outer membrane fuse with a lysosome forming the autolysosome. e) The cargo containing membrane compartment is then lysed and the content is degraded;
Although some of the proteins involved in autophagic process are already known, it is still necessary to disclose many of the cellular factors involved in autophagy. The autophagosome formation requires Beclin-1-class III phosphatidylinositol 3-kinase complex to generate phosphatidylinositol 3-phosphate, as well as generation and insertion of LC3 (Rabinowitz; White et al. 2010). This protein specifically associates with autophagosome membranes and usually is present in the cytosol as soluble form of LC3 (LC3-I). When autophagy is activated LC3-I conjugates with phosphatidylethanolamine (PE) and occurs its lipidation, leading to the conversion in autophagic vesicle-associated form, LC3-II (Yang and Klionsky 2010; Yang and Klionsky et al. 2010; Yen, Shintani et al. 2010; Hansen; Johansen et al. 2011). Thus, LC3-II is widely used as a biomarker for autophagy activation.
Many proteins that possess an LC3-interacting region (LIR) are considered as adaptors proteins to target autophagic degradation. The ubiquitinylation on modified proteins or in mitochondria, is recognized and bound by autophagy receptors, such as p62 (also known as sequestosome1, SQSTM1), Nbr1 (Kirkin, McEwan et al. 2009), as well as BNIP3L (also known as Nix). These receptors bind to mitochondrial membranes by the interaction with LC3 delivering cargo to autophagosome (Novak, Kirkin et al. 2009; Rabinowitz; White et al. 2010; Hansen; Johansen et al. 2011).
Since alterations in autophagy have a role in pathogeneses of several diseases, there is a great interest in identifying compounds that can be used for therapeutic purposes.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
13
Mitophagy
3.2
A billion years ago, aerobic bacteria colonized eukaryotic cells, and this symbiotic relationship resulted in bacteria evolution into mitochondria, while the host cells acquired the ability to metabolically use oxygen. In this moment mammalian cells rely on mitochondria for vital functions such as; pyruvate oxidation, the Krebs cycle, the metabolism of amino acids, fatty acids, calcium homeostasis, steroids and the generation of energy as ATP. Mitochondria also play a significant role in several mechanisms of cell death because they are sources of pro-apoptotic molecules, such as cytochrome c and are decked with proteins of the Bcl-2 family such as the pro-apoptotic protein Bax (Youle; Narendra et al. 2010). On the other hand, mitochondria are an important source of ROS (reactive oxygen species). Mitochondria ROS production contributes to cellular redox signaling, as well as to cellular damage, depending on their concentration (Scherz-Shouval, Shvets et al. 2007; Scherz-Shouval; Elazar 2011; Vives-Bauza; Przedborski et al. 2011). Recent studies suggest that one major sensor of redox signaling in cellular response, between stress adaptation and cell death, is autophagy (Jisun, Samantha et al. 2012).
Brain is rich in mitochondria and uses about 20% of oxygen, while 90% of the consumed oxygen is for generating energy, which makes brain cells particularly sensitive to oxidative stress. Oxidative stress is linked to mitochondrial dysfunction as at the same time mitochondria generate and are the target for ROS. In this way, mitochondria homeostasis is very important for avoiding cellular accumulation of ROS and a loop of damage, thus the number and quality of this organelle is critical to the good cellular functioning. The elimination of mitochondria leads to the renewing of mitochondria named as mitochondria turnover. Mitochondria turnover is dependent on autophagy, but the cross-talk between autophagy, redox signaling and mitochondrial dysfunction is not well understood (Jisun, Samantha et
al. 2012).
Mitophagy was discovered in 2004 in Sacchoromyces cerevisae as well as the first involved protein (Uth1p) (Bhatia-Kiššová; Camougrand et al.). The term mitophagy was suggested by Lamesters and colleges in 2005, to describe the engulfment of mitochondria into vesicles with autophagosome marker, therefore, mitophagy concerns to selective process of autophagy (Youle; Narendra et al. 2010). Following the loss of mitochondria, cell might maintain the population of mitochondria (lengths and shape) by altering the balance between fission and fusion and by inducing mitochondrial biogenesis (Vives-Bauza; Przedborski et al. 2011). In the recent years some studies defend that fission or mitochondrial fragmentation is an event that occurs previously to mitophagy (Poole, Thomas
et al. 2010). Recent evidences suggests that mitophagy from yeast to mammals is the primary
mechanism for the elimination of malfunctioning mitochondria. Thus, mitochondrial remodeling through fission, fusion, mitophagy and mitochondrial biogenesis are important for mitochondrial homoeostasis and quality control (Kim, Rodriguez-Enriquez et al. 2007; Jisun, Samantha et al. 2012).
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
14
BNIP3
3.2.1
The receptors of mitophagy are still a subject of discussion and speculation, since so far little is known about them. Recent studies suggested the role of a mitochondrial outer membrane protein, named Nix or BNIP3L, that is required for mitochondrial clearance during erythrocyte maturation and has a WXXL motif (Kanki, Wang et al. 2009). It may also be a mitochondrial receptor for mitophagy in mammalian cells (Kirkin, McEwan et al. 2009; Novak, Kirkin et al. 2009).
Similarly, BNIP3 that was initially identified in yeast as Bcl-2 and adenovirus E1B 19-kDa interacting protein-3, is present in the membrane of mitochondria when it is overexpressed (Zhang; Ney et al. 2009; Quinsay, Lee et al. 2010). Together with Nix, BNIP3 are mitochondrial proteins that have the ability to induce both cell destinies: survival or death.
BNIP3 and Nix share certain features with the BH3-only subgroup of the Bcl-2 family, such as sequence homology in the BH3 domain, residence in the outer membrane and the ability to interact with Bcl-2 and Bcl-XL, which connects them with cell death modulation (Bellot, Garcia-Medina et al.
2009).
Furthermore these proteins are also related with autophagy by Beclin-1, an important autophagic protein, that interacts with Bcl2 and Bcl-XL, thus, BH3-only proteins may induce autophagy by
competing with Bcl-2 for binding to Beclin-1 (Zhang, Meng et al. 2003; Zhang; Ney et al. 2009).
PINK1 and Parkin
3.2.2
The loss of function of the proteins: Parkin (an E3 ubiquitin ligase) or PINK-1 (PTEN-induced kinase 1, a mitochondrial localized serine/threonine kinase) results in a genotype of recessive familial forms of Parkinsonism (Poole, Thomas et al. 2010). The malfunction of these two proteins is responsible for male sterility in Drosophila, apoptotic muscle degeneration, defects in mitochondrial morphology and increased sensitivity to multiple stresses including oxidative stress (Clark, Dodson et al. 2006). Additionally, Parkin and PINK-1 are also related to mitophagy modulation.
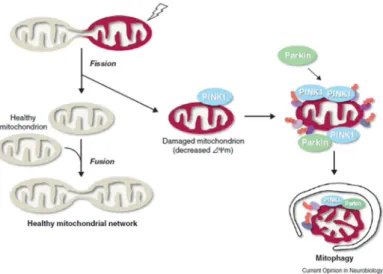
PINK-1 and Parkin are mostly studied in dopaminergic neurons since it was thought that these two proteins are involved in the neurodegenerative Parkinson disease. PINK-1 a constitutively express protein, being proteolysed by mitochondria rhomboid protease PARL, at the mitochondria membrane of healthy mitochondria. This process results in processed forms of PINK-1, which are rapidly degraded by the proteasome. Whenever membrane potential (∆ψm) is reduced there is an accumulation and activation of PINK-1, recruiting Parkin from the cytosol to the mitochondria. Mitochondria accumulation of Parkin leads to their degradation via mitophagy (figure 3.2) (Shiba-Fukushima, Imai et al. 2012).
In 2011 Cui and colleagues found in dopaminergic MN9D mouse cells that the down-regulation of PINK-1 by RNA interference resulted in abnormal mitochondrial morphology, partial loss of mitochondrial membrane potential and increased production of ROS, leading to mitophagy. In these cells mitophagy was associated with up-regulation of Beclin-1 and the opening of mitochondrial
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
15
permeability transition (MPT) pore, suggesting that the mechanisms behind the activation of mitophagy are controlled by opening of MPT pore and by general autophagy regulators (Cui, Fan et al. 2011).All of these processes involving PINK 1/Parkin are thought to be related with the capacity of the mitochondrial machinery regulation through ubiquitination, namely, the mitochondrial fusion-promoting factor mitofusion (dMfn). The steady-state abundance of dMfn is increased by the decrease of PINK1/Parkin complex (Poole, Thomas et al. 2010; Youle; Narendra et al. 2010; Imai; Lu et al. 2011). However, the exact stimuli and mechanism that activate autophagy by the loss of function of PINK-1 and accumulation of Parkin is unclear (Cui, Fan et al. 2011).
Figure 3.2 - Mitophagy process by activation of PINK1 and Parkin induction (Imai; Lu et al. 2011). Mitochondrial fusion and fission events are required for the maintenance of healthy mitochondrial population, in the case of mitochondrial fusion it is thought to facilitate the interchange of internal components. On other hand mitochondrial fusion seems to have an important role in the removal of damaged mitochondria and PINK1/Parkin are likely to be the trigger of this all process. In the case of damaged mitochondrias PINK1 accumulates in the mitochondrial membrane and recruits parkin from the cytosol signaling the mitochondria for degradation via a selective process of elimination – mytophagy;
Mitochondrial depolarization, fission and ROS production (malfunctioning mitochondria) are associated with modulation of mitophagy, but how these potential signals are integrated is not clear, being a scientific subject with broad room for development and research.
Therefore this thesis focuses on understanding CO mode of action in primary culture of astrocytes, namely its capability of cytoprotection against cell death via modulation of autophagy and/or mitophagy.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
19
Aim
4
The role of astrocytes in the CNS and their involvement with malfunction, such as in neurodegenerative diseases, has raised a great interest along the time, since astrocytic function is crucial for neuronal maintenance. The role of CO in the cytoprotection of astrocytes was already demonstrated in previous studies (Queiroga, Almeida et al. 2010; Almeida, Queiroga et al. 2012). Nevertheless, the CO-induced cellular pathways are still a matter of debate. In addition, studies carried out in the recent years, in brain and in other models, introduce a new cellular process of cytoprotection: autophagy, previously claimed as a cell death process (Boya, Reggiori et al. 2013). In summary, and based in all the existent knowledge, this thesis has as a general aim:
The study of the cellular and biochemical pathways involved in CO-induced cytoprotection, in particular the role of autophagy, mitophagy and cell metabolism.
For reaching this aim, three specific objectives will be followed:
1) Understanding whether CO induces autophagy and in particularly mitophagy in astrocytes; 2) The autophagy/mitophagy role in CO-induced cytoprotection in astrocytes;
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
23
This section is reserved to describe the material and methods used to evaluate the experimental hypothesis. The description of the experimental work is divided in five main points.Material, solutions and medium
4.1
Medium and culture material
4.1.1
All plastic material was provided from Sarstedt (Germany) and is sterilized. Glass material and prepared solutions were sterilized by autoclaving at 121ºC for 20 minutes or by filtration using a pore size of 0.22 µM.
Table 4.1- Reagents used culture cells, for the extraction of astrocytes
Reagent Reference Company Final concentration
DMEM low glucose 31885-0230 Gibco – Life
technologies Fetal Bovine Serum
(heat inactivated) (FBS) 41F3715K Gibco- Life technologies 20, 15 and 10% in cell culture medium Penicillin-streptomycin 1514022 Gibco- Life technologies 1% (100U/mL) in cell culture medium
Glucose Sigma 1 g/mL in cell culture
medium
Preparation of Buffer solutions
4.1.2
All the solutions presented in the table below were prepared in milli-Q water:
Table 4.2 - Reagents utilized in the technique of Western Bolt.
Buffers Composition
Phosphate Buffer saline (PBS) 1,54M NaCl,34mM Na2HPO4, 20mM KH2PO4.
With a final pH of 9,4. Cell lysis Buffer
2% (w/v) SDS 50mM tris-HCL
pH= 6.8 Loading buffer (for samples in
WB) 10% (v/v) Glycerol 10mM DTT 0,005% (w/v) Blue Bromofenol T-TBS (WB) 0,25M Tris-HCL; 0,75M NaCL Blocking Buffer (for blocking
WB membranes) T-TBS with 5% (w/v) Milk Running Buffer (protein gel for
WB) 0,25M Tris base,1,92M Glicine, SDS 20%(v/v)
Transfer Buffer (transferring protein from gel to membranes
in WB)
Runing buffer with 10% (v/v) of Methanol
Permeabilization solution
0,1% (w/v) SDS in PBS Blocking solution
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
24
Preparation of CORM-A1
4.1.3
The solution of CORM-A1 was prepared in milli-Q water with a final concentration of 5mM. Then, the solution was filtrated with 0,22 µM filter, aliquoted and stored at -20ºC. For each use, an aliquot was thawed and rapidly added into the culture.
Antibodies
4.1.4
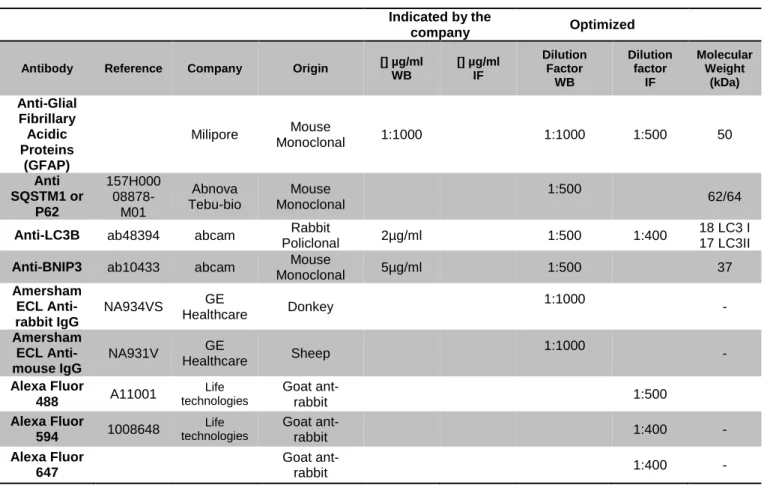
For Western Blot and Immunofluorescence techniques, we used the primary and secondary antibodies described below:
Table 4.3 - Description of conditions and features of the used antibodies. Indicated by the
company Optimized
Antibody Reference Company Origin [] µg/ml WB [] µg/ml IF Dilution Factor WB Dilution factor IF Molecular Weight (kDa) Anti-Glial Fibrillary Acidic Proteins (GFAP) Milipore Mouse Monoclonal 1:1000 1:1000 1:500 50 Anti SQSTM1 or P62 157H000 08878-M01 Abnova Tebu-bio Mouse Monoclonal 1:500 62/64
Anti-LC3B ab48394 abcam Rabbit
Policlonal 2µg/ml 1:500 1:400
18 LC3 I 17 LC3II
Anti-BNIP3 ab10433 abcam Mouse
Monoclonal 5µg/ml 1:500 37 Amersham ECL Anti-rabbit IgG NA934VS GE Healthcare Donkey 1:1000 - Amersham ECL Anti-mouse IgG NA931V GE Healthcare Sheep 1:1000 - Alexa Fluor 488 A11001 Life technologies Goat ant-rabbit 1:500 Alexa Fluor 594 1008648 Life technologies Goat ant-rabbit 1:400 - Alexa Fluor 647 Goat ant-rabbit 1:400 -
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
25
Primary Cultures of Astrocytes: isolation and maintenance
4.2
The animal manipulation was realized according to the recommendation of the Conveção Europeia for
Protecção dos animais vertebratos used for experimental studies and scientific (Directiva do Conselho
86/609/CEE). All efforts were done to minimize the number of used animals and to decrease animal suffering.
Primary culture of astrocytes was obtained from 1-day-old mouse cortex, as described by Schousboe, A., et al. 2001. The animals were rapidly decapitated, brain cortex was removed and the meninges were carefully stripped off, then the cortex was washed in ice-cold phosphate-buffered saline (PBS), and mechanically disrupted. Single-cell suspensions were plated in T-flasks (four hemispheres/75 cm2) in Dulbecco’s minimum essential medium supplemented with 20% (v/v) fetal bovine serum (heat-inactivated), 100units/ml penicillin/streptomycin solution. Cells were maintained in a humidified atmosphere of 7%CO2 at 37 °C.
With the aim of achieving a pure culture of primary astrocytes, after 7 days, when cells reach the confluence, the dark phase cells growing on the astrocytic cell layer were eliminated (such as oligodendrocytes) by vigorous shaking and removing the supernatant. The remaining astrocytes were detached by mild trypsinization using trypsin/EDTA (0.25%, w/v) and were subcultured in DMEM supplemented with 15% (v/v) fetal bovine serum, in a new t-flaks of 75cm2 with a final volume of
15mL, for more two weeks , during this period culture medium was renewed twice a week . Finally at the third week, medium was replaced from 15% (v/v) of fetal bovine serum to 10%. Culture medium was renewed twice a week. All the experiments were performed with 3 to 5 week-old cultures. Depending on the used techniques; WB, IF or Q-PCR, cells were cultured in different culture plates with different cell concentration, according with the table 4.4, represented below.
Table 4.4 - Description of cells cultured in different plates with different concentrations, culture volumes for the different techniques.
Plates Number of plated cells per well Technique Volume per well (ml)
6 well 50*10
4
Western Blot
2,5
100*104 Q-PCR
24 well 35*104 Imunofluorence and flow
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
26
Role of CORM-A1 in primary culture of astrocytes
4.3
Inducers and inhibitors of cell death and autophagy
4.3.1
Starvation was used as a positive control for autophagy stimulation. Medium without supplementation of amino acids, Earle´s Balanced Salt Solution (EBSS) (Gibco Invitrogen, Germany) was added to cell culture for promoting starvation, by blocking nutrient supply.
In contrast, hydroxychloroquine (HCQ), which is an inhibitor of autophagy by blocking the autophagic flux, was used for evaluating it (figure 4.1). HCQ is attracted to the inside of organelles, preventing the fusion of lysosome with the autophagosome structure, by neutralizing the lysosomal pH, and at the same time inhibiting the action of lysossomal proteases. Thus, HCQ promotes the accumulation of the autophagy marker LC3I/II.
The levels of LC3-II are tightly correlated with the amount of autophagosomes and for that reason it is considered as the most reliable marker of active autophagosomes and autophagolysosomes (Yorimitsu; Klionsky et al. 2005). An increase of cellular LC3-II levels occurs during activation of autophagy, nevertheless accumulation of LC3-II can also be obtained by interrupting the autophagosome-lysosome fusion step or by inhibiting lysosome-mediated proteolysis. Therefore, the use of LC3-II as an autophagic marker needs to be complemented by an assay to estimate the overall autophagic flux, using HCQ. Therefore, measurement of autophagy activity includes (i) the increased synthesis or lipidation of LC3 or an increase in the formation of autophagosomes, and (ii) most importantly, the assessment of autophagic flux through the entire system, including lysosomes or the vacuole, and the subsequent release of the breakdown products (Klionsky, Abdalla et al. 2012). Thus, autophagic substrates need to be monitored dynamically over time to verify that they have reached the lysosome/vacuole, and, when appropriate, are degraded. For measuring the autophagic flux, HCQ is added to the cell culture for blocking the flux and LC3II levels were carried out by Western Blot analysis. In summary, autophagy is stimulated whenever an accumulation of LC3-II is obtained in the presence of HCQ (Klionsky, Abdalla et al. 2012)
.
Figure 4.1 - Autophagic Flux. a) Phagofore formation, b) Formation of a double-membrane named by autophagosome, with incorporation of damaged components, c) incorporation of lysosome and elimination of cargo. Adapted from (Hansen; Johansen et al. 2011)
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
27
Another inhibitor of autophagy used along this study was Wortmannin. Wortmannin is a cell-permeable, fungal metabolite that acts as a potent, selective and irreversible inhibitor of phosphatidylinositol 3-kinase (PI3K). PI3K is required for autophagy, thus inhibition of PI3K with wortmannin can prevent autophagic sequestration (at the beginning of the process) (Klionsky, Abdallaet al. 2012).
Cell death is induced with tert-Butylhydroperoxide (t-BHP), which is an organic peroxide widely used in a variety of oxidation processes. It is a pro-oxidant molecule, causing oxidative stress and resulting in cell or organ injury. Exposure to t-BHP promotes cell death, increasing ROS production and mitochondrial dysfunction. Cell viability was measure by flow cytometry analysis (Queiroga, Almeida et
al. 2010).
Hence, for achieve this aim we used the following compounds described at table 4.5.
Table 4.5 - Inducers and Inhibitors of autophagy and cell death inducers;
Compounds Action Final Concentration
tert-Butyl hydroperoxide
(T-TBS)
Pro-oxidant agent – cell death
inducer 0, 80, 160, 240, 280, 320µM EBSS Autophagy inducers starvation
Hydroxychloroquine
(HCQ) Autophagic flux blocker 3 mg/ml
Wortmannin
(WM) Autophagy inhibitor 100 nm
Induction of autophagy
4.3.2
Astrocytes, plated according to the table 4.4, were incubated with CORM-A1 at a final concentration of 12,5 and 25 µM with and without hydroxyclochroquine (HCQ) at 30µg/ml for 30 minutes or 1 hour at 37ºC, as described in figure 4.2 a).
Cells were collected for posterior analysis through Western Blot, Immunochemistry and Q-PCR in order to evaluate autophagy induction, mitophagy and mitochondria population, respectively. These assays were prepared accordingly to figure 4.2.
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
28
Figure 4.2- Scheme of autophagy induction in primary culture of astrocytes along time. Cells were treated with CORM-A1 (a) for assessing autophagic flux HCQ were also added and LC3 analysis by Western Blot, b) for measuring specific mitophagy stimulation, a mitochondrial dye was also added for further analysis by imunofluorescence (IF), c) Mitochondrial population is followed along 24h after CORM-A1 treatment by mitochondrial DNA quantification via Q-PCR analysis.
Induction of cell death and assessment of cell viability
4.3.3
Astrocytes were incubated for 18h with t-BHP (80-320µM). Then, medium was collected, cells were washed once with PBS and trypsined. Supernatant medium, PBS and cell pellet were pulled together for incubating with the dyes: propidium iodide (PI) and 3,3’-dihexyloxacarbocyanine iodide (DiOC) for 30 minutes at 37ºC, followed by cytofluorometric analysis with FACS scan (BD FACSCalibour). DiOC is used for measuringmitochondrial potential and PI for assessing plasmatic membrane integrity, an indirect way for assessing cell viability.
For evaluating the cytoprotective role of CO, in particular CORM-A1, and the correlation with autophagy, astrocytes were pre-treated with wortmanin 100µM for 1 hour (for preventing autophagic
Carbon monoxide, autophagy and cytoprotection in response to cerebral ischemia
29
process), then astrocytes were treated with CORM-A1 at 12,5µM for 30 minutes and 1h, followed by cell death induction as described in figure 4.3:Figure 4.3 - Scheme of the primary culture of astrocytes treatment for cell viability measurement;
Results were assessed by FlowJo analysis, version 10.1. For the quantification of cell viability two gates were outlined limiting death cells and alive cells, for the death cells just cells marked with PI positive (PI+) and DiOC negative (DiOC-) were considered and for live cells just cells regarding to PI negative (PI-) and DiOC positive (DiOC +) was showed in figure 4.4.
Figure 4.4 – Example of the gates used to evaluate the cell viability; Primary culture of astrocytes treated with and without CORM-A1 and pro-oxidante agent, t-BHP.