Caracterização do perfil hematológico e dos parâmetros
bioquímicos do soro da cabra (Capra aegagrus hircus) e a
sua comparação com os de indivíduos da sub-família
Caprinae já caracterizados
Dissertação de Mestrado em Biologia Clínica Laboratorial
Patrícia Alexandra Sobrinho da Silva
Orientadora: Amélia Maria Lopes Dias da Silva Co-orientadora: Maria Isabel Ribeiro Dias
_____________________________ A Orientadora
(Professora Doutora Amélia Maria Lopes Dias da Silva)
_____________________________ A Co-Orientadora
(Professora Doutora Maria Isabel Ribeiro Dias)
_____________________________ A Directora do Curso de Mestrado
Dissertação apresentada na Universidade de Trás-os-Montes e Alto Douro, com vista à obtenção do Grau de Mestre em Biologia Clínica Laboratorial
AGRADECIMENTOS
Quero agradecer em primeiro lugar à minha orientadora de dissertação de Mestrado, a Professora Doutora Amélia Maria Lopes Dias da Silva, e à co-orientadora, a Professora Doutora Maria Isabel Ribeiro Dias, pela atenção, disponibilidade, empenho e dedicação demonstrados. Quero também agradecer todos os conhecimentos que me transmitiram ao longo do estágio.
Agradeço ao Laboratório de Patologia Clínica do Pioledo, especialmente ao Doutor Nuno Aguiar, pelo estágio disponibilizado na área de Hematologia e Bioquímica.
Como não poderia deixar de ser, quero também agradecer aos meus queridos pais e irmão pelo apoio incondicional em todos os sentidos e em todos os momentos, principalmente nos mais difíceis.
Por último, quero agradecer ao amor da minha vida, o Carlos, por estar sempre ao meu lado, apoiando-me nos momentos felizes e nos momentos difíceis. Aos meus amigos do coração, Alexandra Alves e Ricardo Sousa, agradeço o apoio que sempre me deram durante a minha jornada na universidade.
i
Índice
Resumo ... vii Abstract ... ix Lista de Abreviaturas ... xiCapítulo I – Introdução ... 1
1. O sangue: função e composição ... 2
1.1. Componentes celulares ... 3
1.1.1. Glóbulos vermelhos: função... 3
1.1.2. Glóbulos brancos: composição e função ... 3
1.1.3. Plaquetas ou trombócitos ... 5
1.2. Principais componentes séricos ... 6
1.2.1. Minerais ... 6
1.2.2. Proteínas, enzimas e hormonas ... 8
1.2.3. Electrólitos ... 8
1.2.4. Substâncias nitrogenadas... 9
1.2.5. Hidratos de carbono e lípidos ... 10
1.2.5.1. Glicose ... 10
1.2.5.2. Beta-hidroxibutirato ... 10
1.2.5.3. Colesterol e triglicerídeos ... 11
2. Raças Autóctones de Caprinos em Portugal ... 11
2.1. Raça Bravia ... 12
2.2. Raça Serrana ... 13
ii 2.2.2. Ecótipo da Serra ... 14 2.2.3. Ecótipo Ribatejana ... 15 2.2.4. Ecótipo Transmontana ... 15 2.3. Raça Charnequeira ... 16 2.4. Raça Serpentina ... 17 2.5. Raça Algarvia ... 18
3. Parâmetros hematológicos e bioquímicos em Caprinos ... 19
Objectivos ... 21
Capítulo II – Material e Métodos ... 22
1. Modelo experimental e manutenção dos animais ... 23
2. Colheita de sangue ... 24
3. Determinação dos parâmetros do hemograma ... 24
3.1. Determinação do Hemograma... 25
3.2. Determinação da Fórmula Leucocitária ... 25
4. Determinação dos parâmetros da hemostase ... 27
4.1. Tempo de Tromboplastina Parcial (TTP) ... 27
4.2. Tempo de Protrombina (TP) ... 28
5. Determinação da electroforese de proteínas ... 28
6. Determinação dos parâmetros de bioquímica sanguínea ... 29
6.1. Glicose ... 30
6.2. Triglicerídeos ... 31
6.3. Colesterol... 31
6.4. Aspartato aminotransferase (AST) ... 32
6.5. Alanina aminotransferase (ALT) ... 32
iii
6.7. Fosfatase alcalina (ALP) ... 33
6.8. Ureia ... 34 6.9. Creatinina ... 35 6.10. Cálcio ... 35 6.11. Fósforo ... 36 6.12. Magnésio ... 36 6.13. Cobre e Selénio ... 37
6.14. Sódio, Potássio e Cloro ... 37
Capítulo III – Resultados e Discussão... 39
1. Padronização dos valores hematológicos e bioquímicos da cabra Serrana ... 40
2. Estudo dos parâmetros hematológicos da cabra Serrana ... 40
2.1. Parâmetros relativos aos eritrócitos ... 41
2.2. Parâmetros relativos aos leucócitos ... 44
2.3. Parâmetros da hemostase ... 47
3. Estudo dos parâmetros da bioquímica do soro da cabra Serrana ... 48
3.1. Parâmetros bioquímicos gerais ... 49
3.2. Parâmetros bioquímicos: electrólitos do plasma... 53
3.3. Parâmetros bioquímicos: perfil de proteínas plasmáticas ... 56
Capítulo IV – Conclusão ... 58
Referências Bibliográficas ... 60
iv
Índice de Figuras
Figura 1. Componentes do sangue separados por gravidade.. ... 2
Figura 2. Representação dos glóbulos vermelhos.. ... 3
Figura 3. Leucócitos: (a) Neutrófilos, (b) Eosinófilos, (c) Basófilos, (d) Monócitos, (e) Linfócitos.. ... 5
Figura 4. Fotagrafia da cabra de raça Bravia. ... 13
Figura 5. Ecótipo Jarmelista. ... 14
Figura 6. Ecótipo da Serra. ... 15
Figura 7. Ecótipo Ribatejano... 15
Figura 8. Ecótipo Transmontano. ... 16
Figura 9. Raça Charnequeira. ... 17
Figura 10. Raça Serpentina. ... 18
Figura 11. Raça Algarvia. ... 19
Figura 12. Aparelho usado para a determinação do hemograma, Horiba ABX®. ... 25
Figura 13. Aparelho utilizado para a determinação da fórmula leucocitária, Cell-Dyn 4000®... 26
Figura 14. Ilustração do aparelho usado para determinação dos parâmetros da hemóstase, STA Compact CT ®. ... 27
Figura 15. Aparelho utilizado para a determinação da electroforese de proteínas, CapillarysTM2. ... 29
Figura 16. Aparelho usado para a determinação dos parâmetros de bioquímica sanguínea, Architect Ci8200. ... 30
v
Índice de Tabelas
Tabela 1. Composição da alimentação oferecida aos animais. Matéria seca (MS) e composição química do alimento. ... 23 Tabela 2. Parâmetros hematológicos relativos às células vermelhas, referentes à cabra Serrana e respectivos intervalos de referência para espécie caprina em geral. ... 41 Tabela 3. Comparação dos valores hematológicos relativos às células vermelhas obtidos para a cabra de raça Serrana com os valores obtidos para outras cabras descritas na literatura. ... 42 Tabela 4. Parâmetros hematológicos relativos às células brancas, referentes à cabra Serrana e respectivos intervalos de referência para a cabra em geral. ... 45 Tabela 5. Comparação dos valores hematológicos relativos aos leucócitos obtidos para a cabra de raça Serrana com os valores obtidos para outras cabras descritas na literatura. ... 46 Tabela 6. Parâmetros hematológicos relativos à hemostase, referentes à cabra
Serrana. ... 47 Tabela 7. Comparação dos valores obtidos dos parâmetros hematológicos relativos à hemostase, referentes à cabra Serrana e outras cabras citadas na literatura. ... 48 Tabela 8. Parâmetros da bioquímica referente à cabra Serrana e respectivos
intervalos de referência para a cabra em geral. ... 50 Tabela 9. Comparação dos parâmetros bioquímicos gerais da cabra de raça Serrana com outras cabras citadas na literatura. ... 52
vi Tabela 10. Parâmetros da bioquímica referente à cabra Serrana e respectivos
intervalos de referência para a cabra em geral. ... 54 Tabela 11. Comparação dos parâmetros bioquímicos relativos aos electrólitos da cabra de raça Serrana com outras cabras citadas na literatura. ... 55 Tabela 12. Parâmetros da bioquímica referente à cabra Serrana e respectivos
intervalos de referência para a cabra em geral. ... 56 Tabela 13. Comparação dos resultados obtidos para a raça Serrana e outras raças citadas na literatura. ... 57
vii
Resumo
Os parâmetros hematológicos e de bioquímica sanguínea geral, incluindo os electrólitos plasmáticos, foram determinados em doze cabras de raça Serrana (Capra
aegagrus hircus) clinicamente saudáveis, uma raça típica e autóctone da região de
Trás-os-Montes, bem ambientada ao seu meio ambiente, com objectivo de padronização destes parâmetros na raça em estudo.
Os valores hematológicos obtidos foram: eritrócitos 7,12 ± 1,5 x1012/L; hemoglobina 8,6 ± 1,3 g/dL; hematócrito 15,1 ± 2,1 % e leucócitos 14,1 ± 3,88 x109/L. O tempo de tromboplastina parcial, tempo de protrombina e taxa de protrombina foi 25,1 ± 3,27 s, 16,98 ± 0,800 s e 60,25 ± 4,90 %, respectivamente. Os valores dos parâmetros de bioquímica sanguínea geral foram: glicose 62,7 ± 9,78 mg/dL; triglicerídeos 9,92 ± 3,99 mg/dL; colesterol total 77,58 ± 13,2 mg/dL; AST 89,42 ± 15,5 UI/L; ALT 11,9 ± 3,90 UI/L; γ-GT 57,9 ± 12,1 UI/L; ALP 831,0 ± 649,3 UI/L; ureia 32,8 ± 10,7 mg/dL; creatinina 0,77 ± 0,079 mg/dL; cálcio total 8,41 ± 0,470 mg/dL; fósforo 5,59 ± 1,64 mg/dL; magnésio 2,00 ± 0,270 mg/dL; cobre 134,8 ± 18,43 μg/dL; selénio 48,5 ± 21,2 μg/dL; sódio 143,8 ± 1,90 mEq/L; potássio 3,98 ± 0,44 mEq/L e cloreto 104,8 ± 1,52 mEq/L. Em relação ao perfil electroforético os valores foram: proteínas totais 6,9 ± 0,76 g/dL; albumina 2,80 ± 0,381 g/dL; α-1-globulina 0,46 ± 0,064 g/dL; α-2-globulina 0,82 ± 0,060 g/dL; β-α-2-globulina 0,49 ± 0,061 g/dL e γ-α-2-globulina 2,31 ± 0,575 g/dL.
Os valores obtidos para a hemoglobina, VCM, neutrófilos, eosinófilos, monócitos, glicose, AST, ureia, fósforo inorgânico, sódio, potássio, cloreto, proteínas totais, albumina, α-2-globulina e γ-globulina, encontram-se dentro do intervalo de referência considerado normal para a espécie caprina em geral. Por outro lado, os valores obtidos para o HCM, CHCM, leucócitos, linfócitos, γ-GT e ALP situam-se acima do intervalo de referência considerado normal para a espécie caprina em geral e os valores obtidos para os eritrócitos, HTC, colesterol total, creatinina, cálcio total, magnésio e α-1-globulina encontraram-se abaixo do intervalo de referência considerado normal para a espécie caprina em geral. Não foi possível proceder à
viii
comparação dos valores obtidos para o cobre, selénio e β-globulina por não existirem valores de referência destes parâmetros na cabra.
Palavras chave: Bioquímica sanguínea, hematologia geral, padronização dos
ix
Abstract
The haematological and blood general biochemistry parameters, including serum electrolytes were determined in twelve clinically healthy Serrana breed goats (Capra hircus aegagrus), well adapted to its environment, a typical race indigenous to the region of Trás-os-Montes, aiming the standardization of this parameters in the breed.
Hematologic values obtained were: erythrocytes 7.12 ± 1.5 x1012/L, hemoglobin 8.6 ± 1.3 g/dL, hematocrit 15.1 ± 2.1% and leukocytes 14.1 ± 3.88 x109/L. The time of parcial thromboplastin and prothrombin and the rate of prothrombin was 25.1 ± 3.27 s, 16.98 ± 0.800 s and 60.25 ± 4.90%, respectively. The values of blood general biochemistries parameters were: glucose 62.7 ± 9.78 mg/dL; triglycerides 9.92 ± 3.99 mg/dL; total cholesterol 77.58 ± 13,2 mg /dL; AST 89.42 ± 15.5 IU/L; ALT 11.9 ± 3.90 IU/L; γ-GT 57.9 ± 12.1 IU/L, alkaline phosphatase 831.0 ± 649.3 IU/L, urea 32.8 ± 10.7 mg/dL; creatinine 0.77 ± 0.079 mg/dL; total calcium 8.41 ± 0.470 mg/dL; phosphorus 5.59 ± 1.64 mg/dL; magnesium 2.00 ± 0.270 mg/dL; copper 134.8 ± 18.43 mg/dL; selenium 48.5 ± 21.2 mg/dL; sodium 143.8 ± 1.90 mEq/L, potassium 3.98 ± 0.44 mEq/L and chloride 104.8 ± 1.52 mEq/L. Regarding to the electrophoretic protein profile the values were: total protein 6.9 ± 0.76 g/dL; albumin 2.80 ± 0.381 g/dL; α-1-globulin 0.46 ± 0.064 g/dL; α-2-globulin 0.82 ± 0.060 g/dL; β-globulin 0.49 ± 0.061 g/dL and γ-globulin 2.31 ± 0.575 g/dL.
The values obtained for the hemoglobin, MCV, neutrophils, eosinophils, monocytes, glucose, AST, urea, inorganic phosphorus, sodium, potassium, chloride, total protein, albumin, α-2-globulin, and γ-globulin were found within the reference range considered normal for the goat specie in general. Moreover, the values obtained for the MCH, MCHC, leukocytes, lymphocytes, γ-GT and ALP were situated above the normal reference range for the goat specie in general, and the values for the erythrocytes, HTC, total cholesterol, creatinine, total calcium, magnesium, and α-1-globulin were below the reference range considered normal for the goat specie in general. We were unable to make a comparison for the values of copper, selenium and
x
β-globulin, because there are no references values published for these parameters for goat specie.
Keywords: Blood biochemistry, general hematology, standardization of biochemical
xi
Lista de Abreviaturas
ADP – Adenosina difosfato ALP – Fosfatase alcalina
ALT – Alanina aminotransferase AOAC – Official methods of analysis AST – Aspartato aminotransferase ATP – Adenosina trifosfato
Ca – Cálcio
CHCM – Concentração de hemoglobina corpuscular média Cl – Cloreto
Co – Cobalto
Co2 – Dióxido de carbono
Cu – Cobre
DNA – Ácido Dexosirribonucleico DP – Desvio padrão
ECF – Fluído Extracelular
EDTA – Ácido etilenodiamino tetra-acético ELF – Fluído extracelular
FND – Fibras neutras de detergentes G-6-P – Glicose-6-fosfato
xii H2O2 – Peróxido de hidrogénio
Hb – Hemoglobina
HCM – Hemoglobina corpuscular média
HEDTA – N-(2-hideroxietl)etilenodiaminotriacetic acid HTC – Hematócrito
ICF – Fluído Intracelular
IDE – Índice de distribuição eritrocitária K – Potássio
Mg – Magnésio MS – Matéria Seca Na – Sódio
NADH/NAD+ – Nicotinamida adenina dinucleótido
NCR – Nutrient Requerimento of Sheep O2 - Oxigénio
P – Fósforo
PC – Proteínas cruas
PMN – Neutrófilos polimorfonucleares RNA – Ácido Ribonucleico
Se – Selénio
SNC – Sistema nervosa central TP – Tempo de Protrombina
xiii TTP – Tempo de tromboplastina parcial
VCM – Volume corpuscular médio γ-GT – γ-Glutamil transferase
2
1. O sangue: função e composição
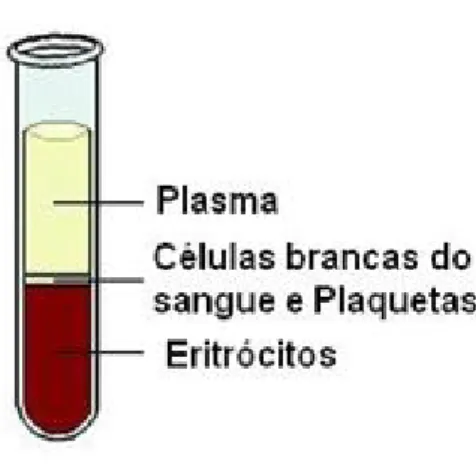
O sangue é um fluido formado por células sanguíneas (glóbulos vermelhos, glóbulos brancos e plaquetas) e plasma, constituindo cerca de 70 mL/kg do peso corporal. O sangue exerce uma grande variedade de funções, tais como o transporte de gases e nutrientes, actua como tampão, estabiliza a temperatura corporal e defende o organismo contra agentes patológicos. Para além disso, os componentes do sangue são capazes de evitar a perda de líquidos da circulação como resultado de ferimentos através do processo de coagulação (Ashton, 2010). Os componentes do sangue separam-se por gravidade, evidenciando-se três camadas: i) a mais densa, composta por glóbulos vermelhos (≈ 40 % do total, no humano e ≈ 56 % na cabra (Smith e Sherman, 2009)), ii) uma camada intermédia, composta pelos glóbulos brancos e plaquetas (uma reduzida percentagem), e iii) a superior, de cor amarelo-pálido, que se designa por plasma, é composta maioritariamente por água, proteínas (como por exemplo albumina, enzimas, anticorpos), açúcares, entre outros. O plasma constitui cerca de 60 % do total (nos humanos), verFigura 1 (Dean, 2005).
3
1.1.
Componentes celulares
1.1.1. Glóbulos vermelhos: função
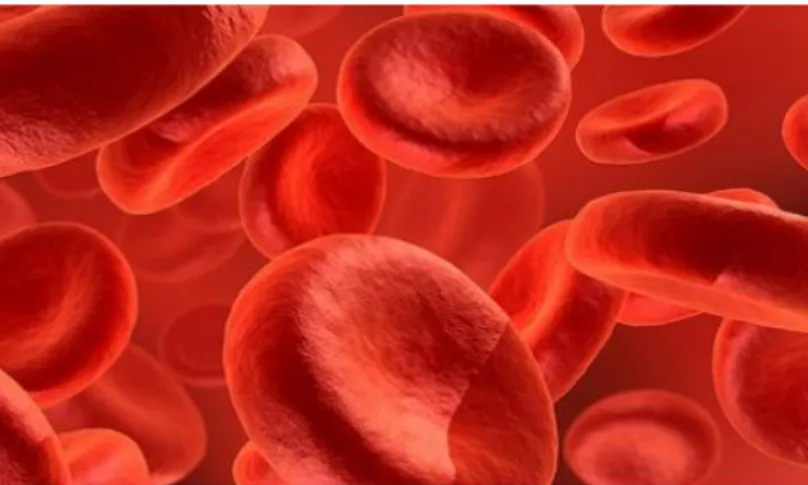
Os glóbulos vermelhos (Figura 2), também conhecidos por eritrócitos ou hemácias, têm como principal função o transporte de oxigénio dos pulmões para os tecidos, e o transporte de dióxido de carbono (CO2) dos tecidos para os pulmões. A
hemoglobina (Hb) é a proteína maioritária dos eritrócitos, é uma metaloproteína que contém ferro na sua composição. A Hb é a responsável pelo transporte de O2 (HbO2) e
de uma parte do CO2 (Ashton, 2010). Nos mamíferos, os glóbulos vermelhos quando
maduros são células sem núcleo, são produzidos na medula óssea à taxa de 2 - 3 milhões por segundo e têm um tempo médio de vida de 100 - 120 dias.
Figura 2. Representação dos glóbulos vermelhos (figura retirada de http://noticias.up.pt/catalogo_noticias.php?ID=4907).
1.1.2. Glóbulos brancos: composição e função
Os glóbulos brancos ou leucócitos (Figura 3) são células produzidas pela medula óssea, são menos abundantes no sangue que os eritrócitos, não possuem hemoglobina mas contêm núcleo e outros organelos. Os leucócitos têm como função defender o organismo contra agentes patológicos e remover células danificadas, toxinas e outras substâncias residuais. Os leucócitos são geralmente divididos em granulócitos (neutrófilos, eosinófilos e basófilos) e agranulócitos (monócitos e linfócitos), de acordo com a presença ou ausência, respectivamente, de grânulos no citoplasma (Alberts,
4
2005). Os granulócitos também são conhecidos por células polimorfonucleares, devido a apresentarem núcleos segmentados. Os granulócitos são produzidos na medula óssea (Bergman, 1999).
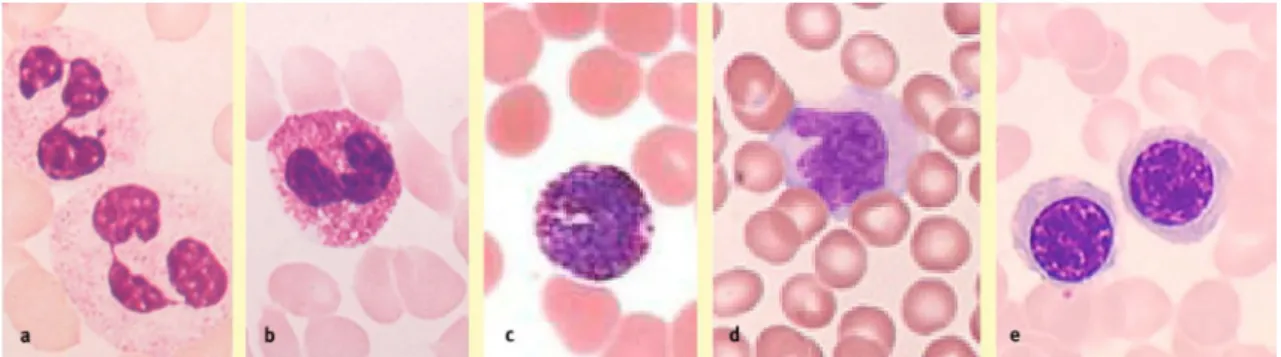
Os neutrófilos (Figura 3a), também conhecidos como neutrófilos polimorfonucleares (PMN), devido à estrutura lobular do seu núcleo, são a forma mais comum dos leucócitos, representando 50 - 70 % dos glóbulos brancos circulantes. Os neutrófilos têm uma elevada mobilidade e têm como principal função a fagocitose e destruição de bactérias ou fungos (Gordon-Smith, 2009; Ashton, 2010).
Os eosinófilos (também designados por acidófilos) (Figura 3b) correspondem a 2 - 4 % dos leucócitos circulantes e são reconhecidos por possuírem dois lóbulos no núcleo e abundantes grânulos vermelhos no citoplasma após coloração com eosina (um corante vermelho ácido). Os eosinófilos aumentam drasticamente em reacções parasitárias e em processos alérgicos, como por exemplo asma (Gordon-Smith, 2009; Ashton, 2010).
Os basófilos (Figura 3c) são raros, correspondendo apenas a 1 % dos leucócitos. Estes têm como principal modo de acção a libertação de histamina e heparina nos locais de lesão. A histamina provoca a vasodilatação e a heparina evita a coagulação do sangue (Gordon-Smith, 2009; Ashton, 2010).
Os monócitos (Figura 3d) correspondem a 2 - 8 % dos leucócitos e são distinguidos pelo seu núcleo oval ou em forma de rim. Os monócitos passam um curto período de tempo na circulação antes de se deslocarem para os tecidos, onde são denominados de macrófagos (principais células fagocíticas). Os monócitos aumentam o seu número em situações infecciosas e inflamatórias crónicas (Gordon-Smith, 2009; Ashton, 2010). Os monócitos, quando deixam a corrente sanguínea e migram para os tecidos, transformam-se em macrófagos (Dean, 2005).
Os linfócitos (Figura 3e) possuem um núcleo grande e redondo, rodeado de pouco citoplasma e corresponde a 20 - 30 % dos leucócitos circulantes, mas apresentam um tempo de vida muito curto em circulação. Os linfócitos são responsáveis pela especificidade nas respostas imunitárias. Os linfócitos podem ser
5
divididos em três grupos (ou tipos): i) células T, ii) células B e iii) células natural killer (NK), cada uma com uma função específica. Os linfócitos T são as células principais da imunidade celular. Funcionalmente, existem três subtipos de linfócitos T. Os linfócitos T citotóxicos (células Tc) desempenham funções como a destruição de células infectadas por vírus ou células neoplásicas. Os linfócitos T auxiliares (células Th) ajudam as outras células a desempenharem as suas funções, nomeadamente os linfócitos B a produzirem anticorpos e os macrófagos nas suas actividades microbicidas. Os linfócitos T supressores (células Ts) inibem as células T e activam as células B, moderando a resposta imune. Os linfócitos B são responsáveis pela imunidade humoral, isto é, as células B activadas diferenciam-se em plasmócitos que produzem anticorpos em resposta a antigénios estranhos (Minors, 2004).
Os linfócitos NK são mediadores da resposta imunitária inata e estão implicados em várias actividades como a destruição de células tumorais, na resistência a infecções virais, na regulação da hematopoiese e em fenómenos de rejeição de órgãos transplantados (Minors, 2004).
Figura 3. Leucócitos: (a) Neutrófilos, (b) Eosinófilos, (c) Basófilos, (d) Monócitos, (e) Linfócitos (Figura adaptada de Gordon-Smith, 2009).
1.1.3. Plaquetas ou trombócitos
As plaquetas sanguíneas ou trombócitos são partículas ou fragmentos celulares (sem núcleo) de megacariócitos que circulam no sangue e a sua principal função é a de formação de um coágulo durante o processo de coagulação sanguínea. A sua intervenção no processo de coagulação é um processo físico, por alteração de forma, e
6
químico, pela libertação de vários factores que fazem parte do seu conteúdo. Do seu conteúdo destaca-se RNA, mitocôndrias, vários tipos de grânulos como os grânulos alfa (contendo o factor de von Willebrand, fibrinogénio, factor V) e corpos densos (contendo ATP, adenosina, serotonina, histamina, cálcio) (Kaneko et al., 1997). Estas têm uma duração média de 5 dias.
1.2. Principais componentes séricos
O plasma sanguíneo, ou soro, é o que sustenta as células sanguíneas. O plasma sanguíneo contém gases, proteínas, hidratos de carbono, aminoácidos, lípidos, sais inorgânicos (ou minerais), enzimas, hormonas, entre outros.
1.2.1. Minerais
O cálcio (Ca), o sódio (Na), o potássio (K) e o fósforo (P) são os minerais que apresentam grande importância de diagnóstico, em todas as espécies animais, enquanto o magnésio (Mg), o cobre (Cu), o cobalto (Co) e o selénio (Se) são mais importantes em ruminantes (Kerr, 1989).
O cálcio é o mineral mais abundante, a sua maioria encontra-se no tecido ósseo e a restante porção desempenha um papel importante em vários processos biológicos fundamentais. É um mineral que desempenha uma função central na manutenção da hemostase dos animais vertebrados, incluindo na contracção muscular, na coagulação do sangue, regulando a actividade enzimática, na excitabilidade neuronal, na secreção de hormonas e na adesão celular. Para além disso, é um componente estrutural essencial do esqueleto (Kaneko et al., 1997).
Nos mamíferos, o fósforo e o cálcio apresentam-se predominantemente na forma de hidroxiapatite [Ca10(PO4)6(OH)2] na matriz mineralizada do osso, que
representa um depósito de fósforo (80 - 90 %) e de cálcio (95 %). A restante parte do fósforo encontra-se a nível intracelular. O fosfato (PO43-) é o principal anião que existe
7
nucleicos, fosfoproteínas, ATP, entre outros) ou inorgânica e desempenha uma função integral em vários processos metabólicos (Kaneko et al., 1997).
O magnésio é necessário à actividade enzimática, em particular as enzimas que usam o complexo ATP-Mg2+. O magnésio desempenha uma função essencial na contracção muscular, no metabolismo das proteínas, ácidos gordos e hidratos de carbono, na transferência de grupos metilo, na fosforilação oxidativa, nas propriedades funcionais e estabilização das membranas, na divisão celular e nas respostas imunes. Além disso, o Mg regula a permeabilidade da membrana mitocondrial e a estrutura do RNA ribossómico e do DNA; e é necessário para a manutenção dos níveis normais de potássio na célula (Kaneko et al., 1997).
No plasma, o cobre encontra-se maioritariamente (80 - 95 %) associado a uma proteína, a ceruplasmina, que também o transporta aos tecidos (Hauschild et al., 2008). O cobre desempenha um papel importante em vários processos biológicos (como por exemplo no metabolismo energético, na hemostase de ferro e em mecanismos de protecção antioxidante). O cobre desempenha um papel importante no transporte de ferro (Kerr, 1989), oxidando-o para facilitar a sua ligação à transferrina (Chaney, 2006). O cobre é um cofactor de algumas enzimas com elevada relevância (e.g., citocromo C oxidase, dopamina-β-oxidase).
O cobalto é importante para a o metabolismo da vitamina B12 (Kerr, 1989),
fazendo parte desta, daí que esta também se designa por cobalamina. Actua como cofactor de várias enzimas (Chaney, 2006). Nos ruminantes, as bactérias do rúmen usam o cobalto da alimentação para sintetizar vitamina B12 que actua como co-enzima
nas vias de síntese de propionato, o principal precursor de glicose, e de alguns aminoácidos.
O selénio encontra-se associado a selenoproteínas, como por exemplo a glutationa peroxidase e a selenoproteína W muscular. A glutationa peroxidase destrói peróxidos no citoplasma, efeito que complementa a acção da vitamina E. A selenoproteína W muscular é importante para o correcto funcionamento tanto do músculo esquelético como do cardíaco (Kerr, 1989).
8 1.2.2. Proteínas, enzimas e hormonas
As funções das proteínas no organismo são inúmeras, incluindo as presentes no plasma. Mantêm a pressão coloidosmótica; são catalisadoras (enzimas) em reacções bioquímicas; actuam como tampão para a manutenção do equilíbrio ácido-base; são reguladoras (hormonas); participam na coagulação sanguínea e na defesa do organismo (anticorpos); são nutrientes; são transportadas e transportam componentes para a maioria dos constituintes do plasma (Kaneko et al., 1997).
No plasma, as principais classes de proteínas, separadas de acordo com o tamanho e a carga, são as albuminas plasmáticas, α-1-globulinas, α-2-globulinas, β-globulinas e γ-β-globulinas.
1.2.3. Electrólitos
Os electrólitos são substâncias que existem como partículas com carga positiva (catiões) ou negativa (aniões) em solução aquosa (Kaneko et al., 1997). O ião sódio (Na+), potássio (K+) e cloreto (Cl-) são os electrólitos de maior relevância em medicina veterinária (Kerr, 1989).
O ião sódio (Na+) é o electrólito que está intimamente associado com o
equilíbrio osmótico e a maioria dos distúrbios tendem a ser primariamente problemas de fluido. Os sintomas estão normalmente relacionados com alterações no volume dos fluidos dos compartimentos corporais, particularmente o volume do compartimento intracelular em relação ao volume do compartimento extracelular (ICF/ECF) no SNC (Kerr, 1989). O Na+ é um ião essencialmente extracelular muito importante na excitabilidade celular.
O ião potássio (K+) é essencialmente um ião intracelular e a sua concentração
no ECF é baixa. Tem menor participação no equilíbrio osmótico que o sódio e a maioria dos distúrbios são directamente relacionados com a perda excessiva ou falta de excreção deste, independentemente do estado de hidratação. Os sintomas estão normalmente relacionados com a diminuição da actividade eléctrica, particularmente
9
no coração (Kerr, 1989), dado que é um regulador do potencial de repouso de células excitáveis.
O ião cloreto (Cl-) tende a ser o electrólito menos considerado, mas pode
oferecer informações muito úteis. Como anião, a sua concentração é afectada pela concentração de outros aniões, como o bicarbonato. Isso significa que pacientes com acidose e com baixas concentrações de bicarbonato, o cloreto encontra-se geralmente elevado, enquanto em paciente alcalóticos com elevada concentração de bicarbonato, o cloreto está geralmente baixo. Na ausência do equilíbrio ácido/base significativas, a concentração plasmática de cloreto é geralmente paralela à do ião sódio (Kerr, 1989).
O dióxido de carbono (CO2) total é o único indicador de relevância directa da
condição ácido/base. A maior parte do CO2 total do plasma está na forma de
bicarbonato (HCO3-). O uso deste indicador tem as suas limitações, mas pode ser extremamente útil em pacientes com distúrbios metabólicos de ácido/base. Neste caso, o CO2 total é suficiente quer para demonstrar a presença de acidose ou alcalose
quer para elucidar quanto à sua gravidade (Kerr, 1989).
1.2.4. Substâncias nitrogenadas
A ureia é um produto nitrogenado que é produzido no fígado como produto final do catabolismo dos aminoácidos. Depois de formada no fígado, a ureia é transportada através do plasma para os rins, onde é excretada para a urina (Kerr, 1989). A elevação dos níveis desta pode estar associada a estados de desidratação, em ruminantes (Kaneko et al., 1997; Coomes, 2006)
A creatinina é, assim como a ureia, um composto nitrogenado formado a partir da degradação da creatina. Os níveis de creatinina no plasma dependem da massa muscular e da actividade física. Nos ruminantes, alteração nos níveis de creatinina são indicadores da função renal, melhores que os níveis de ureia (Kerr, 1989; Kaneko et al., 1997; Chizzotti et al., 2008).
10 1.2.5. Hidratos de carbono e lípidos
1.2.5.1. Glicose
A glicose é fornecida pela absorção intestinal proveniente da dieta e pela produção hepática de glicose a partir dos seus precursores, tais como hidratos de carbono (glicogénio, frutose, galactose) e aminoácidos (gliconeogénese) (Kaneko et al., 1997).
Na fase pós-absortiva, a glicose é transportada do local de absorção no intestino para os locais de armazenamento de glicogénio, principalmente o fígado e músculos. Em jejum, a concentração plasmática de glicose nos animais é mantida pela mobilização de hidratos de carbono a partir de qualquer forma que possa ser metabolizada – normalmente predomina a glicogenólise, com alguma lipólise e gliconeogénese, mas se o jejum prolongar a lipólise e mesmo a proteinólise tornam-se mais importantes (Berg et al., 2002).
1.2.5.2. Beta-hidroxibutirato
Os corpos cetónicos, que são compostos por ácido acético, β-hidroxibutirato (também conhecido como ácido 3-hidroxibutírico) e acetona são importantes no metabolismo dos animais e a sua principal origem são os ácidos gordos, quer esterificados como não esterificados (Kaneko et al., 1997).
A cetose desenvolve-se devido à deficiência na passagem de glicose através da via glicolítica nas células. A falta de produtos da glicólise impede o funcionamento do ciclo de Krebs. Simultaneamente, os ácidos gordos são utilizados como fonte de energia alternativa e os corpos cetónicos são formados como produtos anormais do catabolismo dos ácidos gordos devido ao bloqueio do ciclo de Krebs (Kerr, 1989). Variações na concentração plasmática de β-hidroxibutirato indicam alterações metabólicas dos glúcidos e/ou dos lípidos.
11 1.2.5.3. Colesterol e triglicerídeos
O colesterol é um componente das membranas celulares, a par com outros lípidos, regulando a fluidez membranar. O excesso de colesterol é eliminado na bílis, parcialmente como ácido biliar e sais biliares e algum como colesterol inalterado, que pode ser reabsorvido (Kerr, 1989).
O armazenamento do excesso de energia em gordura é importante dado que a mobilização desta em períodos de baixo balanço energético é fundamental. A gordura é armazenada como triglicerídeos, constituídos pela ligação de três resíduos de ácidos gordos esterificados a um glicerol. A mobilização da gordura normal é estimulada pela adrenalina e envolve a actividade das lipases e esterases dentro dos depósitos de gordura para a divisão dos ácidos gordos, resultando na libertação de ácidos gordos livres e glicerol para o plasma. A concentração plasmática dos triglicerídeos, que é normalmente baixa, não é afectada pela lipólise (Kerr, 1989), mas, por outro lado, em parte, reflecte o tipo de alimentação.
Alterações nos níveis de colesterol e/ou de triglicerídeos podem denotar distúrbios alimentares, metabólicos e/ou hormonais.
2. Raças Autóctones de Caprinos em Portugal
Segundo García-Perea e Gisbert (1997), a cabra doméstica situa-se na escala zoológica da seguinte forma:
Classe: Mammalia; Sub-classe: Theria; Infra-classe: Eutheria; Coorte: Ferungulata; Super-ordem: Praxonia; Ordem: Artiodactyla; Sub-ordem: Ruminantia; Infra-ordem: Pércora;
12
Família: Bovidae; Sub-família: Caprinae;
Espécie: Capra hircus (Linneaeus, 1758).
Em Portugal estão registadas cinco raças autóctones de caprinos, desde 1987, as quais são designadas por Raça Bravia, Raça Serrana (ecótipos Jarmelista, da Serra, Ribatejana e Transmontana), Raça Serpentina, Raça Algarvia e Raça Charnequeira. A descrição a seguir apresentada, relativamente às raças autóctones caprinas, foi retirada das publicações da Sociedade Portuguesa de Ovinotecnia e Caprinotecnia (www.ovinosecaprinos.com), Associação Nacional de Caprinicultores da raça Serrana (www.ancras.pt) e Recursos Genéticos (DGP, 1987).
2.1.
Raça Bravia
A Raça Bravia, também conhecida como Brava ou Cabra da Serra, é assim designada devido ao domínio selvagem em que é criada (Figura 4).
Este grupo racial esteve incluído na raça Charnequeira, ao qual se atribuiu a mesma origem filogenética. Ambas as raças podem ter tido uma origem comum, mas as condições geoclimáticas, socioeconómicas e culturais muito diferentes, além de outras interferências, foram determinantes para moldar animais com atributos zootécnicos distintos.
Os animais desta raça caracterizam-se pelo seu pequeno porte, grande agilidade e pela sua pelagem curta, a qual tem como cores predominantes o castanho e preto, apresentando-se esta última tonalidade mais escura na cabeça, ventre e extremidades dos membros, ou mais clara no ventre. O pêlo é curto nas fêmeas e mais comprido e áspero nos machos. A cabeça é triangular, com cornos em ambos os sexos, nas fêmeas são finos, erectos ou ligeiramente encurvados para trás, enquanto nos machos os cornos são ligeiramente espiralados para fora e para cima. As orelhas, em ambos os sexos, apresentam tamanho médio, horizontais para a frente. O pescoço é comprido, fino e bem ligado. Geralmente, o tronco destes animais é pouco desenvolvido, com uma linha dorso-lombar recta, uma garupa descaída e diâmetro
13
dorso-esternal e bicostal pequeno. Os membros são curtos e finos, com articulações bem salientes, unhas resistentes e aprumos moldados para trepar e saltar.
Em 2011, esta raça é representada por 9.800 animais inscritos no Registo Zootécnico (www.ovinosecaprinos.com), difundidos pelas seguintes zonas geográficas: Arcos de Valdevez, Ponte da Barca, Terras de Bouro, Ribeira de Pena, Vila Real, Vila Pouca de Aguiar e Mondim de Bastos. Algumas destas zonas encontram-se inseridas em Parques Naturais, nomeadamente no da Peneda Gerês e no do Alvão.
Figura 4. Fotografia da cabra de raça Bravia (figura retirada de www.ovinosecaprinos.com).
2.2.
Raça Serrana
A raça Serrana tem origem na Serra da Estrela e, actualmente, com maior expressão em Portugal, predominando nas zonas a norte do Tejo, com excepção do distrito de Castelo Branco e da península de Setúbal. Em 2011, estavam registados no livro genealógico 19.338 cabras de raça Serrana (www.ovinosecaprinos.com).
A pelagem da cabra Serrana pode apresentar diversas cores, tais como preta, castanha ou ruça, podendo ainda apresentar coloração amarela em algumas regiões. A cabeça é grande, comprida, de perfil subcôncavo, com fronte ampla e ligeiramente abaulada, possui face triangular, chanfro largo, rectilíneo e com uma depressão na união frontal. As orelhas são relativamente curtas e horizontais e o focinho fino. Os
14
cornos são de secção triangular, rugosos, dirigidos para trás, com hastes paralelos ou divergentes, ou ligeiramente dirigidas para trás, divergentes ou espiraladas. O pescoço é comprido e mal musculado. O tronco é de corpulência média. A linha dorso lombar é oblíqua de trás para a frente, o dorso e os rins são descarnados, mas rectilíneos e a garupa é descaída. Possuem cauda curta e arrebitada. Os membros são finos e resistentes, com unhas pequenas e muito duras.
Nesta raça podem distinguir-se quatro ecótipos: Jarmelista (Figura 5), da Serra (Figura 6), Ribatejana (Figura 7) e Transmontana (Figura 8). Estes ecótipos resultam das diferenças do meio em que vivem (DGP, 1987).
2.2.1. Ecótipo Jarmelista
Estes animais estão bem adaptados às condições edafo-climáticasdas zonas de montanhas da Guarda, Pinhel, Almeida e Sabugal. Estes animais são de aptidão dupla, produção de leite e carne.
Figura 5. Ecótipo Jarmelista (figura retirada de www.ancras.pt).
2.2.2. Ecótipo da Serra
Estes animais habitam a zona da Serra da Estrela, nomeadamente a Guarda, Manteigas e Covilhã.
15
Figura 6. Ecótipo da Serra (figura retirada de www.ancras.pt).
2.2.3. Ecótipo Ribatejana
Estes animais habitam as zonas do Ribatejo, estendendo-se para o Oeste e são, tal como o ecótipo Jarmelista, bons produtores de leite.
Figura 7. Ecótipo Ribatejano (figura retirada de www.ancras.pt).
2.2.4. Ecótipo Transmontana
Estes animais habitam a região de Trás-os-Montes, apresentando uma produção de leite de ≈1/2 do ecótipo Ribatejano e Jarmelista.
16
Figura 8. Ecótipo Transmontano (figura retirada de www.ancras.pt).
2.3.
Raça Charnequeira
A raça Charnequeira parece proceder da cabra Aegagrus, tendo mais tarde recebido influência do tronco Pirenaico. Porém, relativamente a este ponto existem outras opiniões, que dizem ser a Charnequeira descendente da cabra Falconeri ou da sua representante na Europa, a cabra palustre de Reitimageri, Capra hircus
sterpsicerus ou Céltica de Augusto. O nome de Charnequeira resulta do facto da maior
parte destes animais ser explorado na “Charneca”. Em 2009, havia 4.881 cabras de caça Charnequeira inscritas no livro genealógico (www.ovinosecaprinos.com).
Os animais desta raça possuem pelagem de cor vermelho claro ou mais escuro até castanho carregado, pêlo liso, curto e por vezes brilhante (Figura 9). A cabeça da cabra Charnequeira possui tamanho médio, de perfil rectilíneo a subcôncavo, de fronte convexa, seguida de uma pequena depressão, e de chanfro rectilíneo; as orelhas pouco destacadas, direitas e de comprimento médio. Os cornos, quando presentes, são grandes, largos e juntos na base, dirigidos para cima e ligeiramente inclinados para trás, divergentes, retorcidos nas pontas ou nitidamente espiralados, rugosos e de secção triangular; barba frequente nos bodes e rara nas fêmeas. O pescoço é comprido e estreito. O tronco é amplo, com peito estreito e profundo. A cauda é curta, horizontal e arrebitada na ponta; abdómen regularmente desenvolvido. Os membros são fortes, curtos, com aprumos regulares e unhas resistentes (DGP, 1987).
17
Figura 9. Raça Charnequeira (figura retirada de www.ovinosecaprinos.com).
2.4.
Raça Serpentina
A origem filogénica da raça Serpentina não é conhecida, mas pensa-se que tenha resulta de animais provenientes de Espanha, posteriormente seleccionados até atingirem o aspecto actual e difusão por todo o Alentejo. A sua denominação tem sofrido algumas alterações de acordo com as deslocações e a expansão ocorridas. Inicialmente, era denominada de Cabra Espanhola ou Castelhana, uma vez que os primeiros efectivos eram originários da Espanha, e, posteriormente, Raiana. Finalmente, após as características actuais bem fixadas, de que resultam as particularidades da sua pelagem, conquistou a denominação de Serpentina (também conhecida por Alentejana ou Portuguesa). No ano de 2011, estavam inscritas no livro genealógico 4.162 cabras da raça serpentina (www.ovinosecaprinos.com).
A cabra Serpentina possui uma pelagem branca ou creme, com um listão preto no dorso (Figura 10). O ventre é preto, assim como a parte interna das orelhas, o focinho e a extremidade dos membros. A cabeça é grande, de tipo longilíneo; a fronte é larga e ligeiramente convexa; o chanfro é rectilíneo; as orelhas são grandes e semi-pendentes; a barba existe em ambos os sexos, mas é mais reduzida nas fêmeas. Os cornos são largos e juntos na base, dirigidos para cima e para trás, divergentes nas
18
extremidades e sensivelmente espiralados. O pescoço é médio e bem musculado, tendo grande desenvolvimento no terço anterior. O tronco é bem desenvolvido. Os membros são fortes, compridos e com articulações volumosas e secas. As unhas são de tamanho médio, duras, com boa base de apoio.
Figura 10. Raça Serpentina (figura retirada de www.ovinosecaprinos.com).
2.5.
Raça Algarvia
A raça Algarvia deve o seu nome ao facto de ter o seu berço no Algarve. A sua origem filogenética é desconhecida. No entanto, pensa-se que tenha resultado da Charnequeira e da penetração de genes de cabras exóticas que aumentaram o seu potencial leiteiro e contribuíram para a sua pelagem característica.
No ano de 2011, estavam inscritas 4.668 cabras de raça Algarvia no livro genealógico (www.ovinosecaprinos.com).
Os caprinos da raça Algarvia possuem pêlo curto, de cor branca, com malhas castanhas de várias tonalidades ou pretas, que podem ser bem definidas ou disseminar-se irregularmente por todo o corpo (Figura 11). A cabeça é de dimensão regular com chanfro rectilíneo ou ligeiramente côncavo. As orelhas são de quatro tipos díspares: compridas, abertas e pendentes; médias, em “corneto” e lançadas para fora; curtas, em “corneto” e direitas e muito curtas, com cerca de 3 a 4 cm de comprimento e erectas. Em ambos os sexos, os cornos são geralmente largos na base, dirigidos para cima, para trás e para os lados e espiralados. O pescoço apresenta comprimento regular e bem musculado. O tronco é bem desenvolvido, com linha dorsal quase
19
horizontal, abdómen volumoso, garupa descaída e cauda pequena. Os membros são fortes e aprumados. Esta raça encontra-se muito bem adaptada a zonas pobres como as que podemos encontrar na Serra e Barrocal do Algarve.
Figura 11. Raça Algarvia (figura retirada de www.ovinosecaprinos.com).
3. Parâmetros hematológicos e bioquímicos em Caprinos
A Cabra (Capra aegagrus hircus) é uma raça típica e autóctone da região de Trás-os-Montes, estando bem ambientada ao seu meio ambiente. Nos últimos anos, a cabra tem sido utilizada como modelo animal em investigação em Ortopedia, pelo facto dos pequenos ruminantes serem animais de grande porte, que possuem um sistema esquelético com as fases de crescimento e adulta bem diferenciadas, após o encerramento das placas epifásicas de crescimento, e uma marcada resposta de remodelação Haversiana (An e Friedman, 1999). Aquando a transladação de informação científica entre espécies (neste caso da cabra para o Homem) devemos ter o máximo de informação acessória para o melhor enquadramento.
A caracterização do perfil hematológico e dos parâmetros bioquímicos permite, não só caracterizar a raça, como, comparando com outros indivíduos da mesma espécie, avaliar sobre o estado de saúde, e também, na transladação para o Homem, avaliar sobre o ambiente interno e a importância de factores
20
hematológicos/bioquímicos a ter em conta quando se colhem órgãos ou parte deles para estudos em bioengenharia de tecidos.
A composição bioquímica sanguínea reflecte com precisão a situação metabólica dos tecidos animais, de forma a poder avaliar lesões teciduais, transtornos no funcionamento de órgãos, adaptação do animal diante desafios nutricionais, fisiológicos e desequilíbrios metabólicos específicos (Araújo e Silva, 2008).
Os componentes bioquímicos sanguíneos comummente determinados no perfil metabólico representam as principais vias metabólicas do organismo, das quais a glicose, colesterol, triglicerídeos e β-hidroxibutirato representam o metabolismo energético; a ureia, hemoglobina, globulina, albumina, proteínas totais, ácido úrico representam o metabolismo proteico; o cálcio, fósforo inorgânico, magnésio, sódio e potássio representam os minerais. Adicionalmente são avaliadas a actividade das enzimas aspartato aminotransferase (AST), γ-glutamil transferase (γ-GT), alanina aminotransferase (ALT) e fosfatase alcalina (ALP), biomarcadores sanguíneos de grande valor para avaliar distúrbios metabólicos, funcionamento hepático, alterações ósseas e desequilíbrio na relação cálcio:fósforo, embora a ALT seja de pouco valor no diagnóstico em ruminantes (Mundim et al., 2007).
Vários factores podem influenciar os valores fisiológicos dos marcadores bioquímicos séricos, entre eles a raça, a idade, o sexo, a actividade física, condições nutricionais e ambientais, regime de maneio, estádio reprodutivo e da lactação, estação do ano e produção leiteira (Sakha et al., 2009).
21
Objectivos
O principal objectivo deste trabalho baseia-se na caracterização dos parâmetros hematológicos e de bioquímica sanguínea de um número pré-definido de animais (n=12) (Cabra, Capra aegagrus hircus), nomeadamente a determinação do hemograma, proteinograma, bioquímica sanguínea (tais como glicose, transaminases, fosfatase alcalina, entre outros referidos anteriormente) e ionogramas, com vista à caracterização e padronização destes parâmetros em animais saudáveis.
É ainda objectivo deste trabalho a comparação dos valores obtidos para esta raça com os valores de outras raças da mesma espécie e /ou de outras espécies próximas já caracterizados.
23
1. Modelo experimental e manutenção dos animais
Este estudo foi realizado em 12 cabras portuguesas de raça Serrana (Cabra,
Capra aegagrus hircus) saudáveis, não gestantes e não lactantes, com idades
compreendidas entre 2 e 4 anos e com peso médio de 40 kg. Os animais foram escolhidos aleatoriamente do rebanho experimental da Universidade de Trás-os-Montes e Alto Douro e criados em Vila Real (latitude 4119’N, longitude 444’W e altitude de 479 m), na Quinta do Prado, Portugal.
Todos os animais foram mantidos sob as mesmas condições ambientais. Antes de se iniciar o estudo, foi administrado uma dose de anti-helmínticos e uma injecção intramuscular de vitaminas lipossolúveis. O rebanho foi permanentemente mantido em estábulo ventilado próprio para caprinos e foi alimentado uma vez ao dia às 8:00 da manhã. A dieta foi baseada em feno suplementado com 250 g de concentrado por cada animal. A dieta foi oferecida numa estimativa de 1,20 x manutenção de energia requerida segundo as recomendações do NCR (Nutrient Requeriments of Sheep, 1985). A composição da alimentação oferecida é mostrada na Tabela 1. Os animais tinham livre acesso a água e a sal mineralizado. Semanalmente, foram retiradas amostras do feno e do concentrado oferecido aos animais que foram secadas para ser determinado a matéria seca (MS) (65ºC, 24 horas). As amostras secas de comida foram moídas, de modo a passar numa tela de 1 mm, para análises laboratoriais. Cinzas, proteínas cruas (PC) e fibras neutras de detergentes (FND) foram analisadas de acordo com AOAC (Official methods of analysis, 1990).
Tabela 1. Composição da alimentação oferecida aos animais. Matéria seca (MS) e composição química do alimento. Alimentação Feno Concentrado MS (g/kg) 88,5 90,4 Composição química* (g/kg MS) Cinzas 5,9 8,5 FND 73,3 31,6 PC 6,1 20,7
24
2. Colheita de sangue
As amostras sanguíneas foram obtidas por punção venosa na veia jugular durante a primavera no período entre as 8:00h e as 10:00h da manhã, antes da primeira alimentação do dia, e esta colheita repetida duas semanas mais tarde. Para o estudo hematológico e para obtenção do plasma, foram utilizados tubos para hematologia com EDTA (Sarstedt - Monovette – Nümbrecht, Germany) e para a determinação do tempo da trombina foram utilizados tubos com citrato de sódio (Sarstedt - Monovette - 9NC). Para o estudo de bioquímica sanguínea geral e para a determinação dos electrólitos plasmáticos, 20 mL de sangue venoso foi colhido para tubos que não continham anticoagulante (Sarstedt - Monovette - Serum Gel S). Os soros foram obtidos, recorrendo ao uso de uma centrífuga a 3000 rpm durante 10 minutos, 30 minutos depois da colheita. O hemograma e o tempo de trombina foram determinados imediatamente após a colheita da amostra sanguínea e o soro para a determinação de parâmetros bioquímicos foi pipetado para tubos cónicos de 2 mL e armazenados a -20°C para posteriormente se proceder às referidas determinações.
A determinação dos diferentes parâmetros (hemograma e parâmetros da hemostase, electroforese de proteínas e parâmetros de bioquímica sanguínea geral) foi realizada em aparelhos apropriados no Laboratório Pioledo em Vila Real.
3. Determinação dos parâmetros do hemograma
A hematologia geral engloba a determinação dos parâmetros do hemograma (eritrócitos, hemoglobina, hematócritos, volume corpuscular médio – VCM, hemoglobina corpuscular média – HCM, concentração de hemoglobina corpuscular média – CHCM, índice de distribuição eritrocitário – IDE, e leucócitos) e dos parâmetros da fórmula leucocitária (neutrófilos, eosinófilos, basófilos, linfócitos e monócitos).
25
3.1. Determinação do Hemograma
A determinação dos parâmetros do hemograma foi realizada no aparelho
Horiba ABX® (Abbott Laboratories) (Figura 12). Os resultados são obtidos por leitura automática, após calibração do aparelho.
Figura 12. Aparelho usado para a determinação do hemograma, Horiba ABX® (retirada de http://focushealth.co.in/web/index.php?option=com_content&view=article&id=15&Itemid=34).
3.2. Determinação da Fórmula Leucocitária
A determinação dos parâmetros da fórmula leucocitária foi realizada no aparelho Cell-Dyn 4000® (Abbott Laboratories) representado na Figura 13.
O Cell-Dyn 4000® é um analisador hematológico completamente automático que conta, mede e classifica as células utilizando a combinação das seguintes tecnologias: citometria de fluxo, impedância eléctrica e dispersão óptica e fluorescência. Os dados são analisados, calculados e visualizados num monitor, podendo ainda fazer-se a ligação a um sistema informático do laboratório, para gravação dos resultados.
As células são classificadas de acordo com o tamanho, estrutura interna, lobularidade nuclear e granularidade citoplasmática. Estas quatro características são fornecidas por 4 detectores de dispersão da luz. As células sanguíneas dispersam a luz
26
em várias direcções e o Cell-Dyn 4000® só vai aproveitar quatro ângulos resultantes dessa dispersão:
0 Axial Light Loss (AxLL) para definir o tamanho das células; 7 Intermediate Angle Scatter (IAS) para definir a complexidade; 90 Polarized Side Scatter (PSS) para definir a lobularidade nuclear; e 90D Depolorized Side Scatter (DSS) para definir a granularidade.
Figura 13. Aparelho utilizado para a determinação da fórmula leucocitária, Cell-Dyn 4000® (retirada http://www.pabloriviere.com/Job_History/Abbott/cd4000.htm).
A citometria de fluxo é um processo em que as células isoladas ou outras partículas biológicas, envolvidas num fluido passam alinhadas, uma a uma, em frente de um sensor, que avalia as suas características físicas e químicas. Os componentes básicos de um citómetro são: aspirador e transportador de amostra, sistema de fluxo com focagem hidrodinâmica, fonte de medida, detectores de sinal, capacidade de recolha e armazenamento de dados e capacidade de analisar e permitir a visualização de dados (Robinson, 2004; Tarrant, 2005).
27
4. Determinação dos parâmetros da hemostase
A hemostase engloba a determinação dos parâmetros do tempo de tromboplastina parcial (TTP), o tempo de protrombina (TP) e a taxa de protrombina.
Os parâmetros da hemostase foram determinados no aparelho STA Compact
CT® (AbbottLaboratories) representado na Figura 14.
Figura 14. Ilustração do aparelho usado para determinação dos parâmetros da hemóstase, STA Compact CT ® (retirado http://www.dotmed.com/listing/coagulation-analyzer/diagnostica-stago/sta-compact/775965).
4.1. Tempo de Tromboplastina Parcial (TTP)
A determinação do tempo de tromboplastina parcial é realizada de acordo com o descrito por Langdell et al. (1953) e Larrieu e Weilland (1957).
O tempo de tromboplastina parcial é um teste geral de coagulação, mais precisamente da via intrínseca da coagulação. A determinação deste parâmetro tem como principal aplicação a detecção de deficiências congénitas e adquiridas dos factores envolvidos na via intrínseca da coagulação (factores XII, XI, IX, VIII, X, V, II e I).
A determinação do TTP envolve a recalcificação do plasma na presença de uma quantidade padronizada de cefalina (um substituto plaquetário) e um factor activador XII (um composto polifenólico). O teste do TTP avalia o sistema intrínseco de
28
coagulação, isto é, os factores envolvidos e não as plaquetas. Os reagentes utilizados são a cefalina, preparada a partir de tecido cerebral de coelho de acordo com Bell e Alton (1954) e um activador polifenólico em meio tamponado. O tempo é avaliado após adição de iões cálcio para a formação do coágulo.
4.2. Tempo de Protrombina (TP)
O tempo de protrombina é um teste que tem como finalidade detectar deficiências dos factores envolvidos na via extrínseca da coagulação (factores II, V, VII e X) e do fibrinogénio (Hood e Eby, 2008).
A adição de tromboplastina com cálcio ao plasma citratado inicia a cascata de reacções, da que resulta a formação de um coágulo de fibrina. O tempo decorrido até ao início da coagulação é comparado com o tempo determinado utilizando o padrão normal. As actividades dos seguintes factores de coagulação são medidas: factor II (protrombina), factor V (proacelerina), factor VII (proconvertina) e factor X (factor Stuart-Prower). O tempo decorrido entre a pipetagem do reagente e a formação do coágulo é medido. Os resultados podem ser expressos como uma percentagem da actividade normal com a ajuda de curvas de calibração. Os reagentes utilizados neste teste são um reagente de tromboplastina (cérebro de coelho) liofilizado e solução de cloreto de cálcio (aproximadamente 1,47 g/L).
5. Determinação da electroforese de proteínas
A técnica de electroforese é actualmente um método padrão para o fraccionamento das proteínas séricas em bioquímica clínica. Existem vários métodos para a electroforese de proteínas séricas que diferem apenas no tipo de suporte utilizado (Berg, 2002).
O princípio da separação electroforética das proteínas séricas é baseado na migração das partículas proteicas carregadas num campo eléctrico. A direcção e a taxa
29
de migração das partículas são baseadas no tipo de carga da proteína (+ ou -), na dimensão da proteína, na intensidade do campo eléctrico e no suporte através do qual as partículas proteicas são induzidas a migrar. O suporte, o pH, o tampão usado e a intensidade de corrente eléctrica aplicada devem ser plenamente descritos de modo a poderem comparar os resultados da electroforese das proteínas séricas (Berg, 2002).
Historicamente, o suporte de acetato de celulose era o mais comummente utilizado, mas actualmente o gel de agarose é amplamente usado em bioquímica clínica. Além dos suportes mencionados, o gel de agar, o gel de amido e o gel de poliacrilamina são também utilizados, contudo com menor frequência (Berg, 2002).
Neste estudo, a determinação da electroforese de proteínas foi realizada no aparelho CapillarysTM2 (AbbottLaboratories) tal como representado na Figura 15, por electroforese capilar. Os resultados obtidos foram guardados em ficheiros para posterior análise.
Figura 15. Aparelho utilizado para a determinação da electroforese de proteínas, CapillarysTM2 (retirado www.sebia-usa.com/products/capillarys.html).
6. Determinação dos parâmetros de bioquímica sanguínea
Os parâmetros bioquímicos foram determinados com kits disponíveis comercialmente, nomeadamente para a avaliação da glicose, triglicerídeos, colesterol, aspartato aminotransferase, alanina aminotransferase, γ-glutamil transferase, fosfatase alcalina, ureia, creatinina, cálcio, fósforo, magnésio, sódio, potássio e cloreto.
30
A determinação de cada um destes parâmetros foi realizada por espectrofotometria de absorção molecular num analisador bioquímico automático, o Architect Ci8200® (AbbottLaboratories) (Figura 16).
Figura 16. Aparelho usado para a determinação dos parâmetros de bioquímica sanguínea, Architect Ci8200 (retirada http://dila.com.ua/about/tools/).
6.1. Glicose
A determinação da glicose no soro baseia-se na metodologia da enzima hexocinase/glicose-6-fosfato desidrogenase (G-6-PDH), a qual é realizada por um kit comercial (referência 3L82, Abbott®) incorporado no aparelho contendo os seguintes reagentes NAD+ (5,0 mg/mL), G-6-PDH (3000 U/L), hexocinase (15000 U/L) e ATP∙2Na (9,0 mg/mL).
A glicose é fosforilada pela enzima hexocinase na presença da adenosina trifosfato (ATP) e iões magnésio produzindo glicose-6-fosfato (G-6-P) e adenosina difosfato (ADP). A glicose-6-fosfato desidrogenase (6-PDH) oxida especificamente G-6-P a 6-fosfogluconato com a concomitante redução da nicotinamida adenina dinucleotído (NAD+) a nicotinamida adenina dinucleotído (NADH). Uma micromole de NADH é produzida for cada micromole de glicose consumida. A NADH produzida
31
absorve luz a 340 nm e pode ser determinada espectrofotometricamente com o aumento da absorvância (Glucose, 3L82-21).
6.2. Triglicerídeos
A determinação dos triglicerídeos baseia-se na metodologia da enzima glicerol fosfato oxidase, utilizando um kit comercial (referência 7D74, Abbott®) que contém os seguintes reagentes ATP (2,5 mmol/L), Mg2+ (2,5 mmol/L), 4-amino-antipirina (0,4 mmol/L), 4-clorofenol (2 mmol/L), peroxidase (> 2000 U/L), glicerol cinase (> 600 U/L), glicerol fosfato oxidase (> 6000 U/L) e lipoproteína lípase (> 3000 U/L).
Os triglicerídeos são hidrolisados enzimaticamente pela lipase a ácidos gordos livres e glicerol. O glicerol é fosforilado pelo ATP e glicerol cinase produzindo 3-fosfato e ADP. O 3-3-fosfato é oxidado a di-hidroxiacetona 3-fosfato pela glicerol-3-fosfato oxidase, produzindo peróxido de hidrogénio (H2O2).
Numa reacção de cor catalisada pela peroxidase, o H2O2 reage com
4-amino-antipirina e 4-clorofenol, produzindo um corante vermelho. A absorvância desse corante é proporcional à concentração de triglicerídeos presente na amostra.
Esta metodologia analítica é baseada na sequência de reacções descritas por Fossati et al. (1982) e McGowan et al. (1983).
6.3. Colesterol
A determinação do colesterol baseia-se na metodologia enzimática, utilizando um kit comercial (referência 7D62, Abbott®) que contém os seguintes reagentes: colesterol oxidase (> 200 U/L), colesterol esterase (> 500 U/L), peroxidase (> 300 U/L), 4-amino-antipirina (< 0,5 mmol/L) e ácido hidroxibenzóico (10 mmol/L).
Os ésteres de colesterol são hidrolisados enzimaticamente pela enzima colesterol esterase produzindo colesterol e ácidos gordos livres. O colesterol livre,
32
incluindo o originalmente presente, é então oxidado pela enzima colesterol oxidase produzindo colest-4-ene-3-one e H2O2. O H2O2 combina-se com o ácido
hidroxibenzóico e 4-amino-antipirina formando um cromóforo (corante quinoneimina) que é quantificada a 500 nm (Allain et al., 1974; Roeschlau et al., 1974).
6.4. Aspartato aminotransferase (AST)
Na determinação da aspartato aminotransferase o aparelho utiliza um kit comercial (referência 7D81, Abbott®) contendo os reagentes R1 e R2. O R1 contém β-NADH (0,16 mg/mL), malato desidrogenase (0,64 U/mL), lactato desidrogenase (0,64 U/mL) e aspartato (232 mmol/L). O R2 contém α-cetoglutarato (51,3 mmol/L) e L-aspartato (100 mmol/L). A metodologia utilizada baseia-se na absorvância do NADH a 340 nm.
A aspartato aminotranferase presente na amostra catalisa a transferência do grupo amino do aspartato para o α-cetoglutarato, formando oxaloacetato e L-glutamato. O oxaloacetato na presença de NADH e enzima malato desidrogenase é reduzido a L-malato. Nesta reacção, o NADH é oxidado a NAD+. A reacção é monitorizada pela medição da taxa de decréscimo da absorvância a 340 nm devido à oxidação do NADH a NAD+ (Friedman e Young, 1989; Burtis et al., 1994).
6.5. Alanina aminotransferase (ALT)
A determinação da alanina aminotranferase é realizada através de um kit comercial (referência 7D56, Abbott®) que utiliza a metodologia da absorvância do NADH a 340 nm. Esse kit contém os regentes R1 e R2. No R1 estão presentes o β-NADH (0,16 mg/mL), o lactato desidrogenase (2,57 U/mL) e a L-alanina (392 mmol/L). No R2 estão presentes o ácido α-cetoglutárico (77 mmol/L) e L-alanina (1000 mmol/L).
A alanina aminotransferase presente na amostra catalisa a transferência do grupo amino da L-alanina para o α-cetoglutarato, formando piruvato e L-glutamato. O
33
piruvato na presença de NADH e lactato desidrogenase é reduzido a L-lactato. Nesta reacção o NADH é oxidado a NAD+. A reacção é monitorizada pela medição da taxa de decréscimo da absorvância a 340 nm devido à oxidação do NADH a NAD+ (Alanine Aminotransferase, 7D56-21).
6.6. γ-glutamil transferase (γ-GT)
A determinação da γ-glutamil transferase é realizada através de um kit comercial (referência 7D66, Abbott®) contendo os reagentes R1 e R2, utilizado a metodologia baseada no substrato L-γ-glutamil-3-carboxi-4-nitroanilida. O R1 contém glicilglicina (191 mmol/L), enquanto o R2 contém L-γ-glutamil-3-carboxi-4-nitroanilida (30,6 mmol/L).
A γ-glutamil transferase catalisa a transferência do grupo γ-glutamil do substrato dador (L-γ-glutamil-3-carboxi-4-nitroanilida) para o aceitador glicilglicina para produzir 3-carboxi-4-nitroanilina. A taxa do aumento de absorvância a 412 nm é directamente proporcional à glutamil transferase na amostra. O procedimento da γ-GT é uma modificação do método descrito por Theodorsen et al. (1976).
6.7. Fosfatase alcalina (ALP)
A determinação da fosfatase alcalina é realizada por um kit comercial (referência 7D55, Abbott®) contendo os reagentes R1 e R2 na sua composição. No R1 estão presentes o 2-amino-2-metilpropanol (> 1,2 mol/L), o magnésio (> 7,2 mmol/L), o sulfato de zinco (> 3,6 mmol/L) e HEDTA (> 7,2 mmol/L). O R2 contém a 4-nitrofenil fosfato (> 171,6 mmol/L).
Vários substratos têm sido utilizados para a determinação da actividade da fosfatase alcalina, tais como glicerolfosfato (King e Armstrong, 1934), fenil fosfato (King e Armstrong, 1934) e ρ-nitrofenil fosfato (Bessey et al., 1946). Bowers e McComb (1966) melhoraram o método de Bessey et al. (1946) para abranger a determinação
34
cinética. Tietz et al. (1983) optimizaram este método de forma a incluir um tampão quelatado de iões metal de zinco, magnésio e HEDTA. Esta medição da fosfatase alcalina é uma modificação deste método.
A fosfatase alcalina na amostra catalisa a hidrólise de ρ-nitrofenil fosfato (incolor), originando nitrofenol e fosfato inorgânico. No pH do ensaio (alcalino), o ρ-nitrofenol encontra-se na forma fenóxida amarela. A taxa do aumento de absorvância a 404 nm é directamente proporcional à actividade da fosfatase alcalina na amostra. As concentrações optimizadas dos iões zinco e magnésio estão presentes para a actividade da fosfatase alcalina na amostra.
6.8. Ureia
A determinação da ureia realiza-se através de um kit comercial (referência 7D75, Abbott®), que contém os reagentes R1 e R2. O R1 contém NADH (2,95 mmol/L) e azida de sódio (< 0,1 %) e o R2 contém ácido α-cetoglutárico (99,8 mmol/L), urease (feijão) (23,5 KU/L), glutamato desidrogenase (fígado de bovino) (63,5 KU/L), adenosina difosfato (ADP) (7,6 mmol/L) e azida de sódio (< 0,2 %).
A análise da ureia é uma modificação de um procedimento totalmente enzimático inicialmente descrito por Talke e Schubert (1965). O teste é realizado como uma análise cinética em que a taxa inicial da reacção é limitada durante um período de tempo. A ureia na amostra é hidrolisada pela enzima urease em amónia e dióxido de carbono. A segunda reacção, catalisada pela enzima glutamato desidrogenase, converte amónia e α-cetoglutarato em glutamato e água com a concomitante oxidação da NADH reduzida em NAD+. Assim, duas moles de NADH são oxidadas por cada mole de ureia presente. A taxa inicial da diminuição na absorvância a 340 nm é proporcional à concentração de ureia na amostra.