Universidade de Trás-os-Montes e Alto Douro
INDUÇÃO DO PARTO EM COELHAS COM PROSTAGLANDINAS OU
OXITOCINA. EFEITOS SOBRE AS PERFORMANCES PRODUTIVAS
DAS REPRODUTORAS E DAS NINHADAS.
Dissertação para a obtenção do grau de
Mestre em Engenharia Zootécnica
Gustavo Daniel Martins Nunes
Orientador
Prof
.
Doutor Victor
Manuel de Carvalho
Pinheiro
CECAV - UTAD
Coorientador
Prof. Doutor José Carlos Marques de Almeida
CECAV - UTAD
I
Universidade de Trás-os-Montes e Alto Douro
INDUÇÃO DO PARTO EM COELHAS COM PROSTAGLANDINAS OU
OXITOCINA. EFEITOS SOBRE AS PERFORMANCES PRODUTIVAS
DAS REPRODUTORAS E DAS NINHADAS.
Dissertação para a obtenção do grau de
Mestre em Engenharia Zootécnica
Gustavo Daniel Martins Nunes
Orientador
Prof
.
Doutor Victor
Manuel de Carvalho
Pinheiro
Coorientador
Prof. Doutor José Carlos Marques de Almeida
Composição do Júri:
Doutor José Júlio Gonçalves Barros Martins
Doutor José Luís Teixeira de Abreu Medeiros Mourão Doutor José Carlos Marques de Almeida
Doutor Victor Manuel de Carvalho Pinheiro
III “As doutrinas apresentadas neste trabalho são da exclusiva responsabilidade do autor”
V
Agradecimentos
Ao Professor Doutor Victor Pinheiro por ter aceite a orientação desta tese, por ter acreditado no meu trabalho e por toda a confiança que depositou em mim.
Ao Professor Doutor José Carlos Almeida, coorientador deste trabalho, pelo seu incentivo, pela ajuda no delineamento da experiência e interpretação dos resultados e pela revisão do trabalho.
Aos restantes Professores do Departamento de Zootecnia da UTAD e também aos Professores do Departamento de Zootecnia da U.Évora, pelos ensinamentos e pela total disponibilidade para me ajudar sempre que necessitei, mas acima de tudo por me terem transmitido a paixão que tenho pela Zootecnia.
Ao Engenheiro Paulo Fontes e ao Engenheiro António Silva pela sua disponibilidade e pelo apoio prestado na realização do trabalho prático.
Aos meus colegas de casa durante a minha passagem por Vila Real, António, Ana, David e Nuno, agradeço pela amizade, pela paciência e pelo companheirismo.
A todos os meus grandes amigos, que me dão força e preenchem o meu coração e especialmente à “Dream Team”, que tão importante foi para mim nesta vivência académica.
À Daniela, pelo apoio incondicional, pela compreensão, por aceitar a distância e mesmo assim estar sempre comigo e me fazer sorrir de uma forma que só ela consegue.
À minha grande família, Tias, Tita, Primas, Primos, Avó Inácia, Avó Adelaide e Avôs Antónios. Sem vocês não seria aquilo que sou.
Por último, mas sem dúvida o mais importante, queria mostrar a minha profunda gratidão aos meus pais, Libânia e José Eduardo, e ao meu irmão, Cristóvão, por todo o esforço que fizeram para me dar uma educação superior, por incentivarem sempre as minhas escolhas, por me apoiarem em todos os momentos e por me mimarem sempre que podem.
VII
Resumo
Título: Indução do parto em coelhas com prostaglandinas ou oxitocina. Efeitos sobre as
performances produtivas das reprodutoras e das ninhadas.
Foi avaliada a produtividade de coelhas sujeitas a induções de parto sucessivas com cloprostenol (análogo sintético da PGF2α) ou oxitocina. O ensaio teve início com 75 coelhas
nulíparas, inseminadas às 18 semanas de idade, que foram acompanhadas ao longo de 4 ciclos reprodutivos (desde a IA até ao desmame dos láparos). As coelhas foram alojadas em jaulas individuais e divididas em 4 tratamentos: grupo controlo (C; n=18), no qual o parto não foi induzido; grupo PG28, em que foi administrado cloprostenol (Estrumate®), ao dia 28 de gestação (n=19); grupo PG29, em que foi administrada a mesma substância mas ao dia 29 de gestação (n=19); e grupo OX30, em que foi administrado oxitocina (Placentol®) ao dia 30 de gestação (n=19). Foram administrados 0,2ml/coelha de Estrumate® e 0,15ml/kgPV de Placentol®, por via intramuscular. Ao longo do ensaio foi controlada a taxa de fertilidade e a prolificidade das coelhas, a mortalidade das reprodutoras e dos láparos e a performance de crescimento dos láparos através de pesagens ao nascimento, aos 18 dias de idade e ao desmame. Os dados referentes ao 1º ciclo reprodutivo não foram contabilizados. Apesar de não se terem verificado diferenças significativas para a taxa de fertilidade entre os tratamentos, as diferenças observadas foram elevadas, sendo os grupos PG28 e PG29 os que apresentaram maior valor em comparação ao grupo C (82,2% e 87,8% vs. 73,6%, respetivamente). A prolificidade foi significativamente afetada pelos tratamentos, com o grupo PG28 a apresentar um valor superior em relação ao grupo C (11,05 vs. 9,25). O peso da ninhada ao nascimento não foi afetado pelo tratamento, mas o peso individual dos láparos ao nascimento foi menor no grupo PG28 (51,6 g) em relação a C e OX30 (57,3 g e 59,8 g, respetivamente). O grupo OX30 apresentou elevada incidência de distócia (33,3%). A taxa de mortalidade das reprodutoras e dos láparos não foi afetada pelos tratamentos, assim como os ganhos médios diários dos láparos até ao desmame. Não se verificaram diferenças significativas entre os tratamentos para a produtividade das coelhas (Kg de láparo desmamado por coelha inseminada), embora os grupos PG28 e PG29 apresentem valores mais elevados em comparação ao grupo C (4,93 Kg e 4,93 Kg vs. 4,45 Kg, respetivamente). Embora o estudo não seja conclusivo em relação à produtividade, verificou-se que a indução do parto com o análogo sintético da PGF2α pode melhorar a
performance reprodutiva das coelhas. Uma maior dimensão da amostra poderia ser mais expressiva em termos de resultados.
IX
Abstract
Title: Induction of parturition in the rabbit does using prostaglandins or oxytocin. Effects on
productivity of does and litters.
Was evaluated the rabbit’s productivity when subjected to successive induced parturitions with cloprostenol (a synthetic PGF2α analogue) or oxytocin. The trial began with
75 nuliparous does, which were inseminated at 18 weeks old. The does were followed during the 4 reproductive cycles (from artificial insemination to weaning). The does were housed in individual cages and divided in 4 treatments: control group (C; n=18), in which the parturition was not induced; PG28 group, in which was administrated cloprostenol (Estrumate®) on 28th day of pregnancy (n=19); PG29 group, in which was administrated the same substance but on 29th day of pregnancy (n=19); and OX30 group, which was administrated oxytocin (Placentol®) on 30th day of gestation (n=19). Were injected 0,15ml/kgBW of Placentol® and 0,2ml/doe of Estrumate®, intramuscularly. During the trial was controlled the fertility rate and the prolificacy of the does, the mortality of does and kits and also the growth performance of kits, throw the weighing at birth, at 18th day old and on weaning. The data about the 1st reproductive cycle were not accounted. Although there have been no significant differences about the fertility rate between treatments, the differences observed were high. The PG28 and PG29 groups showed a higher value comparative to C (82,2% and 87,8% vs. 73,6%, respectively). The prolificacy was significantly affected by the treatments. The PG28 group presented a higher value than C (11,05 vs. 9,25). The litter birth weight was not affected by treatment, but the individual birth weight of kits was lower on PG28 (51,6 g) comparative to C or OX30 (57,3g and 59,8g, respectively). OX30 presented a high incidence of dystocia (33,3%). The does mortality rate and the kits mortality rate were not affected by treatments and also the daily average growth of kits until weaning. There were no significant differences between treatments for the productivity (Kg of kits weaned per inseminated doe), although the groups PG28 and PG29 presented a higher value than C (4,93 Kg e 4,93 Kg vs. 4,45 Kg, respectively). Despite the study was not conclusive about the productivity, was showed that the induced parturition with the synthetic PGF2α analogue can improve the does reproductive
performance. A larger sample could be more expressive about the results.
XI
Índice
1 Introdução... 1
2 Revisão Bibliográfica ... 3
2.1 Ciclo reprodutivo e produtividade da coelha ... 3
2.2 Comportamento maternal da coelha ... 6
2.3 Mecanismos que desencadeiam o parto ... 7
2.4 Utilização de prostaglandinas e oxitocina em produção animal ...10
2.5 Indução do parto em cunicultura (estado da arte) ...11
3 Trabalho experimental ...15 3.1 Objetivos ...15 3.2 Material e Métodos ...16 3.2.1 Animais testados ...16 3.2.2 Profilaxia ...16 3.2.3 Período de ensaio ...17 3.2.4 Alojamento ...17 3.2.5 Alimentação ...18
3.2.6 Tratamentos e delineamento experimental ...19
3.2.7 Determinações ...21 3.2.8 Análise de dados ...23 3.3 Resultados e Discussão ...24 3.3.1 Performance reprodutiva ...25 3.3.1.1 Fertilidade ...25 3.3.1.2 Duração da gestação ...27 3.3.1.3 Parto ...31
XIII
3.3.1.4 Características das ninhadas à nascença ...37
3.3.1.5 Comportamento maternal ...41
3.3.2 Performance na lactação e evolução das ninhadas até ao desmame ...45
3.3.2.1 Número de láparos e mortalidade das ninhadas na lactação ...45
3.3.2.2 Produção leiteira e crescimento das ninhadas ...47
3.3.3 Ingestão de Alimento e variação do Peso Vivo das reprodutoras ...49
3.3.4 Mortalidade das reprodutoras ...51
3.3.5 Produtividade global ...52
4 Considerações finais ...54
5 Referências Bibliográficas ...57
XV
Índice de figuras
Figura 1 – Alterações nas concentrações plasmáticas de progesterona, estradiol,
testosterona e prolactina durante o final da gestação, parto e lactação da
coelha.. ... 9
Figura 2 – Pavilhão onde decorreu o ensaio.. ...17 Figura 3 – Esquema da distribuição dos grupos pelas jaulas de ensaio. ...20 Figura 4 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina na taxa de fertilidade de coelhas ...25
Figura 5 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina na duração da gestação. ...27
Figura 6 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina na distribuição dos partos no tempo. ...28
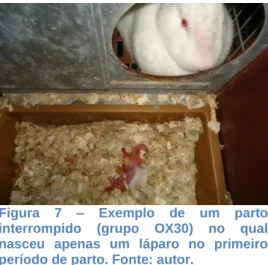
Figura 7 – Exemplo de um parto interrompido (grupo OX30) no qual nasceu apenas um
láparo no primeiro período de parto. ...32
Figura 8 – Administração do tratamento hormonal via intramuscular a uma das
reprodutoras. ...35
Figura 9 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina na dimensão da ninhada ...38
Figura 10 – Recolha dos láparos após o parto para pesagem das ninhadas. ...39 Figura 11 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina no local de ocorrência do parto...41
Figura 12 – Exemplo de ninhada parida na jaula com sinais explícitos de canibalismo e
grande parte dos láparos mortos por arrefecimento. ...42
Figura 13 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
XVII
Índice de tabelas
Tabela 1 – Composição do alimento fornecido durante o ensaio. ...18 Tabela 2 – Caracterização dos grupos de ensaio ...19 Tabela 3 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina: no tempo de intervalo entre a administração e o início do parto; no tempo de duração do parto; e na incidência de partos
interrompidos. ...31
Tabela 4 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina no nº de láparos nascidos e no seu peso ao nascimento. ...37
Tabela 5 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina nos comportamentos maternais das
reprodutoras. ...43
Tabela 6 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina no número de láparos e na sua mortalidade até ao desmame. ...46
Tabela 7 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina no peso e no GMD dos láparos amamentados pelas coelhas tratadas. ...47
Tabela 8 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina na Ingestão Média Diária e no Peso Vivo de coelhas lactantes. ...49
Tabela 9 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina nas causas de mortalidade das reprodutoras entre o 2º e o 4º ciclo reprodutivo. ...51
Tabela 10 – Influência da indução de parto com cloprostenol (administração ao dia 28 ou ao
dia 29 de gestação) ou oxitocina nos indicadores de produtividade das
XIX
Lista de abreviaturas
µg – micrograma
ACTH - Hormona adrenocorticotrófica (Adrenocorticotropic hormone) C – Controlo/parto não induzido (grupo de ensaio)
CL – Corpo lúteo cm – centímetro d – dia
eCG – Gonadotrofina coriónica equina (Equine chorionic gonadotropin) EPM – Erro padrão da média
FSH – Hormona folículo-estimulante (Follicle-stimulating hormone) g – grama
GMD – Ganho médio diário
GnRH – Hormona libertadora das gonadotrofinas (Gonadotropin-releasing hormone) h – hora
IA – Inseminação artificial IGM – Ingestão média diária Kg – Quilograma
LH – Hormona luteínizante (Luteinizing hormone) mg – miligrama
min – minuto
n.s. – não significativo
OX30 – Oxitocina administrada ao dia 30 de gestação (grupo de ensaio) PG28 – Cloprostenol administrado ao dia 28 de gestação (grupo de ensaio) PG29 – Cloprostenol administrado ao dia 29 de gestação (grupo de ensaio) PGE2 – Prostaglandina E2 (Prostaglandin E2)
PGF2α – Prostaglandina F2α (Prostaglandin F2α)
1
1 Introdução
Um controlo preciso do tempo em que vai ocorrer o parto é uma ferramenta preciosa e cada vez mais importante na gestão de grandes efetivos pecuários. Desde há alguns anos atrás, a utilização de tratamentos hormonais à base de prostaglandinas, corticosteroides, ou oxitocina tornaram-se comuns nos sistemas intensivos de produção animal como auxiliares do controlo reprodutivo, quer em procedimentos para indução de cios, quer devido ao seu efeito indutor do parto.
Principalmente devido ao desenvolvimento de protocolos de indução do parto mais baratos e seguros, esta técnica tem sido, cada vez mais, posta em prática em contexto produtivo e alvo de investigação um pouco por todo o mundo. A utilização de análogos sintéticos da PGF2α, como o Cloprostenol ou Fenprostalene, e a utilização de vias de
administração mais adequadas, como a via intramuscular ou a via submucosa vulvar, permitem a redução das doses utilizadas e um aumento da eficiência dos tratamentos.
A indução do parto é utilizada principalmente em produção intensiva de bovinos, ovinos, caprinos, suínos e coelhos, com o intuito de aumentar a eficiência de produção, agrupando os partos em alturas mais propícias, reduzindo o intervalo entre partos e diminuindo a necessidade de supervisão das reprodutoras durante a época de partos. A técnica ganha especial importância quando se fala de zootecnia de precisão e melhoramento animal, em que se investe bastante dinheiro na utilização de reprodutores de elevado potencial e em técnicas como a superovulação, fecundação in-vitro ou transferência de embriões, havendo nestes casos uma necessidade acrescida de minimizar as perdas na fase final do processo reprodutivo, ou seja, o parto. Ao garantir que as crias vão nascer num curto intervalo de tempo pré-determinado, consegue-se garantir a presença de assistência capacitada para auxiliar no parto, caso esta seja necessária, e assim reduzir a mortalidade das crias.
Apesar das vantagens, a indução de parto continua a ter alguns efeitos colaterais nas diferentes espécies, tais como: o aumento de partos distócicos, o atraso no desenvolvimento das crias, a diminuição temporária da produção de leite, a maior incidência de retenções placentárias e de patologias daí recorrentes e ainda o possível comprometimento da performance reprodutiva futura das progenitoras.
É um facto que a indução/sincronização de partos nas diferentes espécies pecuárias é considerada uma boa ferramenta para o controlo de algumas doenças, para fins didáticos e de investigação. No entanto, a sua utilização de forma corrente, voltada exclusivamente
2
para o incremento dos parâmetros produtivos de sistemas de produção pecuária, continua a suscitar dúvidas e a levantar questões relativas ao bem-estar animal, devendo considerar-se não só a relação custo/benefício, mas também as consequências advindas da utilização exagerada desta prática, decorrendo daí o estabelecimento e/ou agravamento de distúrbios até então inexistentes ou de baixa incidência.
Especificamente em cunicultura, a utilização desta técnica não é muito expressiva e a sua eficácia no incremento de produtividade é ainda algo controverso. Porém, é um tema bastante estudado, principalmente a indução do parto com análogos da prostaglandina e por parte de investigadores Europeus (Espanha, Itália, Bélgica, entre outros) e Norte-Americanos. Já a indução do parto com oxitocina está mais estudada em condições laboratoriais, principalmente por investigadores da área da medicina humana. A informação sobre a sua utilização num contexto de produção é escassa. Rebollar, Ubilla, McNitt, Partridge, Negatu e Fuchs são apenas alguns exemplos de investigadores que se dedicaram a estudar estes assuntos. Em relação a Portugal a informação publicada sobre o tema é escassa e supõe-se que esta técnica seja muito pouco utilizada por parte dos cunicultores portugueses.
A decisão estratégica do uso deste tipo de técnicas de maneio em cunicultura deve ter em conta o seu impacto sobre a produção, tanto do ponto de vista biológico como financeiro, sendo necessário investigação que analise a utilização destas técnicas com uma abordagem global, interligando a fisiologia com o maneio.
Este estudo pretende testar vários procedimentos utilizados para a indução do parto, nomeadamente a utilização de cloprostenol (análogo sintético da PGF2α) aplicado a
diferentes datas (28º e 29º dias de gestação) e a utilização de oxitocina. O principal objetivo será verificar a eficiência deste tipo de tratamentos de forma abrangente e perceber os reflexos a nível produtivo da sua utilização ao longo de vários ciclos reprodutivos nos mesmos animais, abordando também as implicações em termos de maneio que a sua utilização acarreta numa cunicultura comercial.
3
2 Revisão Bibliográfica
2.1 Ciclo reprodutivo e produtividade da coelha
A atividade reprodutiva dos coelhos é controlada pelo eixo neuro-endócrino. Porém, fatores como a genética, alimentação e o maneio têm uma forte influência na regulação hormonal (Castellini et al., 2010). Este animal tem algumas particularidades reprodutivas que não são comuns à maioria dos mamiferos.
Nas fêmeas dos bovinos, ovinos, suínos e outros mamíferos a ovulação ocorre espontâneamente com intervalos regulares durante o período de cío ou estro. Diferindo dessas espécies, a coelha não tem ciclo éstrico tão definido, com aparecimento regular de cio quando se dá a ovulação. Nesta fêmea a ovulação e consequente formação do CL só se dão após o estímulo da cópula, sendo por isso classificada como espécie de ovulação induzida. Na ausência do estímulo necessário, a fêmea não ovula e pode manter-se em estro durante períodos mais ou menos longos (Mourão, 2003).
Quando se utiliza a inseminação artificial (IA) em coelhas, o estímulo associado não é suficiente para desencadear a ovulação, sendo obrigatório a aplicação de um maneio adequado para a induzir. Geralmente administra-se GnRH exógena (ou produtos análogos) no momento da IA para induzir a ovulação (Alvariño & Rebollar, 1995; Castellini et al., 2010). Estes produtos são eficazes, económicos e podem ser utilizados sem aparecerem reações imunulógicas (Mourão & Pinheiro, 2004).
Apesar da ovulação não ser espontânea nas coelhas, numerosas observações mostram a existência de alternância entre períodos de estro, durante os quais aceita a cobrição, e de períodos de recusa. A duração destes períodos é variável entre coelhas, podendo haver fêmeas com um período recetivo de 28 dias seguido de um ou dois dias de recusa, enquanto que outras têm 2 dias recetivos seguidos de 4 semanas de recusa. Esta recetividade está relacionada com o ciclo de maturação folicular e com o número de folículos pré-ovulatórios existentes nos ovários da coelha (Mourão, 2003).
O comportamento sexual da coelha, particularmente a recetividade, é dependente dos esteróides ováricos. Os estrogénios e os androgénios produzidos pelo ovário favorecem a aceitação do macho, enquanto que a progesterona tem um efeito depressivo (Mourão & Pinheiro, 2004). O facto de algumas coelhas terem folículos no estado pré-ovulatório quase
4
continuamente, enquanto que outras só têm alguns dias do mês, é um obstáculo à sincronização da reprodução nas explorações (Mourão, 2003). Porém as coelhas podem ser inseminadas independentemente da sua recetividade sexual (Castellini et al., 2010).
Os produtores comerciais de coelhos procuram produtividades elevadas intensificando o ritmo reprodutivo, por isso as coelhas são geralmente inseminadas durante a lactação, período na qual a recetividade é mais variável, em função da fase da lactação. As coelhas conseguem manter a lactação e a gestação simultaneamente, porém esta sobreposição deprime vários aspetos da atividade reprodutiva (recetividade, fertilidade, prolificidade) devido ao antagonismo hormonal entre a prolactina e as gonadotropinas e ao défice energético que se observa nessa altura (Mourão & Pinheiro, 2004; Castellini et al., 2010).
Inseminar as coelhas durante a lactação afeta negativamente a performance reprodutiva, pois as coelhas não recetivas têm uma taxa de fecundidade e fertilidade mais baixas. Como tal, é importante que a coelha esteja recetiva no momento da IA, sendo necessário recorrer a técnicas de sincronização do ciclo reprodutivo nas coelhas de forma a obter taxas de fertilidade elvadas (Castellini et al., 2010).
O ritmo reprodutivo semi-intensivo (IA ao 11º dia pós-parto) é largamente utilizado em cunicultura, mas não tem em consideração a fisiologia das coelhas lactantes, sendo necessário recorrer-se a tratamentos para indução e sincronização da recetividade das coelhas (Castellini et al., 2010). A administração de hormonas estimulantes do desenvolvimento folicular, como a eCG, provoca a maturação de uma nova vaga de folículos num prazo de 48 a 72 h, acompanhada do aumeto dos níveis de estrogénios e aparecimento consecutivo do comportamento de estro (Alvariño & Rebollar, 1995; Mourão, 2003). A administração de eCG 48 h antes da IA é a técnica mais utilizada em cunicultura para indução da recetividade, obtendo-se resultados positivos regulares com a sua aplicação, mesmo em coelhas lactantes. A utilização deste tratamento também aumenta o tamanho da ninhada. Porém a sua utilização continuada pode perder eficácia, pois pode surgir uma resposta imunitária das coelhas contra esta hormona de origem exógena (Mourão & Pinheiro, 2004; Castellini et al., 2010) e neste sentido têm sido estudados e aplicados metodos alternativos ao uso hormonal, designados por métodos de bioestimulação.
Algumas das técnicas de bioestimulação a aplicar antes da IA que mostram resultados positivos no aumento da recetividade e da fertilidade são: flushing energético; aumento do fotoperíodo; a manipulação da fêmea/troca de jaula; e a separação temporária entre fêmea
5 e ninhada. Entre estes métodos o melhor e mais utilizado, tanto pela viabilidade prática em larga escala, como pelos resultados obtidos até ao momento, é a separação temporária entre mãe e ninhada, durante 24 a 48 h antes da IA (Theau-Clément & Boiti, 1998).
A gestação na coelha dura em média 30 a 32 dias, sendo o valor máximo de 33 dias e o mínimo de 28 dias. Com duração inferior nascem nados mortos. Em geral só 70 a 80% dos embriões dão origem a láparos vivos (Mourão, 2003). O número de láparos por parto pode variar de 1 a 20. No entanto, a prolificidade das estirpes de coelhos utilizados na produção intensiva, situa-se entre 8 e 11 láparos nascidos vivos por parto (Simões, 2008). Todavia, a prolificidade é uma característica dependente da taxa de ovulação, da viabilidade dos fetos e de diversos fatores de maneio e ambientais (Mourão & Pinheiro, 2004).
No período antecedente ao parto, e durante este, desencadeia-se um complexo processo hormonal que altera os níveis de progesterona, estrogénios e de outras hormonas da coelha, determinando o comportamento de construção do ninho, o final da gestação, a dilatação da cérvix, o início das contrações uterinas, o comportamento maternal e o inicio da lactação (González-Mariscal, 2001).
Após o parto a coelha entra em lactação, amamentando os láparos durante um período de aproximadamente 4 a 5 semanas (quando se utiliza o ritmo reprodutivo semi-intensivo). Todavia, em condições naturais o período de amamentação pode prolongar-se até aos 45 dias. O efeito negativo que a lactação tem na recetividade e na fertilidade acentua-se quando o número de láparos aleitantes aumenta. O momento (dia) da lactação influencia igualmente a taxa de ovulação e o número de óvulos libertados. No 3º e 4º dias de lactação há um efeito particularmente depressivo sobre estes parâmetros (Mourão & Pinheiro, 2004).
A regressão do útero não é obstáculo absoluto ao arranque de uma nova gestação, devendo ter-se em conta que uma regressão uterina inacabada poderá aumentar a mortalidade embrionária ou diminuir o número de embriões que se implantam (Mourão & Pinheiro, 2004). Porém, o útero das coelhas regride rápidamente depois do parto, perdendo metade do tamanho em 2 dias (Mourão, 2003).
A produtividade numérica das coelhas (nº láparos desmamados/nº coelhas à reprodução) depende da recetividade, fertilidade e prolificidade. Em condições ideais, com um ritmo de reprodução semi-intensivo, cada fêmea pode produzir 64 a 82 láparos
6
nascidos/ano, e produtividades anuais na ordem dos 50 a 70 láparos desmamados/coelha (Simões, 2008).
A utilização deste ritmo de reprodução obriga à realização do desmame precoce. É comum realizá-lo por volta dos 31 a 35 dias após o nascimento dos láparos. Desmamar os láparos nesta altura permite que a coelha não esteja em lactação na fase final da gestação, quando as necessidades para o desenvolvimento fetal são maiores (Mourão & Pinheiro, 2004).
2.2 Comportamento maternal da coelha
O comportamento maternal da coelha consiste na construção de um ninho com palha (ou outro material disponível) e pelos que arranca do próprio corpo durante a fase final da gestação e, posteriormente, consiste nos cuidados que presta à ninhada até à fase do desmame (Caba et al., 1996; González-Mariscal, 2001).
No comportamento de construção do ninho, a coelha transporta para o ninho materiais que tenha à sua disposição (p.ex. feno, palha ou aparas de papel), e que mistura com o seu pelo. Normalmente, retira pelagem da zona abdominal, o que facilita o acesso aos tetos por parte dos láparos, mas muitas são as coelhas que retiram pelo de praticamente todo o corpo (Xu, 1996). O parto acontece geralmente durante as primeiras horas de luz do dia (Boiti, 1999; Ninomiya-Alarcón et al., 2004) e dura em média 11 min (Fuchs, 1964). É relativamente rápido, se considerarmos o número de láparos que nascem. A fêmea ingere as placentas, lava e limpa os láparos, um por um, consoante nascem. Quando termina o parto, a coelha sai do ninho, limpa-se e alimenta-se (Xu, 1996).
Um comportamento maternal adequado durante o período peri-parto é caracterizado, entre outros fatores, pela ausência ou reduzida incidência de desordens como o canibalismo (González-Redondo & Zamora-Lozano, 2008).
Após o parto, a coelha entra em lactação, que tem a duração de aproximadamente 2 meses, no caso do desmame ser feito de forma natural. Nas fêmeas simultaneamente gestantes e lactantes, a diminuição da produção de leite é mais rápida, podendo parar ao fim de 4 a 5 semanas de lactação (Mourão & Pinheiro, 2004). Durante este período a coelha amamenta os láparos com uma periodicidade circadiana e uma duração aproximada de 3 minutos/dia (González-Mariscal, 2001), geralmente durante o período noturno (Ninomiya-Alarcón et al., 2004).
7 O comportamento maternal é regulado pelo sistema neuro-endócrino. Estrogénio, testosterona, progesterona e prolactina regulam aspetos específicos da construção do ninho e promovem o aparecimento da responsabilidade maternal pela ninhada (González-Mariscal, 2001).
Uma vez que a indução do parto através da administração de hormonas exógenas causa uma modificação no perfil hormonal natural, isso pode refletir-se em alterações do comportamento maternal.
2.3 Mecanismos que desencadeiam o parto
Nenhum outro mecanismo neuro-endócrino é tão integrador e dinâmico como o que se observa no trabalho de parto e expulsão dos fetos. Uma coordenação afinada de uma série de reações fisiológicas entre os fetos, a placenta e a mãe é fundamental para o trabalho de parto culminar com o nascimento bem sucedido. Os problemas relacionados com o trabalho e parto (distócia) não são incomuns (Anónimo, 2005).
Durante a gestação o útero aumenta de volume e vai esticando gradualmente para acomodar o aumento do tamanho do(s) feto(s). A progesterona desempenha um papel fundamental na manutenção da quiescência do miométrio bem como na promoção da contração da cérvix (Duarte, S/D.).
Na última parte da gestação, os estrogénios começam a influenciar o miométrio através da estimulação da produção de proteínas contrácteis e da formação de junções gap. As primeiras aumentam o potencial contráctil do útero e as segundas facilitam-no ao permitir uma maior comunicação entre as células musculares lisas. Assim, as importantes alterações que preparam a fêmea para o parto iniciam-se bastante antes deste ter lugar (Duarte, S/D.).
A questão mais importante em relação ao parto prende-se com o fator que o desencadeia. Na maioria dos animais domésticos é aceite que a maturação do feto provoca alterações responsáveis pela indução do parto. O órgão responsável pela iniciação desse processo é o cortex adrenal fetal, com o hipotálamo e a adenohipófise a terem importantes papéis de suporte (North et al., 1973; Duarte, S/D.). Este conceito surgiu com a experiência de Liggins et al. (1967), que demonstrou que a destruição da pituitária anterior de fetos de ovino tinha como consequência o prolongamento da gestação. Na sequência desta descoberta, Drost & Holm (1968) verificaram os mesmos resultados após uma adrenalectomia fetal.
8
Na fase final de gestação há uma elevação na secreção de ACTH pela adenohipófise fetal. Pensa-se que o córtex adrenal fetal se vá tornando progressivamente mais sensível à ACTH, respondendo ao seu estímulo através da produção de cortisol fetal. O momento da maturação do córtex está sob controlo genético, sendo o momento do início da produção de cortisol característico de cada raça ou linhagem (Duarte, S/D.).
O local de ação do cortisol fetal é a placenta, estimulando a síntese de enzimas placentárias que desviam a produção de esteroides da progesterona para os estrogénios. A alteração no rácio estrogénios/progesterona tem como efeito a síntese e libertação de prostaglandinas pelo útero, em particular a PGF2α, que por sua vez induz a luteólise, a
contração do miométrio e o relaxamento do cérvix (Anónimo, 2005; Duarte, S/D.).
O processo acima descrito verifica-se em momentos diferentes da gestação, conforme a espécie. No entanto, é questionável que o eixo adenohipófise–cortex adrenal fetal tenha um papel obrigatório no desencadear do parto em todos os mamíferos, nomeadamente nos coelhos e ratos. Nestas espécies, após se proceder a uma fetotomia, observa-se que a placenta é expulsa apenas na altura normal para o término da gestação, mostrando que as hormonas fetais não desempenham um papel fundamental no desencadear do parto em coelhos e em ratos (Anónimo, 2005).
O passo determinante para o início do parto em coelhas parece estar relacionado com um processo de maturação da placenta determinado geneticamente, e que se traduz num aumento da produção de estradiol ou de prostaglandinas (Anónimo, 2005). Embora o início da síntese de PGF2α não esteja completamente entendido nos coelhos, sabe-se que o
aumento da produção desta hormona pelo útero tem um papel fundamental no término da gestação e desencadear do parto. Em várias espécies animais, incluindo o coelho, a PGF2α
foi identificada como o principal fator uterino responsável pela luteólise (Lytton & Poyser, 1982).
A PGF2α inicia a regressão do corpo lúteo (CL) antes do parto, provocando uma
queda nos níveis de progesterona plasmática. Nos coelhos, a queda da concentração de progesterona no plasma sanguíneo representa uma etapa reguladora fundamental para o inicio do trabalho de parto (Csapo, 1976). A descida dos níveis desta hormona desencadeia uma “cascata” de acontecimentos a nível fisiológico (ver Figura 1) que rapidamente conduz à expulsão dos láparos e à expressão do comportamento maternal no peri-parto.
9 Para além de causar a lise do CL, baixando os níveis de progesterona e dissipando os efeitos que esta tem na manutenção da quiescência do útero, a PGF2 aumenta a
contratilidade do miométrio, promovendo a libertação de cálcio intracelular. O cálcio libertado liga-se à actina e à miosina para iniciar a contração. As prostaglandinas também estão envolvidas diretamente no alargamento/amolecimento do canal cervical (PGE2) e provocam a secreção de relaxina pelo CL e/ou placenta (PGF2. O útero transforma-se de
órgão quiescente em órgão contráctil e a cérvix relaxa e abre-se para permitir a saída dos fetos (Anónimo, 2005; Duarte, S/D.).
Os recetores de oxitocina nas células miometriais sofrem um acréscimo notável no termo da gestação devido ao aumento dos estrogénios (Fuchs, 1964). Como o útero começa a contrair-se e os fetos descem até à cérvix, são enviados para o cérebro neurotransmissores que causam a secreção de oxitocina (reflexo de Ferguson). Só começam a libertar-se quantidades significativas desta hormona após o feto entrar no canal pélvico (Anónimo, 2005; Duarte, S/D.).
A neurohipófise materna ocasiona a secreção da oxitocina em pulsos. Esta hormona estimula a contração do miométrio, atuando em sinergia com a PGF2, propagado um
feedback positivo à medida que as contrações uterinas se tornam mais intensas, causando a produção de mais oxitocina. Embora esta hormona seja um importantíssimo auxiliar na expulsão dos fetos, a oxitocina não tem um papel preponderante no determinismo do momento do parto (Anónimo, 2005; Duarte, S/D.).
A relaxina também é uma hormona importante na preparação do parto. Esta promove o relaxamento dos ligamentos e músculos associados ao trabalho de parto, localizados no canal pélvico. Também tem um efeito pacemaker no ritmo do miométrio,
Figura 1 – Alterações nas concentrações plasmáticas de progesterona, estradiol, testosterona e prolactina durante o final da gestação, parto e lactação da coelha. Fonte: adaptado de González-Mariscal (2001).
10
coordenando as contrações induzidas pelo estradiol, prostaglandinas e oxitocina (Anónimo, 2005; Duarte, S/D.). Na coelha a relaxina atinge a maior concentração ao dia 15 de gestação, mantendo-se em valores elevados até ao momento do parto, caindo abruptamente a sua concentração após este momento (Lee & Fields, 1991).
2.4 Utilização de prostaglandinas e oxitocina em produção animal
As prostaglandinas são ácidos gordos insaturados e pertencem à família dos lípidos biologicamente ativos. São encontradas em vários tecidos e estão envolvidas em quase todas as fases de regulação endócrina das funções reprodutivas. O ácido araquidónico é o precursor mais comum das prostaglandinas mais intimamente associadas com a reprodução, principalmente a PGF2α, e a PGE2 (Boiti, 1999).
As prostaglandinas não são armazenadas nas células e a suas concentrações no espaço intercelular dependem da disponibilidade do seu percursor, o qual é considerado um fator limitante para a síntese de prostaglandinas. No entanto, hormonas e outros fatores podem promover a síntese de prostaglandinas ao ligarem-se a recetores específicos da membrana celular, e assim ativam a libertação do seu percursor (Boiti, 1999).
Entre as prostaglandinas, a PGF2α tem um papel fundamental no término da gestação
e no desencadear do parto. Em várias espécies animais, incluindo o coelho, a PGF2α foi
identificada como o principal fator responsável pela luteólise (Lytton & Poyser, 1982).
Com o avanço da tecnologia, vários análogos sintéticos da PGF2α foram desenvolvidos
pela indústria farmaceutica, com o objetivo de utilisar terapeuticas baseadas na ação luteolítica desta hormona. Entre os vários análogos sintéticos desta hormona que existem no mercado atualmente destaca-se o cloprostenol, bastante utilizado em produção animal. Esta substância é estruturalmente relacionada com a PGF2α e está comprovado que, tal como a
hormona natural, este análogo sintético também provoca a regressão morfológica e funcional do CL (Anónimo, 2012).
O cloprostenol é comercializado com o nome Estrumate®, e a sua utilização terapeutica em produção animal é recomendada para as seguintes circunstâncias: cios silenciosos; término de gestações recentes; término de gestações anormais (fetos mumificados, prolapsos vaginais, etc); endometrite; cistos luteos; indução do parto; termino da pseudogestação; tratamento de anestro de lactação; entre outros (Anónimo, 2012).
11 Quanto à oxitocina, esta é uma hormona de natureza proteica, produzida naturalmente pelo hipotálamo e armazenada na hipófise posterior ou neurohipófise. Esta hormona é libertada pela neurohipófise para a corrente sanguínea no momento do parto, promovendo o início e a manutenção das contrações musculares uterinas necessárias à expulsão dos fetos (Cross, 1958; Fuchs, 1964; Ninomiya-Alarcón et al., 2004). Para além disso, a oxitocina está envolvida noutros processos fisiológicos, desempenhando um papel importante durante a fase de amamentação, pois é a responsável pelo mecanismo de ejeção do leite durante a amamentação (Ninomiya-Alarcón et al., 2004), ligando-se às células mioepiteliais dos alvéolos e promovendo a sua contração.
A indústria farmacêutica também produz e comercializa oxitocina para utilização em medicina humana e veterinária. Este fármaco tem sido um auxiliar valioso na produção animal. Um dos produtos veterinários comercializado em Portugal que tem como princípio ativo a oxitocina é o Placentol®. A utilização deste produto é indicada para: suporte do parto normal; auxiliar do parto indolente por inércia uterina; no prolapso uterino; e na expulsão de produtos patológicos retidos no útero (retenção placentária, metrites, etc.) (Anónimo, 2010).
O Placentol® também facilita o escoamento total do leite a partir do úbere, sendo eficaz como adjuvante da terapêutica de mamites, na congestão mamária e retenção do leite (Anónimo, 2010).
Este medicamento veterinário, assim como outros que tenham oxitocina na sua composição, não devem ser utilizados durante a gestação, salvo se se pretender provocar o parto. Para além disso, a sua utilização está contra-indicada no caso de partos distócicos ou da não abertura do colo uterino (Anónimo, 2010).
2.5 Indução do parto em cunicultura (estado da arte)
Nas últimas décadas tem-se assistido ao desenvolvimento de inúmeras técnicas de controlo reprodutivo em produção animal, com o intuito de aumentar a eficiência produtiva. Uma das técnicas de controlo reprodutivo que surgiu neste contexto foi a indução do parto através de tratamentos hormonais. A sua utilização em produção animal pode justificar-se por diversas razões. Para além da possibilidade de pôr termo a uma gestação demasiado prolongada ou ligada a certos transtornos patológicos, com a utilização desta técnica minimiza-se o tempo gasto na observação da parturiente, pois permite programar a hora do parto, possibilitando o acompanhamento do mesmo (Salles et al., 1987).
12
Especificamente em cunicultura, também se assistiu a um avanço considerável das técnicas de reprodução utilizadas para auxiliar a produção. Entre elas, a inseminação artificial (IA), que começou a ser utilizada em coelhos em 1950 (Ptaszynska, 2007). Esta técnica mostrou ser muito útil em cunicultura e é largamente aplicada atualmente, permitindo a concentração das cobrições de um grande número de animais no mesmo dia. No entanto, esta técnica exige uma grande sincronização dos ciclos reprodutivos das coelhas para que se obtenham resultados satisfatórios, sem que surjam flutuações na produtividade (Alvariño & Rebollar, 1995). Para melhorar os resultados da IA nos coelhos, foram desenvolvidos métodos farmacológicos para o controle da recetividade e da ovulação (Ptaszynska, 2007).
A aplicação generalizada da IA permitiu que grande parte dos cunicultores recorram atualmente ao maneio em bandas. A necessidade de sincronização nos ciclos reprodutivos das coelhas que este tipo de maneio exige, fez com que as técnicas de indução do parto fossem trazidas também para a produção cunícola, com o objetivo de adiantar e agrupar os partos num dia pré-determinado, facilitando o maneio de controlo das ninhadas (Alvariño & Rebollar, 1995). Porém, estes tratamentos são aplicadas com menor expressividade do que os da indução da recetiviade e ovulação e geram alguma controvérsia de opiniões.
O parto em coelhas pode ser induzido recorrendo à administração de corticósteroides, de prostaglandinas, ou de oxitocina. Porém, a utilização de corticósteroides (p.ex. dexametasona) como indutores de parto em coelhas tem pouca eficácia e a sua utilização resume-se exclusivamente para fins de investigação científica (Kendall & Liggins, 1972; North et al., 1973; Challis et al., 1975). Os tratamentos com oxitocina ou prostaglandinas são os mais recomendados para indução do parto em cunicultura (Alvariño & Rebollar, 1995; Ptaszynska, 2007).
Existe uma correlação entre a atividadde uterina durante o parto e os níveis plasmáticos de oxitocina. A administração de oxitocina sintética em coelhas no termo da gestação faz aumentar a concentração plasmática desta hormona (dependo da dose administrada) e faz aumentar as contrações uterinas (Fuchs & Dawood, 1980). Obtém-se a indução do parto com uma injeção intramuscular ou intravenosa de oxitocina nos dias 30 ou 31 de gestação (18 a 24 h antes que este se produza naturalmente) (Hertelendy, 1973; Morgan, 1974; Alvariño & Rebollar, 1995; Rooney et al., 1997). O parto inicia-se 2 a 3 min após a administração do fármaco, porém a resposta é bastante variável (Alvariño & Rebollar, 1995).
Associado aos tratamentos de indução do parto com oxitocina, alguns autores relataram taxas de mortalidade neonatal relativamente elevadas, na ordem dos 5/6%
13 (Alvariño & Rebollar, 1995; Ptaszynska, 2007), e alta incidência de distócia (Ptaszynska, 2007).
O coelho tem sido um dos modelos animais de eleição para o estudo do parto em medicina humana (Ninomiya-Alarcón et al., 2004; Mitchell & Taggart, 2009), por isso existem bastantes publicações desta área relativas a ensaios de parto utilizando este animal. Entre essas publicações existem alguns ensaios de indução do parto com oxitocina (Hertelendy, 1973; Rooney et al., 1997), pois esta hormona é muito utilizada em medicina humana com essa mesma função.
Apesar de bastante estudada noutras espécies, a utilização de oxitocina em coelhos é pouco aceite. Porém, é utilizada como procedimento rotineiro em alguns laboratórios como alternativa à cesariana, para obtenção de láparos recém-nascidos não alimentados, necessários à investigação científica (Morgan, 1974).
Para além das publicações na área da medicina ou outras áreas científicas, a informação sobre uso de oxitocina para indução do parto como uma técnica auxiliar à produção cunícola é bastante escassa. Mesmo assim, este procedimento é recomendado por alguns autores (Alvariño & Rebollar, 1995; Ptaszynska, 2007) e é utilizado com sucesso em algumas cuniculturas (Ptaszynska, 2007).
Quanto à utilização de PGF2α ou dos seus análogos sintéticos, estes são os
tratamentos hormonais mais aceites e mais utilizados em cunicultura para indução e sincronização dos partos (Boiti, 1999; Lavara et al., 2002). À semelhança dos outros mamiferos, nos coelhos a PGF2α também tem um efeito luteolítico (Lytton & Poyser, 1982;
Boiti, 1999) induzindo uma queda precoce nos níveis de progesterona quando administrada nos últimos dias antes do parto e, consequentemente, reduzindo a duração da gestação (Rebollar et al., 1997). Não se conhecem efeitos secundários prejudiciais resultantes da utilização destes tratamentos (Ubilla et al., 1988; Ptaszynska, 2007).
O adiantamento que os tratamentos com prostaglandinas provocam na queda dos níveis de progesterona plasmática faz com que se dissipe mais rápidamente o efeito inibitório que este esteroide produz na libertação de gonadotropinas. Isto provoca um adiantamento na produção de FSH e LH, favorecendo o crescimento dos folículos ováricos (Rebollar et al., 1989). São vários os autores que descrevem os efeitos benéficos que a utilização destes tratamentos causa na fertilidade pós-parto das coelhas (Rebollar et al., 1989; McNitt et al., 1997).
14
A dimensão da ninhada ao nascimento parece não ser afetada por este tipo de tratamentos (Rodriguez et al., 1984; Negatu et al., 1998; Rebollar et al., 1997). Já o peso individual dos láparos resultantes de partos induzidos com PGF2α é geralmente inferior em
comparação com os láparos resultantes de um parto não induzido, devido à redução no tempo de gestação (Partridge et al., 1986; Ubilla & Rodriguez, 1989; Lavara et al., 2002). Apesar disso, quando a administração destes tratamentos é feita no dia 28, 29 ou 30 de gestação obtêm-se maioritáriamente ninhadas viáveis (Alvariño & Rebollar, 1995). Alguns autores relatam inclusivé que a mortalidade neonatal dos láparos resultantes dos partos induzidos com PGF2α é inferior à normal, sugerindo que a causa para esta redução na
mortalidade seja um maior controlo pelos operadores no período de partos (facilitado pela maior sincronização) e/ou uma maior facilidade de parto das coelhas, que poderão ter partos mais rápidos e menos stressantes devido ao peso individual dos láparos ser inferior (Rebollar et al., 1989).
A utilização de análogos sintéticos da PGF2α, como o cloprostenol, o fenprostalene ou o
etiproston, permite a utilização de dosagens bastante inferiores do que as dosagens necessárias para induzir o parto com PGF2α natural (Alvariño & Rebollar, 1995). Alguns
estudos concluíram que a utilização de análogos sintéticos tem os mesmos efeitos na descida de progesterona do que a PGF2α natural, causando respostas semelhantes na
indução do parto (Rebollar et al., 1997; Negatu et al., 1998). Já McNitt et al., 1997 dizem poder haver algumas diferenças em relação ao número de láparos nascidos vivos entre a utilização de PGF2α natural ou o análogo sintético cloprostenol, no entanto a utilização de
15
3 Trabalho experimental
3.1 Objetivos
O estudo teve como principal objetivo avaliar qual o efeito que a indução de parto com cloprostenol (análogo sintético da PGF2α) ou oxitocina tem na performance
produtiva de coelhas quando o tratamento é repetido várias vezes nos mesmos animais, de forma a perceber se haverá vantagens em utilizar este tipo de tratamentos de forma rotineira em contexto de produção.
Também se pretendeu perceber qual a melhor altura para a administração de cloprostenol como indutor do parto, se ao 28º ou 29º dia de gestação, uma vez que havia a necessidade de comparar os diferentes protocolos de administração.
Averiguar se o tratamento com oxitocina seria uma alternativa viável, uma vez que a utilização desta hormona como indutor do parto está mais estudada para coelhos em condições de laboratório e em condições de emergência, do que num contexto de rotina planificada, sendo interessante fazer uma análise comparativa da sua utilização em relação aos tratamentos com efeito luteolítico, mais comummente utilizados.
Para além disso, pretendeu-se mencionar as alterações em termos de mão de obra que advêm da utilização deste tipo de tratamentos, ou seja, se os tratamentos facilitam ou dificultam o maneio durante a época de partos.
16
3.2 Material e Métodos
3.2.1 Animais testados
O ensaio teve início com 75 coelhas nulíparas de uma estirpe comercial resultante do cruzamento das raças Neozelandês x Californiano (NZxC), habitualmente utilizada em cunicultura como linha maternal para obtenção da geração de abate para produção de carne.
No início do ensaio, os animais foram identificados individualmente através de um número tatuado no pavilhão auricular. Procedeu-se à primeira inseminação artificial às 18 semanas de idade, sendo o peso médio das coelhas nessa altura de 3,5 Kg.
3.2.2 Profilaxia
Toda a sala de ensaio foi higienizada e desinfetada antes da entrada dos animais, procedendo-se a limpezas regulares durante todo o ensaio. As caixas ninho utilizadas foram removidas após cada desmame e devidamente lavadas e desinfetadas antes de nova utilização. Esporadicamente procedeu-se à queima dos pelos que se acumulavam nas jaulas.
Foi utilizado aparas de madeira esterilizadas como substrato para o ninho. Os láparos nascidos mortos e as placentas foram removidos imediatamente após o parto. Foram feitas rondas diárias durante todo o ensaio de forma a retirar os animais mortos e proceder à eliminação dos cadáveres. Esporadicamente foram realizados tratamentos com enxofre em pó, polvilhado sobre os ninhos, de forma a controlar o desenvolvimento de fungos (p.ex. tinha).
Os animais receberam vacinação e tratamentos antiparasitários em conformidade com o programa padrão utilizado em cuniculturas comerciais, nomeadamente a vacinação contra a Mixomatose e a Doença Hemorrágica Viral, que foram administrados antes da primeira inseminação.
No período de lactação do 2º ciclo reprodutivo procedeu-se a um tratamento com antibiótico, adicionado à água de abeberamento, de forma a controlar o surgimento de um surto de enteropatia mucoide.
17
3.2.3 Período de ensaio
O ensaio decorreu na cunicultura experimental do Departamento de Zootecnia na Universidade de Trás-os-Montes e Alto Douro, entre outubro de 2011 e maio de 2012, tendo uma duração de 6 meses, aproximadamente. Iniciou-se aquando da IA do primeiro ciclo reprodutivo das coelhas e finalizou-se no desmame do quarto ciclo. Foram por isso controlados 4 ciclos reprodutivos completos, desde a IA até ao desmame, sendo o tratamento atribuído a cada animal repetido nos sucessivos ciclos (exceto no caso de animais que não ficaram gestantes em algum dos ciclos).
3.2.4 Alojamento
O pavilhão onde decorreu o ensaio era utilizado exclusivamente como maternidade, cumprindo as regras básicas de higiene, segurança e bem-estar animal necessárias à produção cunícula (ver Figura 2).
As reprodutoras foram alojadas individualmente em jaulas (40x30x50 cm; LxAxC) com ninho, onde permaneceram durante todo o ensaio, sendo os láparos retirados para outro pavilhão aquando do desmame.
Durante o ensaio o pavilhão esteve à temperatura ambiente e sujeito a ventilação natural suplementada com um extrator. No entanto, no segundo e terceiro ciclos reprodutivos, por se terem registado temperaturas bastante baixas, foi reduzida a ventilação do pavilhão e colocado um sistema de aquecimento elétrico durante as primeiras duas semanas após o parto, de forma a garantir um maior conforto térmico para as ninhadas.
Os animais foram sujeitos a um fotoperíodo artificial de 12 h luz /12 h escuro, sendo este aumentado para 16 h luz /8 h escuro durante os 7 dias precedentes à IA. Após o parto, o período de luz era reduzido 1 hora por dia até alcançar as 12 horas diárias.
Figura 2 – Pavilhão onde decorreu o ensaio. Fonte: autor.
18
3.2.5 Alimentação
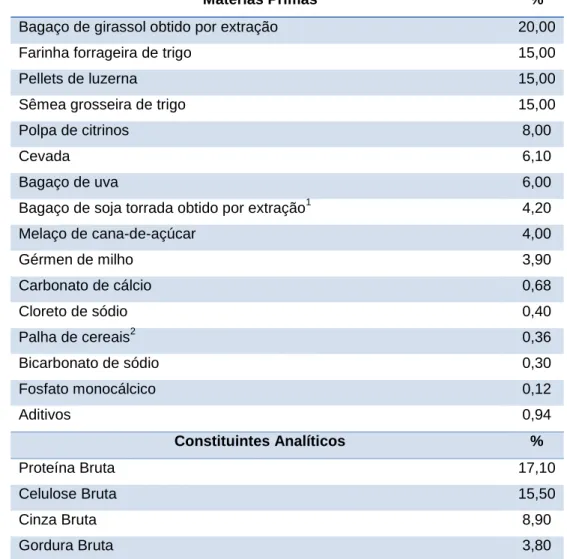
Os animais foram sustentados com um alimento composto completo de uma marca comercial destinado a coelhas reprodutoras (Fazcunir®; Nanta), cuja composição se pode ver na Tabela 1. Este alimento foi distribuído ad libitum às coelhas lactantes. Nas coelhas não lactantes foi feito um flushing alimentar antes da IA e posteriormente o alimento foi restringido a 120 g diárias até 1 semana antes do parto, sendo distribuído novamente ad libitum na última semana de gestação. Os láparos foram alimentados com o mesmo alimento até à data do desmame. A água de abeberamento foi também disponibilizada permanentemente.
Tabela 1 – Composição do alimento fornecido durante o ensaio.
Matérias Primas %
Bagaço de girassol obtido por extração 20,00
Farinha forrageira de trigo 15,00
Pellets de luzerna 15,00
Sêmea grosseira de trigo 15,00
Polpa de citrinos 8,00
Cevada 6,10
Bagaço de uva 6,00
Bagaço de soja torrada obtido por extração1 4,20
Melaço de cana-de-açúcar 4,00 Gérmen de milho 3,90 Carbonato de cálcio 0,68 Cloreto de sódio 0,40 Palha de cereais2 0,36 Bicarbonato de sódio 0,30 Fosfato monocálcico 0,12 Aditivos 0,94 Constituintes Analíticos % Proteína Bruta 17,10 Celulose Bruta 15,50 Cinza Bruta 8,90 Gordura Bruta 3,80 1
produzido a partir de sementes de soja geneticamente modificadas;
2
19
3.2.6 Tratamentos e delineamento experimental
As reprodutoras foram sujeitas a um ritmo de produção semi-intensivo, com ciclos em banda única de 42 dias, sendo inseminadas 11 dias após o parto. O desmame dos láparos realizou-se quando estes tinham 35 dias de idade. Para estimular a recetividade das coelhas utilizou-se a técnica da amamentação controlada até ao momento da IA, em que o acesso da coelha aos láparos só era permitido uma vez por dia, com intervalos de 24 h entre amamentações, sendo este intervalo aumentado para 28 h/29 h no dia da IA. Foi também administrada eCG, dois dias antes de cada inseminação, a todas as reprodutoras. A IA foi feita entre as 10:00 h e as 11:00 h da manhã em todos os ciclos reprodutivos.
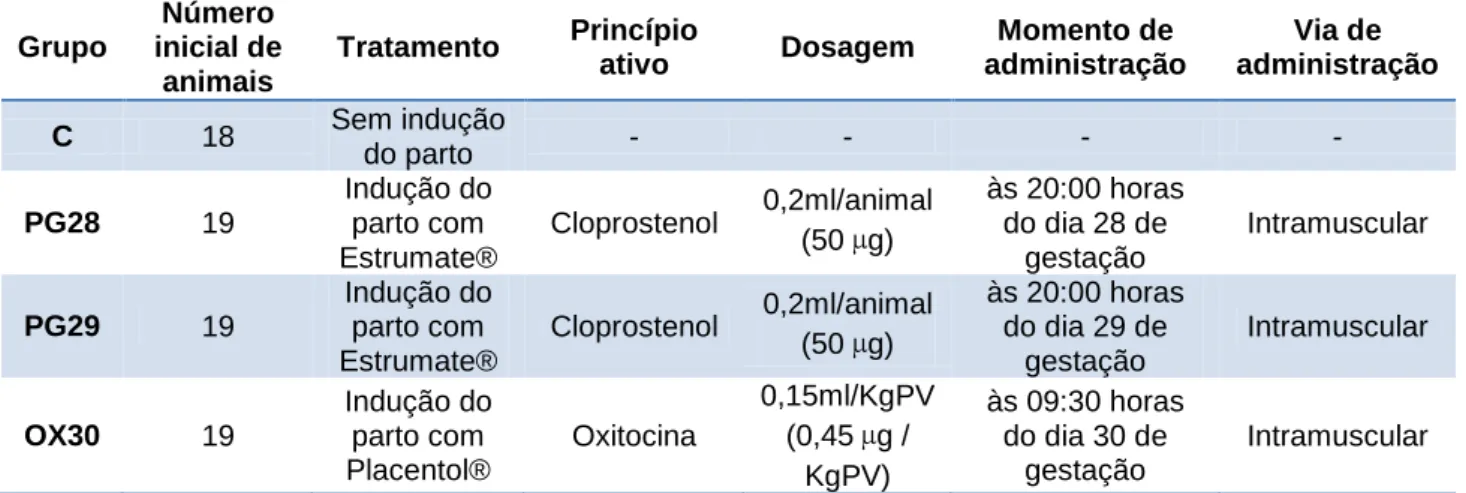
Vinte e oito dias após a primeira inseminação artificial os animais foram distribuídos em 4 grupos: grupo controlo (C) em que não foi induzido o parto; grupo PG28, em que foi administrado cloprostenol - análogo sintético da PGF2α (Estrumate®), ao dia 28 de gestação;
grupo PG29, em que foi administrada a mesma substância, mas ao dia 29 de gestação; e grupo OX30, em que foi administrado oxitocina (Placentol®) ao dia 30 de gestação (ver Tabela 2). As dosagens utilizadas foram calculadas seguindo as recomendações para outras espécies presentes na bula dos fármacos, uma vez que não estavam presentes recomendações para coelhas, estimando-se a dose a partir do peso medio das reprodutoras.
Tabela 2 – Caracterização dos grupos de ensaio Grupo Número inicial de animais Tratamento Princípio ativo Dosagem Momento de administração Via de administração C 18 Sem indução do parto - - - - PG28 19 Indução do parto com Estrumate® Cloprostenol 0,2ml/animal (50 µg) às 20:00 horas do dia 28 de gestação Intramuscular PG29 19 Indução do parto com Estrumate® Cloprostenol 0,2ml/animal (50 µg) às 20:00 horas do dia 29 de gestação Intramuscular OX30 19 Indução do parto com Placentol® Oxitocina 0,15ml/KgPV (0,45 µg / KgPV) às 09:30 horas do dia 30 de gestação Intramuscular
A distribuição dos animais pelos quatro grupos foi feita de forma a que houvesse uniformidade entre eles, tendo em conta 3 fatores:
- estado fisiológico após a primeira IA – gestante ou não gestante (grupos uniformes nesta relação);
- peso vivo das coelhas no dia da IA e 28 dias após;
20
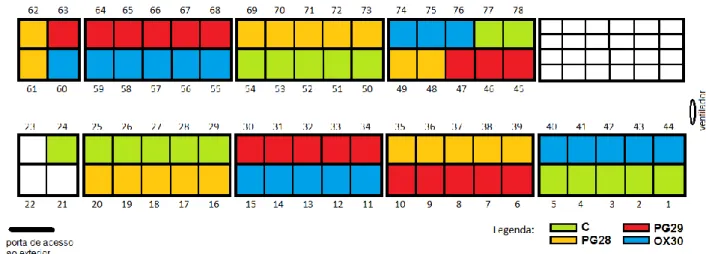
Os animais foram alojados nas jaulas de modo a que os diferentes tratamentos ficassem uniformemente distribuídos pelo pavilhão onde decorreu o ensaio (ver Figura 3), evitando-se assim que possíveis variações ambientais numa determinada zona do pavilhão tivessem influência nos resultados de apenas um dos grupos. Para facilitar o maneio a aplicar, cada jaula foi identificada com uma mola com a cor do tratamento atribuído a esse animal.
Figura 3 – Esquema da distribuição dos grupos pelas jaulas de ensaio. Fonte: autor.
Procedeu-se ao diagnóstico de gestação por palpação abdominal entre os dias 20 a 25 após a IA, de forma a distribuir caixas ninho e aparas de madeira por todas as coelhas gestantes.
Durante as épocas de parto, a partir do dia 28 de gestação e até todos os animais gestantes terem parido, foram feitas rondas frequentes aos ninhos durante o período diurno (entre as 8:00 h e as 20:00 h), intervaladas cerca de uma hora, para registo dos dados de parto. No dia de maior ocorrência de partos, 30 dias após IA, as rondas iniciaram-se às 6:00 h e prolongaram-se até às 22:00 h. No período noturno os animais não foram controlados, sendo os partos ocorridos nesta altura registados na primeira ronda da manhã.
Após cada parto foi feita a homogeneização das ninhadas, não havendo distinção entre os grupos de onde provinham os láparos. O objetivo foi ter ninhadas semelhantes entre os tratamentos ao início da lactação, para que fosse possível identificar um possível aumento na produção leiteira ou alterações comportamentais das progenitoras que pudessem ter reflexos nas performances de crescimento dos láparos.
21
3.2.7 Determinações
Com vista a avaliar quais as principais alterações que os tratamentos causam nos animais e a forma como estes afetam a produtividade, foram estudados parâmetros que podem ser indicativos de alterações na performance reprodutiva das coelhas e na performance de crescimento das ninhadas. Para tal realizaram-se as seguintes medições: Para analisar as performances das reprodutoras:
Fertilidade (nº coelhas que pariram/nº coelhas inseminadas); Duração da gestação;
Intervalo entre a administração do fármaco e o parto; Duração do parto;
Incidência de partos interrompidos;
Incidência de construção do ninho (presença ou ausência de pelo no ninho); Local de parto (ninho, jaula ou ambos);
Prolificidade:
Número de láparos nascidos total; Número de láparos nascidos vivos;
Percentagem de láparos nascidos mortos;
Peso da ninhada ao nascimento (antes da homogeneização); Incidência de canibalismo;
Incidência de amamentação forçada nos primeiros dias após o parto; Ingestão média diária de alimento;
Peso da coelha: Após o parto (d0);
No momento da IA (d11); Ao 18º dia após o parto (d18); Ao desmame (d35);
Variação do peso; Mortalidade;
Para analisar as performances das ninhadas e dos láparos: Peso da ninhada:
Após homogeneização (d0); Aos 18 dias de idade (d18); Ao desmame (d35);
22
Ganho médio diário; Mortalidade;
Para analisar a produtividade:
Quilogramas de láparo desmamado por coelha desmamada; Quilogramas de láparo desmamado por coelha parida; Quilogramas de láparo desmamado por coelha inseminada.
A fertilidade das reprodutoras foi calculada através da relação entre o número de partos e o número de fêmeas inseminadas e foram consideradas apenas as fêmeas que estavam vivas no momento do parto.
O intervalo entre a administração do produto e o parto foi determinado através da subtração da hora a que foi registado o início do parto (expulsão do primeiro láparo) à hora a que tinha sido administradotado o tratamento hormonal correspondente a cada grupo.
Quanto ao parâmetro duração do parto, corresponde ao intervalo de tempo entre o início da expulsão dos láparos e o momento em que se detetou que todos os láparos já tinham sido paridos. Nas reprodutoras que pariram na ausência do operador foi registado como hora do parto o momento em que se detetou a ocorrência do mesmo. Nestes animais, aos que aparentemente tiveram um parto normal (não interrompido) foi atribuído um valor médio de 11 min de duração do parto. Para os que apresentaram um parto interrompido (não concluído no momento da deteção) foi registada como hora de inicial a hora de deteção do parto, sendo registado como hora de final do parto o momento em que se detetou que todos os láparos já haviam sido paridos.
Foram considerados nados mortos, todos aqueles que se encontravam efetivamente sem vida no momento de deteção do parto. Alguns deles poderão ter nascido com vida, porém terão sido vítimas de uma mortalidade muito precoce no período periparto (aumentada por exemplo nos partos que ocorreram nas jaulas).
O peso e dimensão da ninhada são referentes aos láparos que se encontravam vivos em cada controlo. As ninhadas foram pesadas no seu todo, calculando-se o peso médio do láparo através da divisão do peso da ninhada pelo número de láparos no momento da pesagem.
Devido à mortalidade, infertilidade ou morte de toda a ninhada, o número de coelhas e ninhadas controlado mudou em diferentes momentos do ensaio, não sendo a amostragem exatamente igual para todos os grupos.
23
3.2.8 Análise de dados
A análise estatística dos dados obtidos no ensaio foi realizada recorrendo ao software estatístico JMP® 7.0. e considerou-se que haveriam diferenças significativas para valores de P < 0,05.
Foi feita uma análise de variância para os seguintes parâmetros: duração da gestação, intervalo entre a administração do fármaco e o parto, duração do parto, prolificidade, peso da ninhada ao nascimento, ingestão média diária, peso da coelha, peso da ninhada, ganho médio diário, mortalidade dos láparos e produtividade. Nos parâmetros em que se observaram diferenças significativas fez-se a comparação múltipla de médias através do teste t-Student.
Para comparar os parâmetros taxa de fertilidade, incidência de partos interrompidos, local de parto, incidência de construção do ninho, incidência de canibalismo, incidência de amamentação forçada e mortalidade das reprodutoras procedeu-se a uma análise de frequência pelo teste de Pearson (Qui-quadrado).
Embora o procedimento experimental se tenha realizado da mesma forma nos 4 ciclos reprodutivos em estudo, os dados referentes ao primeiro ciclo não foram incluídos na análise estatística, isto porque no primeiro ciclo reprodutivo não houve nenhum tratamento prévio de indução de parto que pudesse ter influenciado os dados da fertilidade ou da prolificidade referentes a esse período. Este primeiro ciclo também serviu como período de aclimatação, permitindo a habituação dos animais aos operadores e ao maneio do ensaio, reduzindo assim o impacto do stresse nos resultados obtidos ao longo dos 3 ciclos seguintes.
Os dados de animais que não ficaram gestantes em algum dos ciclos ou que pariram antes da hora de administração do tratamento foram contabilizados na análise, pois numa situação real estes casos também aconteceriam, devendo por isso ser tidos em conta para que se obtenham resultados mais representativos em relação à produtividade em condições de produção.