The above publications are included at the end of this thesis with the permission of the copyright holder, Oxford University Press (I, II) and under the terms of the Creative Commons CC-BY license (III). The author and Sergei Suprun performed the majority of the measurements with the help of Sarita Keski-Saari.
Bridging northern plant ecophysiology with the capacity to
Relatively recent Common Garden (CG) studies in North America (Gornall and Guy 2007, McKown et al. 2014, Kaluthota et al. However, leaf longevity may determine growing environment more than latitude per se (Reich et al. 1996).
Silver birch as a model species for ecological changes
Survival of northern birches in southern conditions is much more uncertain, as the increased temperature sum and insect herbivory in the south can become additional stressors, which in principle can result in lower survival rates, especially if these interact with poor soil nutrient status (Heimonen 2015, Possen et al. 2021 ). This so-called critical night length requirement is lower the further north the provenance originates from (Viherä-Aarnio et al. 2006), also for species other than birch (Mølmann et al. 2006, Luquez et al. 2008), and in CG studies northern silver birch provenances have earlier cessation of height growth than southern regardless of the latitude at which they have been transplanted (Possen et al. 2021).
Measuring CO 2 assimilation and photosynthesis connects
Gas exchange (GE)
Gas exchange can be measured with an open configuration gas exchange system (in this thesis a LI-6400, Li-Cor, Inc., USA). Furthermore, with the LI-6400 the temperature of the measuring head or blade can be accurately monitored and adjusted in several ways.
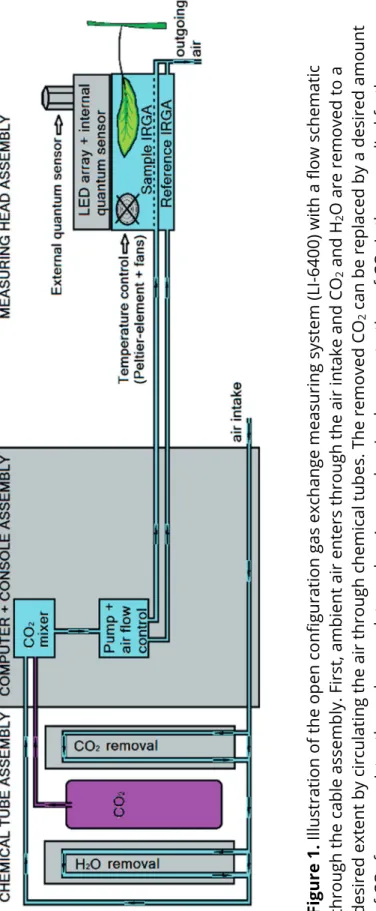
Chlorophyll fluorescence (Chl-F)
It can be measured on scales ranging from (smaller than) a cell to entire ecosystems (from . satellites). With different techniques and outcomes, Chl-F can be measured from either the light- or dark-adapted conditions.
Aims and objectives
This thesis aims to characterize some of the most important photosynthetic, growth and biomass allocation traits of silver birch provenances from a latitudinal gradient (60 – 67°N) across Finland. The provenances were grown in equal conditions with optimal light, temperature, humidity and nutrients to study differences between provenances, free from the confounding effects in the field.
Plant material and experimental setups
YFD leaves are leaves of the same age, usually around the 4th to 5th leaf from the plant tip. However, the room studies used first-year plants and all measurements can be considered to have been made on LS leaves (Fig. 1 in II and Fig. 2 in III).

Gas exchange measurements
Schematic view of the leaf types measured both in the common garden (CG) and room experiments. The leaf was kept at constant temperature (except for temperature response curves) by keeping the cooling block temperature either at 23°C (in the field) or at 20 – 22°C (in the chambers). PPFD provided to the leaf was 1800 μmol photons m-2 s-1 (except for the variable PPFD provided to the leaf in the light response curves in II, or the measurements in III).
In (II), snapshot GE was monitored from all plants throughout the lifetime of a single leaf (YFD1) from the moment it was fully expanded and. Since there were more measurements in the first data set, it was used in the main text in (III).
Growth, leaf longevity, structural leaf traits and biomass
In the field, data on number of leaves, total leaf area and growth end DOY were obtained as a subset of data collected by Heimonen et al. 2017), used in the supplementary materials (Supplementary Fig. S6 of I). In (III), the absolute height growth rate (AGR) was additionally calculated as the height gain since the previous measurement. Stomatal density was calculated as stomata per cm2, and the total number of stomata (per leaf) was calculated by multiplying the density by the total leaf area.
The resulting data provide a representative plant average of leaf DW, leaf area, and leaf LMA of different ages. In (III), LMA was calculated by dividing leaf litter dry mass by leaf dry litter area.
Chlorophyll content and chlorophyll fluorescence kinetics
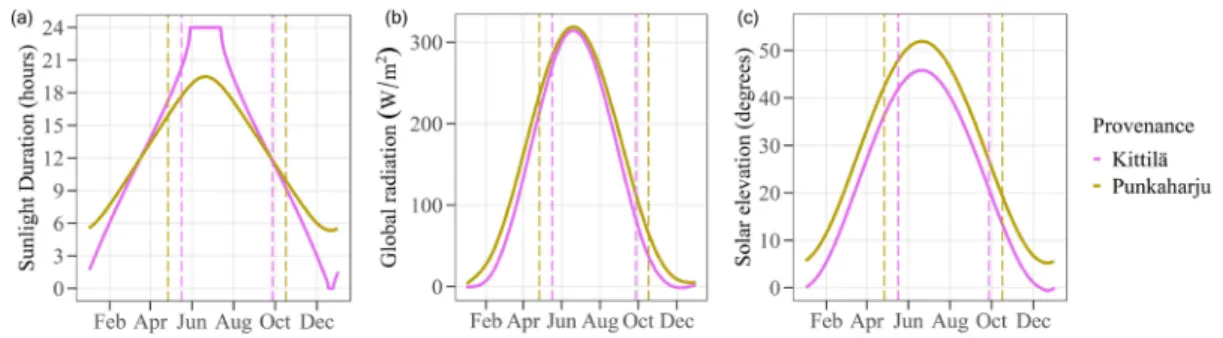
Under field conditions in (I), the measured leaves were on branches that were usually south-facing, and as such received equally bright, but third-year silver birches. Chlorophyll fluorescence kinetics were measured from 23 plants once in (II) with Handy FluorCam FC 1000-H (Photon Systems Instruments, Czech Republic) using the Fv/Fm protocol in the FluorCam 7 software (version 1.0.24.5). The 'measurement flash' duration was 20μs - this is non-actinic red light (620nm) and is only used as a visual aid to fix the blade in the dark.
The leaf was measured while attached to the plant, with as much leaf surface as possible and lying as flat as possible on the instrument head to ensure an even reading. The results obtained from the measurements are raw fluorescence data which must be post-processed in the FluorCam software.
Statistical analyses
Most of the time this involved a selected set of variables, but in some cases (especially for non-linear models) the process started with a "full" model including most of the available predictors, and then the predictors were iteratively dropped to achieve the best fit. For all models in all articles, visual fit diagnoses were preferred over statistical tests. In all cases, the modified models were then compared to the less modified model using Akaike.
In some cases it was then necessary to change the fixed/arbitrary structure (or other parts) of the model and repeat the process again. Models were estimated with maximum likelihood (ML) during model building, but final reported regression coefficients and p-values are from models estimated with limited maximum likelihood (REML).
Main findings
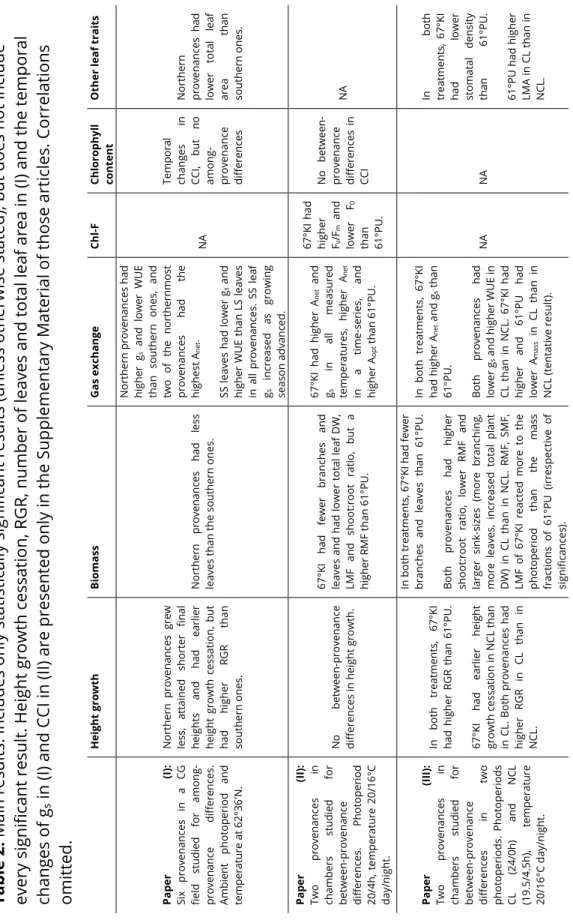
Interestingly, 67°KI had lower stomatal density than 61°PU (III) and stomatal density and the total number of stomata was negative. 67°KI had fewer branches and leaves and had lower total leaf DW, LMF and shoot:root ratio, but a higher RMF than 61°PU. 67°KI had higher Anet and gs in all measured temperatures, higher Anet in a time series and higher Aopt than 61°PU.
67°KI had a higher and 61°PU had a lower Amass in CL than in NCL (preliminary result). When 67°KI was subjected to NCL, it stopped growing in height earlier than in CL(III).
Differences in the growth performance and biomass
67°KI stopped height growth earlier than 61°PU also in the NCL treatment (III), but when the photoperiod was relaxed (section II and CL in III), it clearly did not stop height growth earlier. High allocation to roots may be adaptive in the north (Reich et al. 2014, Zadworny et al. 2016) and will divert resources away from aboveground biomass. In (I) and (III), northern provenances consistently had higher relative height growth rates than southern ones and RGR was calculated in several different ways, increasing confidence in the observation.
Third, in the CL-treatment (III), the linear regression line of 67°KI crossed the regression line of 61°PU (Supplementary Fig. S2b III), and a similar phenomenon was observed in (II) when the model was regressed throughout the growth duration (Fig. 2c in II). In the CL-treatment (III), the northern Ki27 genotype expressed a slender phenotype with good height growth but very little branching and in parallel relatively little leafing (Supplementary Fig. S1 of III).

Photosynthetic gas exchange and chlorophyll fluorescence of
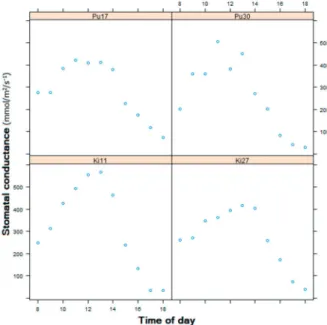
The generally low gs observed for both origins in chambers (II, III) could be attributed either to micropropagated plants having low gs or to a growth thinning effect. It is possible that chamber-grown leaves have the same total gs as in the field, but less per surface area, since plants tend to have large leaves in the first year. In the field, WUE was positively related to temperature variables such as the temperature of the warmest month, July, and negatively related to variables related to snow depth and snow duration (I).
A high WUE may be important during dry spells, and a low WUE in the warm north may pose a future threat depending on changes in precipitation. If leaves further down the stem indeed tend to have lower gs also in mature trees, such an effect was either not apparent in the data or was not strong enough to confound the leaf type effect.
Other leaf traits
The mass-based correlation was expected, as mass-based values of net photosynthesis are more often correlated with LMA compared to area-based values (Wright et al. 2004, Tholen et al. 2012). Typically, both evergreen and deciduous species show reduced mass and surface photosynthesis with increased leaf longevity (Kikuzawa Reich et al. 1992, Wright et al. 2004), but no such relationships were present here (III). Although warming is expected to increase silver birch leaf longevity (Mäenpää et al. 2011), it is not certain.
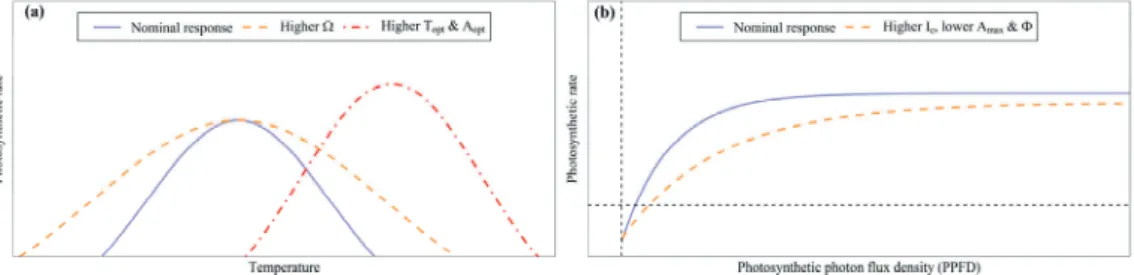
For example, mesophyll conductance can be altered in leaves with modified thickness, which can strongly influence gas diffusion to carboxylation sites (Wright et al. Such intra-leaf gradients can prevent complete light saturation even at very high light levels (Valladares et al.). . 2012).
How will silver birch cope with the changing climate?
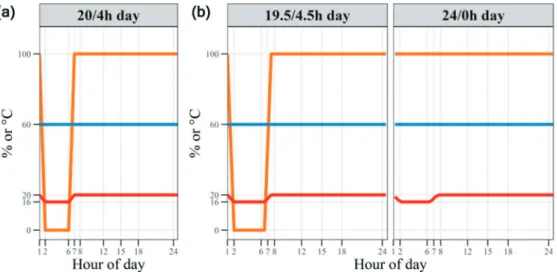
Insect herbivore pressure on northern silver birch is currently generally low (Silfver et al. 2020), but is expected to increase in the future (Heimonen 2015). For example, thick and heavy leaves (high LMA) can adapt to the cold (Poorter et al. 2009). We also used the same provenances (Tenkanen et al. 2020b) but subjected them to two different photoperiods.
Furthermore, a realistic thermoperiod may introduce some circadian rhythmicity, which is otherwise lacking in the CL treatment (Velez-Ramirez et al. 2011). Packages used included nlme (for mixed models) (Pinheiro et al. 2020), lme4 (for generalized mixed models) (Bates et al. 2015), motor (for testing oplme4 models) (Fox and Weisberg 2019), emmeans (for calculation) of mean estimates, confidence intervals and post-hoc pairwise comparisons) (Lenth 2020) enggplot2 (for graphics) (Wickham 2016). Packages used included nlme (for mixed models) (Pinheiro et al. 2020), lme4 (for generalized mixed models) (Bates et al. 2015), motor (for testing oplme4 models) (Fox and Weisberg 2019), emmeans (for calculation) of mean estimates, confidence intervals and post-hoc pairwise comparisons) (Lenth 2020) enggplot2 (for graphics) (Wickham 2016).
For deciduous species, leaf longevity typically declines toward cooler (northward) temperatures in species comparisons (Kikuzawa et al. 2013).