A partir de la description du comportement et de l'organisation des neurones dans différentes zones du cerveau, notamment des neurones impliqués dans la perception visuelle, nous définissons la notion de répertoire, qui sera le concept de base de notre modèle. Sur cette voie, nous sommes amenés à poser le problème de la stabilisation des systèmes d'annuaires et à le résoudre partiellement.
Perception et invariants perceptifs
En abordant ce problème précis, qui est au cœur de toute théorie de la perception, nous espérons pouvoir contribuer, certes de manière très limitée mais instructive, à une compréhension du problème plus général et plus vaste de la perception. , qui ne se fait pas elle-même n'est qu'un aspect très fragmenté de ce qui constitue l'un des plus grands défis de la science contemporaine : comprendre notre cerveau. Comme nous l'avons noté, tous les systèmes perceptifs sont affectés par ce problème d'invariance ; cependant, nous nous concentrerons essentiellement sur le système visuel, qui a été le plus étudié jusqu'à présent, sans oublier le fait que le principe explicatif doit dépasser les spécificités de la vision pour s'appliquer à tous les autres sens.
La théorie de Descartes
Il suppose alors que par ces ajustements la glande pinéale à la fois se rapproche ou s'éloigne dans sa cavité : ainsi si l'objet est éloigné, la glande pinéale, en s'approchant, recevra une empreinte de la même taille que si l'objet étaient près de l'œil et la glande loin de la paroi de la cavité où se termine le nerf optique. En effet, à partir des lois de la mécanique et de l'optique géométrique (qui, ne l'oublions pas, étaient les seules lois physiques connues à l'époque), Descartes forgea une théorie cohérente de la perception, compatible avec les quelques faits expérimentaux dont il put disposer.
Quelques repères historiques
Mais en même temps, un courant diamétralement opposé va naître : la théorie de la Gestalt ou théorie des formes. C'est un exemple frappant de l'absence d'invariance de taille aux premiers âges de la vie.
La recherche contemporaine
Mais c'est dans le domaine de la neurophysiologie que les progrès ont été les plus remarquables.1935 au niveau de la rétine par Hartline (1935), cette méthode conduira à la notion de champ récepteur du neurone (pour la rétine Kuffler, 1953, et pour le cortex visuel Hubel et Wiesel, 1959). Leur relation avec le système visuel est plus ou moins étroite, mais la plupart des concepts issus de ces recherches se sont avérés d'une importance primordiale pour les théories de la perception.
L’état du problème des invariants perceptifs
La vieille tradition de la chaîne S-R est abandonnée ; toute perception provient d'une interaction entre les stimuli et l'activité propre de l'organisme Comme le dit Von Holst. Dans ces circonstances, on peut comprendre une tendance très nette, comme dans d'autres domaines de la psychologie, à renoncer aux théories générales pour ce que Dunham (1977) appelle des "théories miniatures", des modèles fragmentaires qui ne rendent compte que d'un nombre limité de faits expérimentaux.
Plan de cette étude
Un autre type de modèle intéressant du point de vue des invariants perceptifs a connu un certain succès ces dernières années : le cortex visuel fabriquerait une sorte d'« hologramme » qui analyserait l'image en termes de fréquences spatiales (Maffei, 1978). Cette théorie ne prétendra pas résoudre complètement le problème, mais simplement présenter un nouveau point de vue sur cette question, et nous discuterons de l'importance de cette approche en la comparant à diverses théories contemporaines de l'invariance perceptive.
ORGANISATION TOPOLOGIQUE DES RÉGIONS CÉRÉBRALES
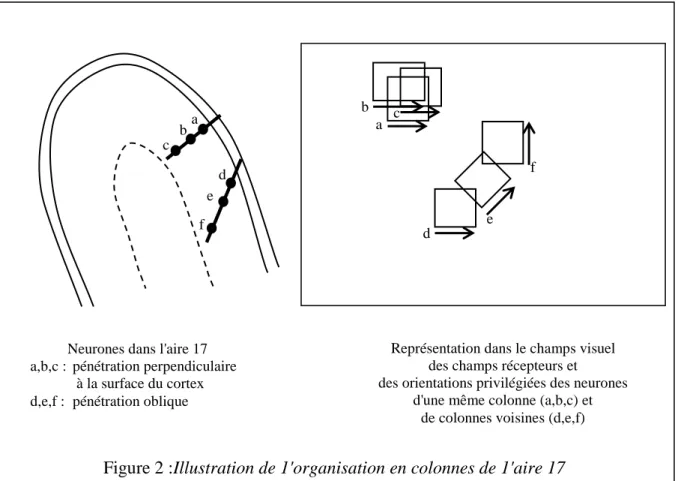
- L'organisation en colonnes du cortex visuel
- Multiplicité des représentations sensorielles et motrices
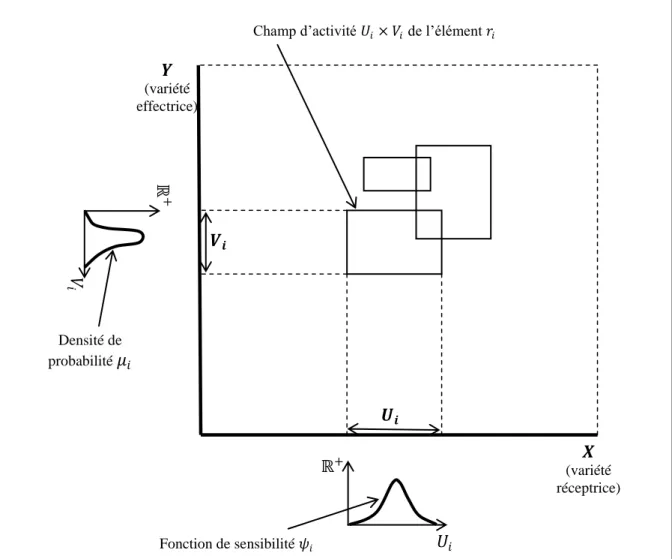
- La notion de répertoire
- Variétés réceptrices et champs récepteurs généralisés
- Position du corps et groupe moteur
- Hiérarchie des répertoires
Un répertoire sensible à plusieurs de ces propriétés aura une variété réceptrice qui sera le produit cartésien des variétés définissant chaque propriété et pourra donc être munie de la structure de la variété produit. Le champ récepteur de tout neurone du répertoire fait donc en fait partie du faisceau tangent de la variété réceptive.

PLASTICITÉ DU SYSTÈME NERVEUX
- Plasticité de la perception
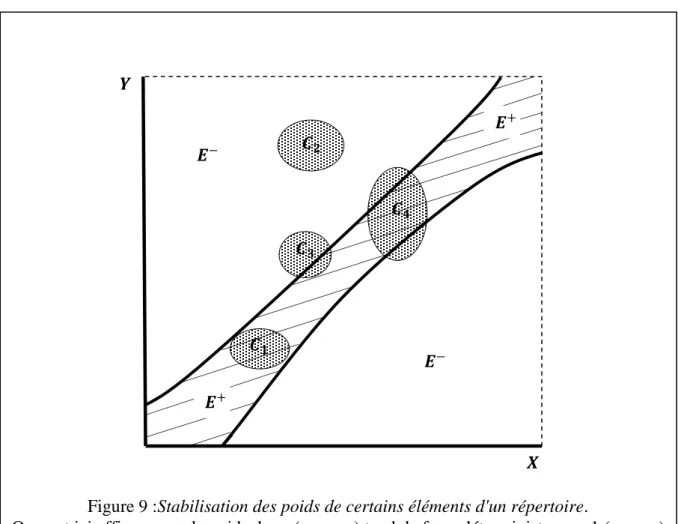
- Stabilisation des répertoires
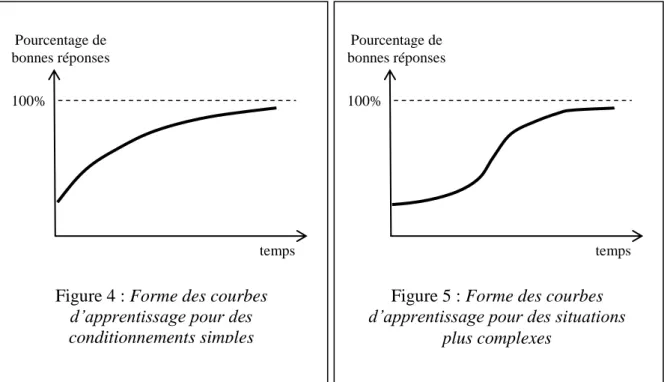
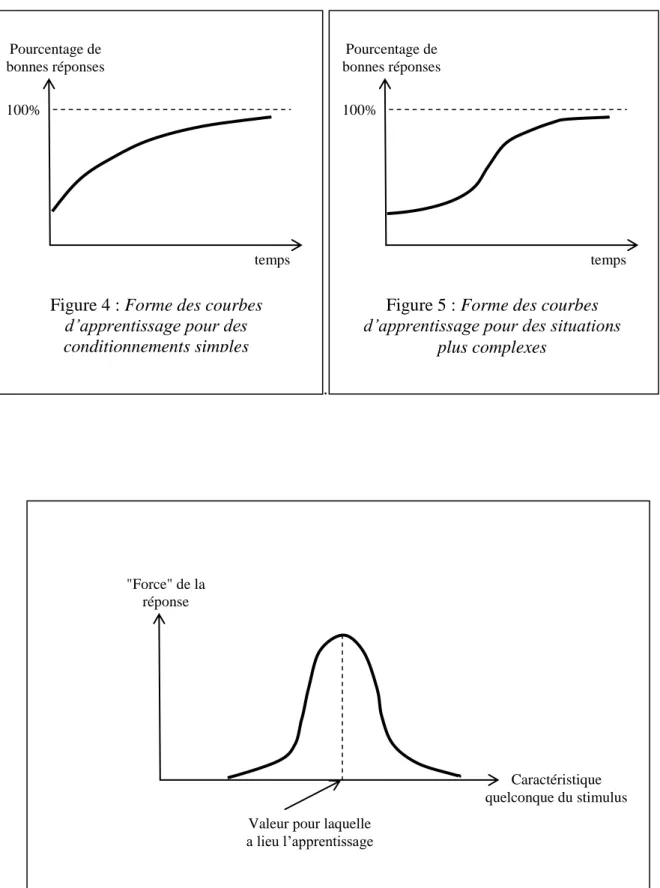
- Psychologie de l'apprentissage
- Neurophysiologie de l'apprentissage et "éléments" d'un répertoire
Nous présentons différents objets dans le champ de vision de la caméra et essayons d'apprendre à la personne aveugle à les reconnaître. En résumé, tous les exemples de plasticité perceptive que nous avons rapidement passés en revue conduisent aux conclusions suivantes.
STABILISATION D'UN RÉPERTOIRE
Présentation du modèle
Ainsi, pour une entrée donnée 𝑥 et des poids 𝑧 fixes, la probabilité de sortie guide va être Ce qui nous intéresse, c'est ce que l'on peut dire, pour 𝑡 assez grand, de la loi de probabilité de 𝑍𝑡.
Modèles d'apprentissage et processus de Markov
Ce qui nous intéressera, c'est ce que l'on peut dire, pour 𝑡 suffisamment, sur la loi de probabilité de 𝑍𝑡. Nous verrons d'abord sous quelles conditions on peut dire que la loi de probabilité de 𝑍𝑡 converge vers une loi de probabilité limite, indépendante de la loi de probabilité initiale des poids. En résumé, 𝑢𝑡 𝑒𝑡, 𝑧 représente la modification que la séquence d'événements 𝑒𝑡 produit en 𝑧 et 𝑝𝑡 𝑧, d𝑒𝑡 représente la probabilité de la séquence 0 de connaître la séquence.
Dynamique associée à un apprentissage à faible coefficient de renforcement
Comme nous l'avons dit, un stimulus distant peut être représenté par un point de type S2. L'action globale du groupe G peut être reconstituée au niveau de la "représentation interne" que nous avons définie au chapitre précédent.
FONCTION D'UN REPERTOIRE
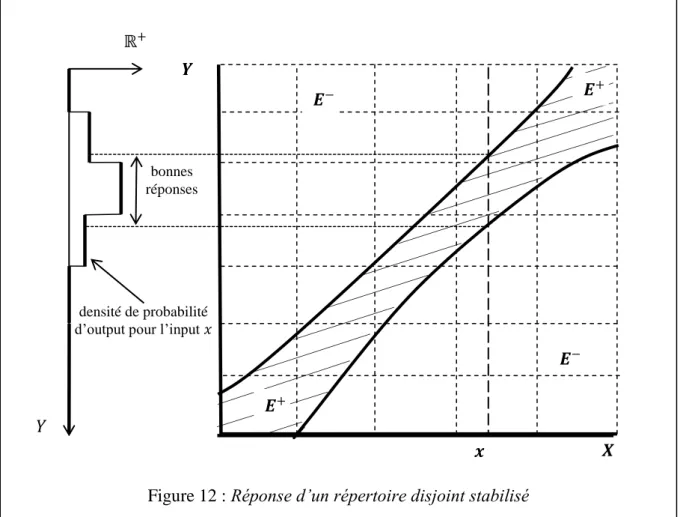
La relation input-output dans un répertoire stabilisé
Pour ce faire, nous devons considérer la forme de la probabilité de transition 𝑃 définie sur 𝑋 × 𝑌, qui donne la probabilité de sortie lorsque l'entrée est présentée au répertoire stabilisé. Par exemple, le centre de la rétine, la fovéa, est bien mieux représenté dans la zone 17 que la périphérie ; il y a beaucoup plus de cellules sensibles à cette partie du champ et leurs champs récepteurs y sont plus petits.
Caractéristiques de l'apprentissage pour un répertoire
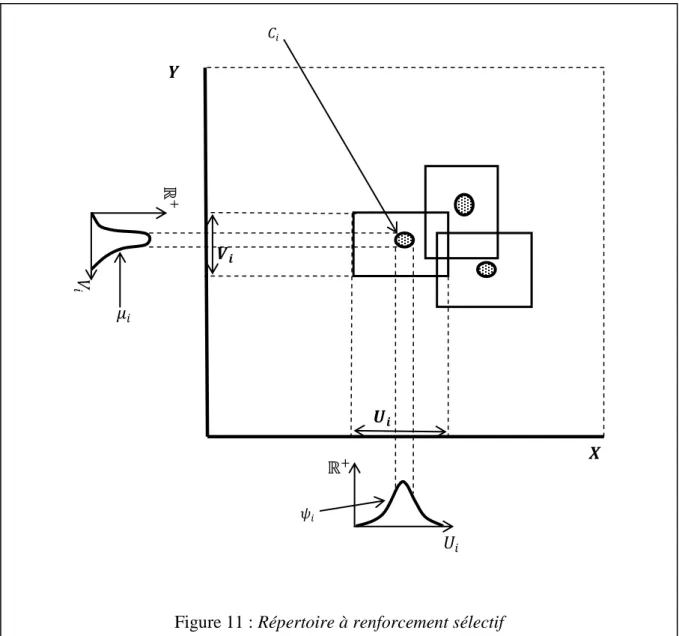
Ici aussi, le répertoire de renforcement sélectif est donc plus proche de la réalité biologique que le répertoire incohérent. On peut caractériser quantitativement ce réflexe, dont le siège est probablement dans le cervelet, en mesurant la vitesse angulaire de la tête 𝑢 (l'entrée) et la vitesse oculaire 𝑣 (la sortie).
Répertoires et "classeurs de forme"
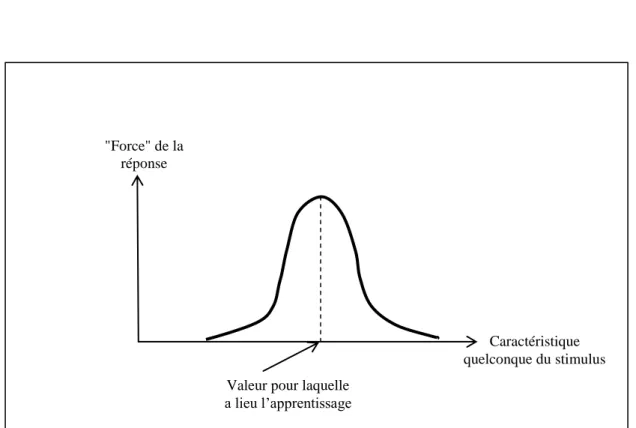
Au temps 𝑡, la machine est caractérisée par une certaine fonction 𝐷𝑡, que nous modifions comme suit : Appelons l'entrée 𝑥𝑡 présentée au temps 𝑡. Le choix de cette fonction est relativement arbitraire, mais on choisit généralement une fonction de la forme 𝜙(𝑥, 𝑦) = 𝑓 𝑑 𝑥, 𝑦 où 𝑑 est la distance sur 𝑋 et 𝑓 est une fonction décroissante (𝜙(𝑥, 𝑥𝑡) est donc une fonction "cloche" en 𝑥 s centrée au point 𝑥𝑡).
Répertoires et opérations "continues" sur les inputs
Ainsi, nous pouvons affirmer que nos annuaires peuvent effectuer n'importe quelle opération d'entrée continue avec une erreur de l'ordre de la capacité de discrimination du système. Bien entendu, une fois la stabilisation réalisée, les neurones correspondants se comporteront de la même manière que s'ils étaient câblés spécifiquement pour effectuer cette opération.
Non-fiabilité des neurones
Mais si 𝐸+ est un voisinage de 𝐺, disons dans le cas d'un répertoire sélectivement amplifié, tous les points de 𝐸 dont la distance à 𝐺 est inférieure ou égale à un nombre de l'ordre du diamètre de 𝐶𝑖 , la stabilisation donnera le résultat attendu, qui est que les réponses ont une probabilité très élevée d'être très proches de la bonne réponse (l'erreur est de l'ordre de la distance de discrimination du répertoire). Quant à la variabilité du second type, le fait que nos répertoires soient sous contrôle permanent d'amplification permettra de corriger toute erreur systématique qui s'y introduirait à la suite de la mort ou du dysfonctionnement de certains éléments.
Conclusion
Et puisque ce mouvement ne dépend que de la position du stimulus par rapport à la tête (ou plus précisément de la position « angulaire » : seule compte la direction du stimulus par rapport à la tête), on aura donc une variété de sortie 𝐿𝑇 de dimension obtenir 2, encoder l'emplacement (angulaire) dans un cadre lié à la tête. Le raisonnement que nous avons esquissé sur les mouvements de la tête peut être facilement généralisé à d'autres mouvements d'orientation.
COORDINATION SENSORI-MOTRICE
Mouvements d'orientation par rapport à des stimuli éloignés
- Mouvements des yeux
Bien entendu, le "calcul" de la vitesse et de la position du stimulus ne se fait dans ces répertoires qu'avec une certaine approximation. Si la personne veut fixer le stimulus, tout mouvement de la tête doit être corrigé par un mouvement des yeux dans la direction opposée.
Mouvements dépendant de la distance du stimulus
Le déplacement de l'image de l'objet sur une rétine dépendra de la distance à l'objet. En ce qui concerne les stimuli tactiles, au contraire, les informations transmises sont sans ambiguïté du point de vue de la localisation égocentrique.
Hiérarchie des répertoires sensori-moteurs
Au niveau des systèmes d'annuaires, le rôle de la structure a priori du système est encore plus clair. Nous avons vu le rôle permanent de ce type de répertoire dans la coordination sensori-motrice (chapitre V).
INVARIANCE PERCEPTIVE ET GEOMETRIE DE LA PERCEPTION
Perception et invariance perceptive
Notre hypothèse consistera à supposer que ces changements sont le résultat de la stabilisation des répertoires impliqués dans l'exécution de ces comportements et qu'ils sont par conséquent le résultat d'apprentissages.Cette hypothèse est conforme à ce que nous avons vu sur l'adaptabilité des le système perceptif et les principes généraux de l'activité neuronale. Selon que l'on privilégiera, dans le système des répertoires, la discrimination opérée par les répertoires d'entrée, de sortie ou par l'organisation du système lui-même, il sera placé dans l'un des trois courants que nous avons cités.
Invariance locale par rapport aux mouvements du corps
Supposons maintenant qu'une partie du corps (par exemple, le bassin) est fixe et reste fixe sous l'influence de 𝑀, et que tous les 𝑚 ∈ 𝑀 spécifient l'état et la position de l'organe sensoriel en question. " source d'information pour le stimulus, cela signifie que 𝑆 et 𝑉 ont la même dimension et qu'il existe une application injective 𝑓𝑚 de 𝑆 à 𝑉, pour tout 𝑚 donné. , par :𝑓 𝑠, 𝑚 = 𝑓𝑚 𝑠 , 𝑚. Nous supposons que cette application 𝑓 est différentiable, il s'agit donc d'un difféomorphisme de 𝑆 × 𝑀 vers un ensemble ouvert 𝑈 de 𝑉 × 𝑀.
Invariance globale par rapport aux mouvements du corps
Dans ce cas, ce n'est pas tout le groupe 𝐺 qui agit sur 𝑆 × 𝑀 : il faut se limiter à un ouvert de 𝐺 contenant l'identité (groupe local)1 définie pour chacun (𝑠, 𝑚) des éléments 𝑔 ∈ 𝐺 tel que 𝑓𝑚 𝑠 , 𝑔𝑚 = 𝑡𝑔∘ 𝑓(𝑠, 𝑚) appartient à 𝑈. Il nous semble donc qu'il existe une voie de recherche intéressante au sein de la reconnaissance de formes.
Mouvements des objets
Par exemple, les informations sur la position et la vitesse d'un point objet ne sont pas suffisantes pour déterminer si l'objet est animé d'un mouvement de translation ou d'une rotation sur lui-même. Cette dernière remarque est particulièrement importante : elle démontre en fait que la reconnaissance du mouvement ne se fait pas à partir de "l'intégration" temporelle de la vitesse à chaque instant, mais directement à partir des données du "champ de vitesse".
Invariance d'un objet par rapport à son propre mouvement
Ces voies forment une partition de la variété, où chaque voie est caractérisée par la valeur d'un paramètre invariant sous l'influence du groupe. Ce point de vue s'oppose à d'autres théories qui ont également tenté d'expliquer l'immuabilité de la reconnaissance de forme par rapport aux déplacements d'image sur la base de données neurophysiologiques récentes.
Mesures perceptives
La corrélation entre distance perçue et taille perçue serait alors facile à expliquer : toute erreur d'appréciation de la distance proviendrait d'une mauvaise représentation des deux extrémités du segment dans le référentiel égocentrique et conduirait automatiquement à une erreur d'appréciation de la longueur dans le répertoire correspondant. qui utilise cette représentation en entrée. Ce raisonnement que nous venons d'envisager concernant la taille pourrait bien entendu être généralisé à toute caractéristique globale des objets.
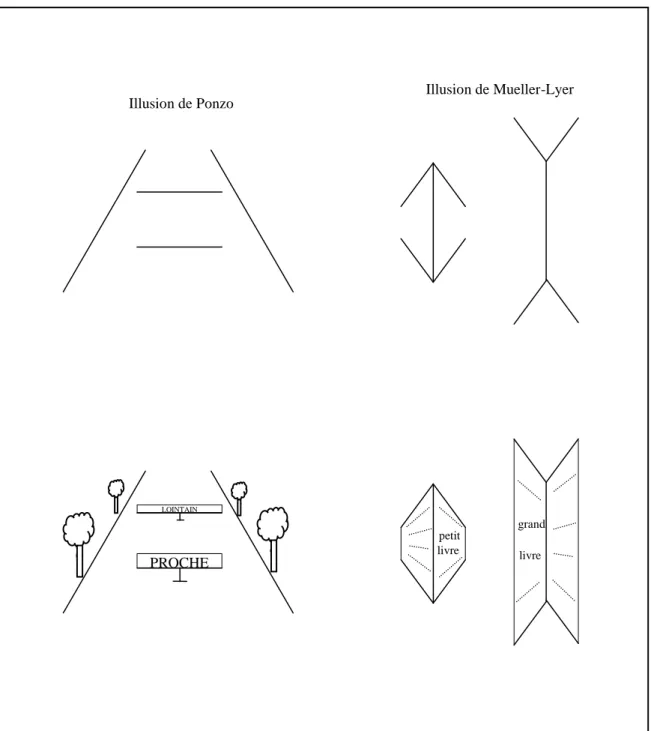
La géométrie de l'espace perceptif
Cela réfute l'idée que l'organisation perceptive met en évidence un véritable tableau dû aux illusions visuelles. Nous retrouvons le rôle fondamental de l'expérience dans la structuration de la perception.
Aspects mathématiques
De telles machines doivent en fait classer les formes selon les mêmes critères que nous pour être utiles. Il est donc essentiel de faire le lien entre l'apprentissage par renforcement tel que nous le connaissons dans le système nerveux et les techniques utilisées en intelligence artificielle.
Aspects psychologiques et épistémologiques
Il nous semble donc que l'organisation fonctionnelle dans ce modèle est le résultat d'une interaction entre le système et son environnement, dans laquelle la structure « innée » et l'expérience jouent toutes deux un rôle essentiel : la structure initiale détermine le type d'invariants que le système est susceptible de détecter et d'apprendre détermine la forme exacte de ces invariants, s'ils existent naturellement dans l'environnement. Si l'on ajoute à cela le rôle essentiel que jouent les mouvements actifs de l'organisme dans le modèle de détermination des invariants perceptifs, notamment au niveau des étapes du développement progressif du système (stabilisation des répertoires sériels), on reconnaîtra Ce modèle ne relève pas d'une conception empiriste mais s'inscrit plutôt dans le cadre de la théorie de l'"épistémologie génétique" de Piaget qui met l'accent sur cette dualité entre structure interne et action de l'environnement.
Aspects neurophysiologiques
Epstein (Red.): Progress inPsychobiol.-and Physiol. 1972), Eye Movements, in Handbook of Sensory Physiology, VII/4: Visuele Psigofisika. Epstein (Red.): Stability and Constancy in VisualPerception. 1978). The Retinotopic Organization of Lateral Suprasylvian visual Areasin the Cat,J.