Jean-Michel Geneste m'a accueilli au sein de l'équipe scientifique de la Grotte Chauvet qu'il dirige et nous a généreusement soutenu dans l'étude génétique que nous avons menée sur les échantillons Chauvet. J'entendrai toujours son accent chantant me décrire les peintures et gravures de la Grotte.
PTB :Phénacylthiazolium
LA FAUNE DU PLEISTOCENE
- Données paléontologiques sur l’ours des cavernes
- Caractéristiques morphologiques de l’ours des cavernes
- Position systématique de l’ours des cavernes
- Paléo-éthologie de l’ours des cavernes
- Données isotopiques définissant le régime alimentaire de l’ours des cavernes
- Données paléontologiques sur l’hyène des cavernes
- Caractéristiques morphologiques de l’hyène des cavernes
- Origine évolutive de l’hyène des cavernes
- Alimentation
- Paléo-éthologie de l’hyène des cavernes
Certains crânes d'ours des cavernes ne possèdent pas cette caractéristique, mais de telles exceptions sont rares (Kurtén, 1976). Cependant, deux études récentes remettent en question le caractère strictement herbivore du régime alimentaire de l'ours des cavernes.

LA PALEOGENETIQUE
- La naissance tumultueuse d’une discipline
- Les limites de la paléogénétique
- La dégradation de l’ADN
- L’établissement de lésions bloquantes
- Les dommages de l’ADN
- La présence de molécules inhibitrices
- Les risques de contamination
- Les mesures de précautions permettant d’éviter les contaminations
- Les méthodes de l’ADN ancien
- Les matériaux utilisés
- Les séquences d’ADN ciblées
- Les apports de la paléogénétique
- Identification de vestiges
- Reconstitution de phylogénies
- Étude en génétique des populations
La contamination de l'ADN humain a également été démontrée dans les solutions d'amorces (Deguilloux et al., 2011). Les échantillons conservés dans le pergélisol peuvent contenir de l'ADN qui peut être amplifié pendant plusieurs centaines de milliers d'années (Willerslev et al., 2004a).

LES GROTTES D’OU PROVIENNENT LES ECHANTILLONS ETUDIES. ETUDIES
- Présentation générale des grottes des Gorges de l’Ardèche
- La Grotte Chauvet-Pont d’Arc
- Découverte de la Grotte Chauvet-Pont d’Arc
- La protection d’un patrimoine unique au monde
- Les représentations artistiques de la Grotte Chauvet-Pont d’Arc
- Les occupations humaines de la Grotte Chauvet-Pont d’Arc
- Les occupations animales
- La Grotte des Deux-Ouvertures
- Découverte de la Grotte des Deux-Ouvertures
- Les occupations humaines dans la Grotte des Deux-Ouvertures
- Les occupations ursines dans la Grotte des Deux-Ouvertures
- La Grotte de Coumère
La grotte Chauvet-Pont d'Arc est la grotte ornée la plus importante de la région. Parmi les témoignages de la présence d'animaux dans la grotte Chauvet-Pont d'Arc, les plus importants concernent l'ours des cavernes. Toutes les chambres de la grotte Chauvet-Pont d'Arc ne stockent pas les échantillons de la même manière.
Les conditions de stockage de la grotte Chauvet-Pont d'Arc limitent le prélèvement d'échantillons osseux, ainsi que leur manipulation. L'ubiquité de l'ours des cavernes permet de le définir comme une grotte à ours, au même titre que la grotte Chauvet-Pont d'Arc (Figure 25).

SEQUENÇAGE DU GENOME MITOCHONDRIAL DE L’OURS DES CAVERNES L’OURS DES CAVERNES
- Introduction
- Résultats : Article 1
Un arbre phylogénétique construit avec ce fragment de la région de contrôle place l'ours des cavernes en dehors de la variabilité de l'ours brun, soutenant une divergence précoce des lignées arctoïdes et spéléoïdes. La région de contrôle des ours bruns et polaires présente une délétion de 13 pb, absente chez les ours noirs américains et tibétains, ainsi que chez l'ours des cavernes. Pour obtenir une phylogénie robuste de l’ours des cavernes, nous avons décidé de séquencer le génome complet de cette espèce.
Un génome mitochondrial d'ours brun de la lignée d'Europe occidentale a également été séquencé pour clarifier la position phylogénétique de l'ours des cavernes. Ces données, ajoutées aux génomes mitochondriaux complets publiés (Delisle & Strobeck, 2002 ; Yu et al., 2007), ont permis de déterminer la position phylogénétique exacte de l'ours des cavernes, ainsi que de dater les différents événements de spéciation chez l'ursine. lignée.
Deciphering the complete mitochondrial genome and phylogeny of the extinct cave bear in the
The Chauvet-Pont d'Arc cave is expected to contain cave bear rather than brown bear remains (16, 17). The length of the mitochondrial genome of the cave bear (16,810 bp) is in the range of that reported for extant bear genomes, which varies between 16,723 (Ursus maritimus) and 17,044 bp (Ursus thibetanus formosanus). This particular domain of the control region could not be recovered by a single PCR from the Chauvet-Pont d'Arc cave bear sample.
Second, the mitochondrial genome of the cave bear allows better investigation of the archaeological patterns attributed to this species. DNA sequence authentication. To ensure the authenticity of the cave bear mitochondrial sequence, we followed previous recommendations for work done on ancient DNA (18, 19).

Supporting Information
Comparaison avec les autres séquences d’ours anciens
Un autre génome mitochondrial complet d'ours des cavernes a été déposé dans GenBank quelques mois après celui de la Grotte Chauvet-Pont d'Arc et publié la même année. La séquence obtenue de l'ours des cavernes de Gamssulzen est très proche de celle de Chauvet-Pont d'Arc. L'ours des cavernes et l'ours à face courte ne sont pas les seuls Ursides étudiés dans l'ADN ancien.
En fait, une mâchoire d'ours polaire préservée dans le pergélisol a également fourni un génome mitochondrial complet (Lindqvist et al., 2010). Cet arbre phylogénétique (Figure 26) montre que les deux séquences d'ours des cavernes sont proches et forment un groupe monophylétique.

Conclusion
ANALYSE GÉNÉTIQUE DES POPULATIONS D'OURS DANS LES ÉTUI DE LA VALLE DE L'ARDÈCHE EN VALLÉE DE L'ARDÈCHE.
ANALYSE GENETIQUE DES POPULATIONS D’OURS DES CAVERNES DE LA VALLEE DE L’ARDECHE CAVERNES DE LA VALLEE DE L’ARDECHE
- Introduction
- Résultats : Article 2
Cependant, les plus anciennes traces d'habitation humaine remontent à cette date, ce qui amène les auteurs à supposer une intervention humaine dans la disparition de la première génération d'ours des cavernes (Hofreiter et al., 2007). Plus sensible que l'ours brun aux climats froids, il aurait pu avoir du mal à tolérer d'importantes variations de température à la fin du Pléistocène (D'Errico et al., 2001). Une richesse de données sur la génétique de l’ours des cavernes permet d’estimer la taille réelle de la population au fil du temps et de la comparer à l’ours brun encore présent (Stiller et al., 2010).
Cette analyse semble exonérer l’homme moderne du déclin de la diversité génétique des ours des cavernes. Les ours des cavernes de la Grotte Chauvet ne peuvent pas être étudiés par les paléontologues car la manipulation des échantillons est limitée.
Introduction
The cave bear has long been considered to have survived until the last millennia of the Pleistocene (Kurtén, 1976), an idea questioned by Pacher and Stuart (Pacher and Stuart, 2009). Consequently, the causes responsible for cave bear extinction are unclear and require further studies. In the present study, we focused on two sites from southeastern France (Fig. 1) that display both a large number of cave bear remains and evidence of Paleolithic human intrusion.
Preservation of ancient DNA in this cave was demonstrated through the recovery of a complete cave bear mitochondrial genome from a 32,000-year-old sample (Bon et al., 2008). In 2007, excavations conducted near these images yielded charcoal fragments as well as abundant cave bear remains (Philippe et al., 2009).
Materials and Methods
They were performed using primers (Table 1) that have been shown to be effective for amplifying a 112-bp fragment of the cave bear mitochondrial 16S gene. To ensure the authenticity of cave bear mitochondrial sequences, we followed previous recommendations for work done on ancient DNA (Pääbo et al., 2004; Willerslev and Cooper, 2005). Phylogenetic analysis was performed on 253 nucleotides of the mitochondrial control region, located between positions 16,364 and 16,623 of the published complete mitochondrial genome US18 (Bon et al., 2008).
To estimate the reliability of the phylogenetic inferences, we used the bootstrap method and performed 1,000 replications. For each site, we calculated the maximum likelihood estimate of the parameter θ ≡ Nµ (where N is the effective population size and µ is the mutation rate) using Ewens' (Ewens, 1972).
Results
The Chauvet-Pont d'Arc Cave bear samples yield radiocarbon dates ranging between 29,050 and 37,300 years BP, but most of them were younger than 32,000 years BP (Table 2). In the Deux-Ouvertures cave we obtained radiocarbon dates that partially overlap those found in Chauvet-Pont d'Arc. However, two samples (US299 and US735) were dated to 28,000 years BP, indicating slightly later occupation in the Deux-Ouvertures than in the Chauvet-Pont d'Arc cave.
This second experimental series demonstrated cave bear DNA in three additional Chauvet-Pont d’Arc samples (Table 2). In contrast, the dominant haplotype is widely distributed and is present in five Chauvet-Pont d'Arc samples, in the Deux-Ouvertures samples, as well as in samples obtained from Arcy (France), Eiros (Spain) and Scladin (Belgium). ).
Discussion
The detection of only two mitochondrial haplotypes suggests that the genetic diversity of cave bears was low in the Ardèche several years ago. The low genetic diversity and effective population size of these late cave bears are consistent with genetic decline that began approximately 50,000 years ago (Stiller et al., 2010). The current work, together with previous analyzes conducted by Bocherens et al. 2006), makes available a dataset of 20 radiocarbon dates for Chauvet-Pont d'Arc cave bears.
Comparison of the cave bear data with this time span (Fig. 6) shows that: 1) animal remains predate the earliest anthropogenic evidence. We further suggest that cave bears had already disappeared from the Ardeche during the Gravettian, which observation is supported by our data for the Deux-Ouvertures Cave.
Conclusions
- Résultats complémentaires sur la Grotte des Deux- Ouvertures
- Discussion
- Conservation des échantillons
- Comparaison des populations d’ours des Grottes Chauvet-Pont d’Arc et des Deux-Ouvertures d’Arc et des Deux-Ouvertures
- Apports sur les causes de la disparition des ours des cavernes en Ardèche et sur la connaissance du site en Ardèche et sur la connaissance du site
Outre les cinq humérus présentés dans l’article, onze autres échantillons de la grotte des Deux-Ouvertures ont été examinés selon le même protocole. Tous ces échantillons représentent le même haplotype, identique à l'haplotype majoritaire chez les ours des cavernes de la grotte Chauvet-Pont d'Arc. Une étude ostéologique et ostéométrique des ours des cavernes de la Grotte des Deux-Ouvertures est prévue pour pallier le manque d'informations sur les spécimens de la Grotte Chauvet-Pont d'Arc (Philippe et al., 2009).
Ce programme de datation des ours des cavernes de la grotte Chauvet-Pont d'Arc avait pour dater auparavant le moment à partir duquel les grands animaux ne pouvaient plus entrer dans la grotte Chauvet-Pont d'Arc. En montrant que les ours des cavernes ont disparu de la région il y a 28 000 ans, nos données permettent de déterminer l'âge minimum des représentations d'ours dans la grotte Chauvet-Pont d'Arc.

INTRODUCTION
Les échantillons de coprolite fournissent également des informations sur le régime alimentaire des animaux qui l'ont produit. Chez les coprolites paresseux, le gène rbcL a été amplifié et séquencé, indiquant la présence d'au moins sept plantes différentes (Poinar et al., 1998). Un fragment du gène 18S et une partie du gène du cytochrome b mitochondrial ont été séquencés, mais n'ont pas permis d'identifier avec certitude à l'espèce à laquelle appartiennent ces échantillons (Loreille et al., 2001b).
Les coprolites humains trouvés sur des sites précolombiens en Amérique du Sud ont révélé des séquences des gènes 5S et SL1 d'Enterobius vermicularis, un ver parasite du tube digestif humain, et ont éclairé l'origine du gène SL1. évolué depuis (Iñiguez et al., 2006). Ainsi, pour identifier les plantes trouvées dans un coprolite paresseux, l'amplification d'un fragment de 183 pb est nécessaire (Poinar et al., 1998).
ETUDE DE COPROLITHES D’HYENE DES CAVERNES
- Extraction d’ADN à partir de coprolithes
- Rôle de l’EDTA
- Rôle du N-PTB
- Conclusion
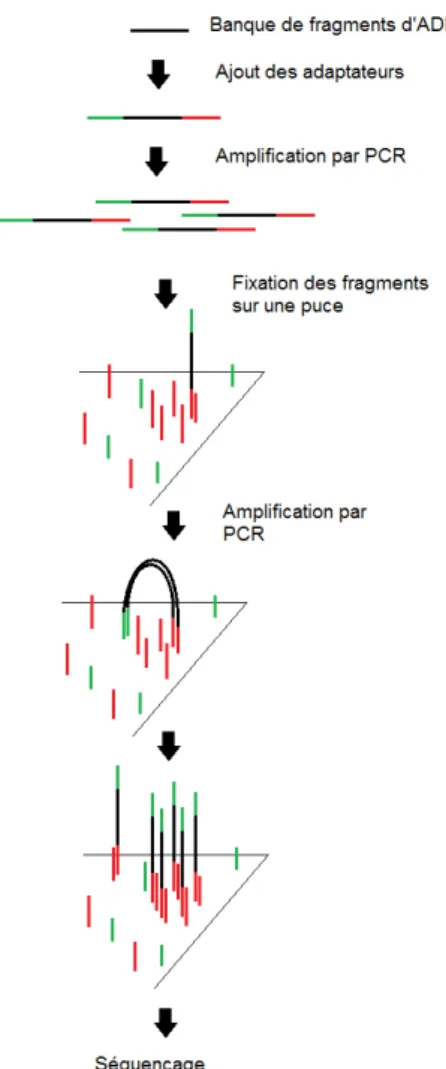
- Analyse métagénomique
- Choix des échantillons
- Séquençage massif des deux banques d’extraits de CC8 et CC9
- Résultats de l’analyse métagénomique : Article 3
- Résultats et discussion complémentaires
The largest number of hits (6.1% of readings) corresponded to the domestic cat (Felis catus). The tree topology was strongly supported and unambiguously establishes the close evolutionary relationship between the spotted hyena and the cave. The extensive coverage of the cave hyena mitochondrial genome indicated that it was important to quantify the number of nuclear DNA sequences for this species in the coprolite samples.
The extensive coverage of the cave hyena mitochondrial genome ( Fig. 3A ) allowed the calculation of a sequence accuracy of >99% ( Fig. S2 ). To check for some of the differences between cave and spotted hyena mitochondrial genome sequences, we performed PCR analysis of coprolite extracts. To independently assess the presence of cave hyena nuclear DNA fragments in coprolites, we performed PCR analysis of the CC8 extract.
Only positions of the alignment for which sequence information (total length, 2221 nucleotides) was available for the cave hyena were used for phylogenetic analysis.









![Reação de Cicloadição [3+2] 1,3-Dipolar. Uma Metodologia Eficiente na Síntese de Sistemas 3,5-Isoxazolínicos Líquido-cristalinos.](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)