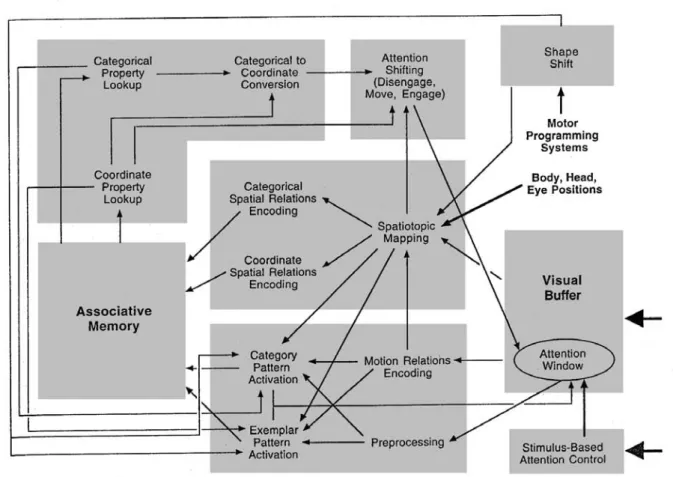
Il est généralement admis que cette voie prend cette information de la voie parvocellulaire (Desimone et al. 1985). Le contenu de la fenêtre attentionnelle est alors repris simultanément par les systèmes ventral et dorsal. Le système s'engage alors dans un autre cycle de reconnaissance, pour lequel la représentation de la tête d'un renard sera privilégiée, et l'attention portée sur cette partie de l'image.
Le modèle est basé sur l'idée d'un grand nombre de vues mémorisées, notamment pour la reconnaissance au niveau des classes suivant un mécanisme ascendant. Après avoir postulé que l'attention doit « formater » la représentation de l'objet avant toute reconnaissance, les auteurs extraient les caractéristiques de la fenêtre attentionnelle. Le contenu de la fenêtre (en haute résolution) est envoyé vers un module de reconnaissance (mémoire associative).
Ce format a l'avantage de réduire le problème de reconnaissance combinatoire : le nombre d'entrées possibles pour le nombre de vues stockées. Par conséquent, ces mécanismes peuvent expliquer de manière simple la propriété principale de l’attention sélective spatiale.
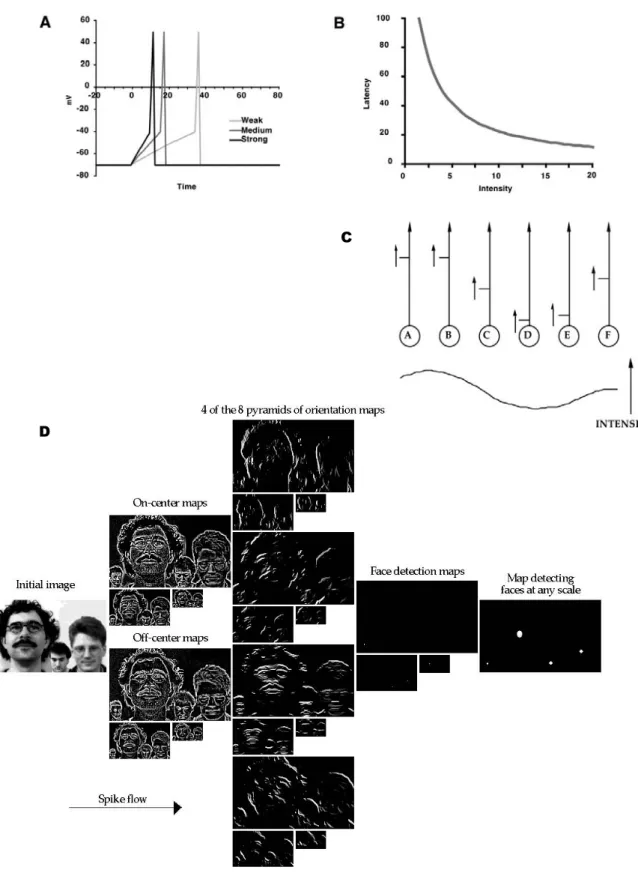
VITESSE DU TRAITEMENT VISUEL
Cela comprend la prise de photos, une tâche à effectuer par les sujets et une méthode d'analyse. a) choisir la tâche. Encore une fois, 150 ms indique le délai après lequel le traitement visuel est suffisant pour établir la présence ou l'absence d'une catégorie aussi diversifiée qu'un animal. En revanche, le délai SN est plus rapide pour la forme globale (pointer à 230 ms pour la configuration globale, à 300 ms pour la forme locale).
Cependant, il apparaît que la topologie de l'activité différentielle « animale » ressemble à celle du SN, notamment dans l'étude d'Anllo-Vento et al. Ces caractéristiques ne semblent pas correspondre à l'activité différentielle des « animaux », bien que l'opération d'attention sur les caractéristiques visuelles de la catégorie animale soit très probable. e) saillance des catégories visuelles connues (P150) Les P150 combinent des ondes provoquées par deux ensembles de stimuli : les visages et les mots. De même, les générateurs que nous avons mis en avant semblent moins datés que les générateurs P150.
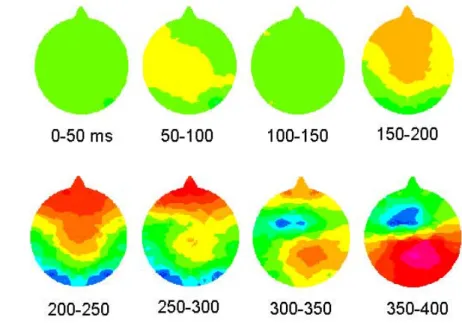
VARIATIONS AUTOUR DE LA CATEGORISATION VISUELLE RAPIDE
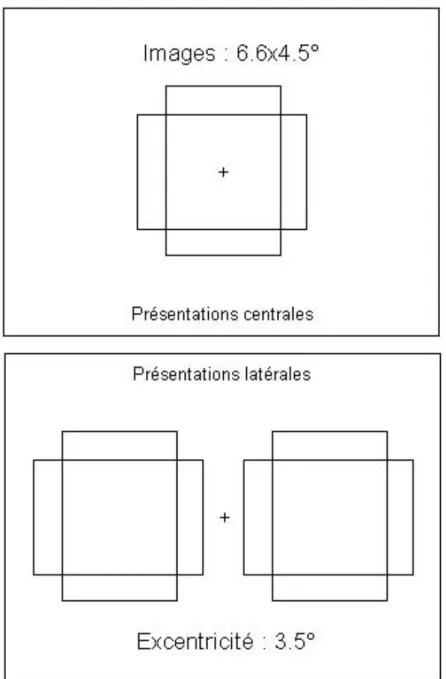
Le numéro précédent montrait une attention accrue portée aux cibles et aux distractions de tâches. Nous montrons que la reconnaissance des catégories d'animaux n'est que légèrement affectée (en termes de taux de réussite et de temps de réaction) par des présentations décentrées de 3,5°, et que les bords latéraux des photographies s'étendent jusqu'à près de 7°. L’observation de la couleur n’est donc pas une stratégie efficace pour accomplir la tâche : il n’y a pas de couleur.
La tâche de catégorisation visuelle recrute probablement de nombreuses étapes de la voie ventrale, impliquées dans la reconnaissance des objets. Les potentiels évoqués par les cibles diffèrent des potentiels évoqués par les attracteurs à partir de 150 ms, indépendamment de la couleur des stimuli (tâches mixtes). Cet effet de tâche est également similaire aux différences cible-distracteur dans la tâche.
Il est remarquable de constater que la présence de la couleur intervient de la même manière dans les scènes, qu'elle soit cible ou non. Cette activité présente des topographies similaires dans les deux tâches, quelle que soit la nature de la cible (couleur, N&B). Cela semble similaire à l'activité observée dans le cas de la détection de la catégorie animale lors d'expériences précédentes. a) codage visuel et adéquation aux tâches.
La différence d'activité à partir de 150 ms est clairement liée au statut de cible et de distraction de la tâche. Cet effet pourrait-il être dû à un mécanisme d’amorçage lié à la répétition de la catégorie animale au cours de l’expérience ? On peut penser que l'activité liée à la reconnaissance visuelle de la catégorie est fortement renforcée lorsque cette catégorie est pertinente à la tâche.
Durant l'événement 'Carrés contre. Dans le cadre de la tâche des cercles, les sujets devaient relâcher un bouton de la souris le plus rapidement possible dès que l'image affichée contenait des carrés. Une différence entre les potentiels évoqués par les deux types de scènes est donc beaucoup plus probable dans la tâche « animaux » que dans la tâche « formes simples ». À l’inverse, les scènes de la tâche animal versus non-animal sont plus complexes et beaucoup moins redondantes, et peuvent contenir des informations ambiguës sur la présence d’un animal dans l’image.
ETUDE EN IRMf
Outre la possibilité de mélanger des types de stimuli dans une même séquence de stimulation (plans expérimentaux mixtes), grâce à cette méthodologie Savoy et al. 1995 a détecté des changements de signal dans les champs visuels dus à des stimulations d'une durée de seulement 34 ms. On voit donc que dans ce cas des plus défavorables, l'ordre des stimuli permet d'obtenir des indices sur l'implication de la zone dans le traitement de chaque type.
Dans le cas plus favorable d'une zone impliquée dans le traitement d'un seul type de stimulus, le niveau du signal permettra facilement l'éclairage, toujours sous réserve d'une variance suffisante du niveau de chevauchement (Figure 41). Pour cinq stimuli consécutifs, le nombre d'événements de type TTDTD (deux cibles, puis un attracteur, suivis d'une cible et d'un distraction) est de 32. Il nous semble impossible qu'un sujet humain puisse deviner la loi d'une telle séquence1.
Lors d'un run, les données sont collectées régulièrement toutes les 2,5 secondes, soit 1,5 et 4 secondes après la présentation de chaque stimulus. Les fonctions c(t) et d(t) ont été définies comme la convolution d'une fonction de Poisson P(λ) (représentant la réponse hémodynamique à une seule stimulation) à la séquence de stimulation modélisée par une série de distributions Dirac δ représentant un stimulus dans une certaine heure. Le choix d'un Poissonien est courant dans ce contexte ; λ choisi arbitrairement pour que la réponse hémodynamique atteigne son maximum 6 s après le stimulus (λ = 6) initialement.
L'inconvénient réside essentiellement dans les longs délais d'attente des sujets, les intervalles entre stimuli étant de l'ordre d'une quinzaine de secondes. Selon Kosslyn, la zone 19 (avec une très grande surface) est impliquée dans des processus associés de mémoire (p. 255 Image et Cerveau), implication observée dans sa partie la plus dorsale (Kosslyn et al. Cet effet s'explique généralement par une série processus qui se termine une fois qu’un objectif est trouvé et qui est par ailleurs atteint.
On peut noter qu'un tel mécanisme pourrait être à l'origine des résultats d'études sur l'amorçage qui ont montré paradoxalement qu'une meilleure reconnaissance et des temps de réaction plus courts aux stimuli amorcés étaient associés à des diminutions de

BASES CEREBRALES DE LA CATEGORISATION
Nous avons proposé que ce phénomène soit lié à une activité de type prise de décision, indépendamment du fait qu'il s'agisse d'un objectif de tâche ou d'un distraction. Cette différence de 150 ms est très proche de la négativité de sélection (SN) observée dans les essais de potentiels évoqués d'attention sélective sur des caractéristiques visuelles simples et des couleurs. Une étude IRMf révèle que les gyri médiaux occipitaux, cingulaires et fusiformes sont des sites d'activité différentielle au cours de la tâche.
Cette étude IRMf nous a permis de proposer que ces zones visuelles réduisent leur activité lors de la détection de cibles, via un mécanisme d'inhibition qui supprime l'activité des neurones non sélectionnés. Nous suggérons donc que l’effet observé avec l’IRMf au cours de la tâche reflète un tel mécanisme. Pour l'activité précoce visible entre 110 et 140 ms, cet effet ne peut être démontré de la même manière.
Animal vs non-animal explicite » de toutes les expériences, suggérant que ce sont les mêmes aires corticales recrutées dans le cas implicite et lors de la tâche animale (la première est identique à l'activité « précoce », la seconde à « l'activité différentielle » à 150 ms'). Cette propriété est tout à fait cohérente avec notre hypothèse de suppression d'activité au cours de la tâche : une zone plus activée par les scènes animales que par d'autres scènes lors de la reconnaissance implicite va fortement réduire son activité lorsque les animaux sont cibles de la tâche, par inhibition de tout ce qui ne l'est pas. -neurones sélectionnés dans la même zone. Cet effet est identique à la « négativité de sélection », qui fait référence au signe de la différence.
Dans un exemple explicite, lors d'une tâche de catégorisation animale, l'attention préparatoire portée à cette catégorie a pour effet d'augmenter la sensibilité des neurones à leurs entrées dendritiques dans cette zone. Lorsqu'une scène cible est présentée, l'activation de quelques neurones très sélectifs pour les formes de l'animal actuel provoque également une forte inhibition des autres neurones de la même zone. Nous proposons donc que ce mécanisme inhibiteur explique le changement de signe de la différence « implicite vs explicite animal vs non-animal » observée dans les potentiels évoqués.
Lors de la tâche de catégorisation explicite utilisant le même type de stimuli, on devrait observer des activations localisées dans les mêmes zones au moyen des mêmes comparaisons, mais cette fois le signe de leur différence devrait être opposé.

COMMUNICATIONS
BIBLIOGRAPHIE
Single neurons with both shape/color differential responses and saccade-related responses in the nonretinotopic pulvinar of the behaving macaque monkey. The functional organization of human extrastriate cortex: a PET-rCBF study of selective attention to faces and locations. Event-related potentials to conjunctions of spatial frequency and orientation as a function of stimulus parameters and response requirements.
The time course of brain activations during response inhibition: evidence from event-related potentials in a go/no go task. Bridging the gap between monkey neurophysiology and human perception: an ambiguity resolution theory of visual selective attention. Combined event-related fMRI and EEG evidence for temporal-parietal cortex activation during target detection.
Common and differential effects of attentive fixation on the excitability of parietal and prestitial (V4) cortical visual neurons in the macaque monkey.