Le développement de futurs médicaments anti-Alzheimer et d’antidotes contre les intoxications aux organophosphorés (OP) bénéficiera d’une compréhension approfondie de la dynamique structurelle de l’AChE. Les travaux menés dans cette thèse s'inscrivent dans le domaine de la cristallographie cinétique.
LA DYNAMIQUE STRUCTURALE DES PROTÉINES
- Le paysage conformationnel
- Mouvements, échelles de temps et méthodes associées
- Relations dynamico-structurales entre une protéine et son solvant
- Transition vitreuse du solvant et transition dynamique d’une protéine
Cependant, en principe, elle permet uniquement d’étudier la structure de la protéine au repos. Certains le considèrent comme faisant partie intégrante de la structure des protéines [Ball 2007].

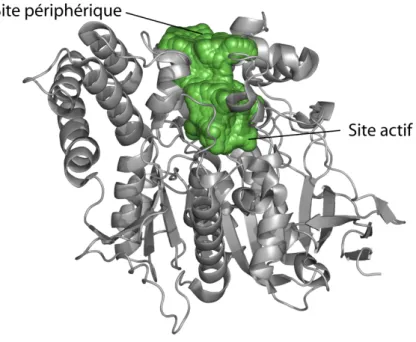
L’ACÉTYLCHOLINESTÉRASE (AChE)
- Découverte
- Rôle classique
- Une enzyme très rapide
- Structure tridimensionnelle
- Le site actif
- La gorge aromatique
- Le site périphérique
- Fonctionnement : mécanisme catalytique et paradoxe
- Rôles non-classiques
Depuis les années 1950, la quête pour comprendre la fonction de l’AChE a donné lieu à des efforts considérables. Le sous-site anionique est également impliqué dans la liaison aux inhibiteurs du site actif AChE.

L’AChE, CIBLE D’UN VASTE RÉPERTOIRE DE LIGANDS
Les organophosphorés
- Origine et utilisation des organophosphorés
- Mécanisme d’inhibition de l’AChE par les OP
- Réactivation de l’AChE inhibée par un OP
Deuxièmement, l'un des substituants OP est clivé (cela correspond généralement à une désalkylation). L'encombrement stérique du site actif AChE joue également un rôle dans l'efficacité des réactivateurs.

Toxines
- La fasciculine 2
- L’aflatoxine
C'est le cas de la phosphotriesterase bactérienne (PTE) [Dumas 1990] et de la paraoxonase humaine [Rochu 2007] (ces enzymes sont optimisées par évolution dirigée). L'aflatoxine est une toxine produite par des champignons de la famille Aspergillus (l'espèce Aspergillus flavus a donné son nom à la toxine : A-fla).

Les médicaments anti-Alzheimer
En effet, AB1 est également un inhibiteur PAS de l'AChE, même si ce rôle n'est pas important dans le phénomène de toxicité. Au cours de cette thèse, nous avons cherché à élucider la structure de l'AChE en complexe avec AB1 (voir section 6.3).

OBJECTIFS DE LA THÈSE
Nous avons donc participé à deux projets de cristallographie et initié la mise en œuvre d'une méthode permettant de caractériser la dynamique lente de l'AChE. A cette occasion, nous avons obtenu les structures de TcAChE en complexe avec AB1 (section 6.3).
MÉTHODOLOGIE
La cristallographie des protéines
- Introduction
- Les principes de la cristallographie des protéines
- La cristallogenèse
- Le montage des cristaux sur le diffractomètre
- La collecte de données
- Traitement des données cristallographiques
- Résolution d’une structure
- Affinement de la structure
- La cristallographie cinétique
Une expression permettant le calcul de l'onde réfractée par le cristal à partir de la distribution de densité électronique de la protéine peut être facilement déterminée. Le but de l'expérience cristallographique sera alors d'inverser cette relation et de calculer la distribution de la densité électronique à partir de l'onde diffractée. Le facteur de structure F(hkl) détermine la relation entre la réflexion correspondante et la distribution de densité électronique ρ(xyz) de la protéine.
L’obtention d’un cristal de protéine d’intérêt adapté à la diffraction des rayons X est évidemment nécessaire en tant que précurseur d’une expérience CRX. Courbe typique de la solubilité d'une protéine en fonction de sa concentration et de la concentration de l'agent précipitant. De petits agrégats de protéines peuvent alors se former : c'est le phénomène de nucléation.
Un diagramme de phases résume le comportement d'une protéine lors de la cristallisation (Fig. 16). Le problème de phase sera abordé dans la section consacrée à la solution d'une structure. Nous avons mentionné ci-dessus que le paysage conformationnel d’une enzyme dépend de la température.

Microspectrophotométrie
- Luminescence et spectroscopie
- Le cryobench
- Temps de vie de phosphorescence en fonction de la température
Le transfert d'énergie par résonance de fluorescence (FRET) peut également provoquer une diminution de l'intensité de la fluorescence. Mesurer l'absorption et la luminescence d'un fluorophore peut fournir des informations structurelles et dynamiques sur ce dernier. Compte tenu des possibilités offertes par la technologie actuelle, l'intensité et la durée de vie d'un fluorophore peuvent être mesurées.
La mesure des durées de vie de fluorescence et de phosphorescence offre ainsi la possibilité de sonder la dynamique d'un fluorophore sur les échelles de temps associées : ns et ms/s. En effet, tout événement affectant le fluorophore ou son environnement pendant la durée de vie de l'état excité est susceptible de modifier l'intensité et la durée de vie de la luminescence. Il est possible de mesurer la durée de vie de la fluorescence à l'aide d'un système TCSPC (Time-corrated Single Photon Counting) [Royant 2007].
Pour étudier la dynamique de TcAChE, et en particulier pour mettre en évidence la transition dynamique de l'enzyme, des expériences de microspectrophotométrie de fluorescence dépendante de la température (TDFM, Temperature-Derivative Fluorescence Microspectrophotometry) ont été récemment réalisées [Weik 2004]. Pour étudier la dynamique de TcAChE, nous nous sommes tournés vers une nouvelle méthode spectroscopique basée sur la mesure de la durée de vie de phosphorescence d'un ligand phosphorescent de l'enzyme AB1. En pratique, l'acquisition des données dépend de l'enregistrement (immédiatement après l'excitation du fluorophore) de l'intensité de la phosphorescence à intervalles de temps réguliers au cours de sa décroissance.
![Figure 19. Diagramme de Jablonski. Reproduit de [2].](https://thumb-eu.123doks.com/thumbv2/1bibliocom/463450.69214/88.892.254.665.215.573/figure-19-diagramme-jablonski-reproduit-2.webp)
RÉSULTATS
Acétylcholinestérase en complexe avec de putatives molécules anti-Alzheimer.84
- Article
- Bilan
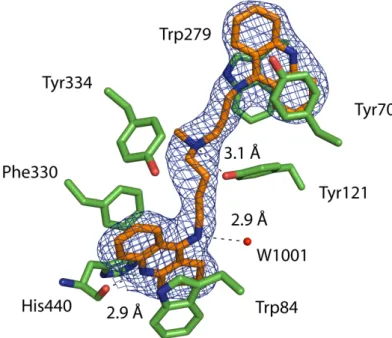
Indeed, the structure of TcAChE with an elongated bisquaternary decamethonium ligand enabled the clear identification of the PAS [Harel 1993]. Overlay of the TcAChE gorge in its native form and in complex with NF595 and NF766. Consequently, the difference observed in the adopted conformations cannot be attributed to the length of the linkers.
View of the gorge (A) and the PAS (B) in the NF1056/TcAChE complex in the orthorhombic crystal form. Co-crystals of TcAChE with NF1056 were obtained (Scheme 1) and the structure of the complex was solved at 2.3 Å resolution (Figure 3A). In the structure of the TcAChE/NF1056 complex, the peptide-like part appears to be extra-enzymatic relative to the bis-tacrine part, which is buried in the gorge.
Phe330 undergoes a conformational change: its side chain rotates ~95° relative to the structure of the native enzyme. From the bottom of the gorge to the PAS, NF1056 binds as in a complex obtained by co-crystallization of orthorhombic crystals. New anti-Alzheimer drugs can also be designed using anchor points with gorge residues in smaller conformations [Xu 2008b; Xu 2008c].
Instantanés de l’acétylcholinestérase avec le soman et le 2-PAM
- Présentation de l’article
- Article
- Bilan
The absolute configuration of the soman stereoisomer adduct in the non-aged conjugate is PSCR. This is in contrast to the total disruption of the catalytic triad observed in the non-aged VX/TcAChE conjugate [Millard 1999a]. Carbon atoms of the non-aged conjugate are shown in yellow and those of the aged conjugate in green.
Overlay of the active sites of non-age soman/TcAChE (orange) and soman/huCE1 (blue) conjugates. Comparison of the crystal structure of the non-aged soman/TcAChE conjugate with those of the non-aged VX/TcAChE conjugate [Millard 1999a] and the non-aged tabun/mAChE conjugate [Carletti 2008a] reveals an interesting difference. In the non-aged soman/TcAChE conjugate, Nε2 of its imidazole ring is within H-bonding distance of Ser 200 Oγ (2.7 Å) and O2 of the pinacolyl moiety (3.2 Å).
Attempts were made to capture and solve the 3D structure of the ternary complex of 2-PAM with the non-aged soman/TcAChE conjugate. In this context, the crystal structure of the non-aged soman/TcAChE conjugate presented here provides a valuable template for design. This alternative model of the non-aged soman/TcAChE conjugate structure has also been deposited in the PDB (accession code 2wg2).

L’acétylcholinestérase en complexe avec l’aflatoxine, un inhibiteur du PAS
- Présentation de l’article
- Article
- Bilan
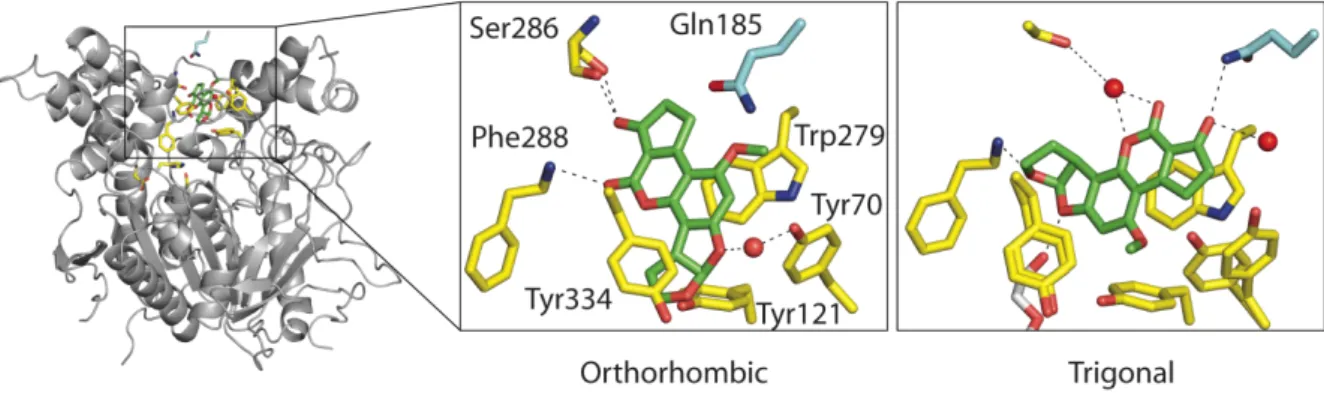
View of the PAS in the AB1/TcAChE complex obtained with orthorhombic (A) and trigonal (B) crystals (AEO and AET, respectively) and superposition of these two views (C). The structure of the AB1/TcAChE complex in the trigonal space group (called AET) was solved at 2.4 Å resolution. Superimposition with the structure of native TcAChE in the same trigonal crystal form (PDB entry code 1EA5) shows that the conformation of Gln185 is unchanged (not shown).
The change in inter-Cα distances upon binding of AB1 to trigonal crystals also involves residues within the catalytic triad. AB1 binds to the PAS in two very different orientations in two crystal forms (Fig. 2C). The observed movements all occur in the C-terminal domain of the enzyme and are relative to the N-terminal domain.
The AB1/TcAChE complex in the orthorhombic form (AEO), but not in the trigonal form (AET), undergoes a conformational change in the side chain of His440 that results in the breaking of both hydrogen bonds (Ser200Oγ-His440Nε2, His440Nδ1-Glu1327O) within the catalytic triad (Fig. 3 and Table 2). Thus, the disruption of the catalytic triad in the orthorhombic AB1/TcAChE complex can clearly be attributed to the presence of the EtOH used to solubilize the inhibitor. EtOH was able to disrupt the catalytic triad of TcAChE in the orthorhombic but not in the trigonal crystal form.

L’aflatoxine : un inhibiteur phosphorescent qui suggère un nouveau moyen
- Présentation de l’article
- Article
- Bilan
The structure of the complex of Torpedo californica (Tc) AChE with AB1 was solved in two crystal forms, revealing two distinct binding modes [Sanson]. Solvent glass transitions and protein dynamic transition were observed through characteristic changes in the temperature dependence of the phosphorescence lifetime. Evolution of the phosphorescence lifetime of AB1 as a function of temperature in complex with TcAChE in orthorhombic crystals (a), trigonal crystals (b) and in the soaking solution (c).
However, we note large differences regarding the evolution of the phosphorescence lifetime in orthorhombic crystals. Arrhenius plot of the phosphorescence lifetime of AB1 as a function of temperature in solution (blue triangles) and bound to trigonal (red circles) and orthorhombic (black squares) TcAChE. Transitions in the evolution of the phosphorescence lifetime of AB1 as a function of temperature were identified in TcAChE crystals and in the soaking solution.
Differences in the slopes of the decline of phosphorescence lifetime as a function of temperature were observed. The lowest transition temperature of the phosphorescence lifetime of AB1 is in the soaking solution (165 K). The slopes of the decrease of the phosphorescence lifetime of AB1 are greater in orthorhombic crystals than in trigonal crystals in both temperature domains.
DISCUSSION GÉNÉRALE, CONCLUSION & PERSPECTIVES
Les structures de TcAChE en complexe avec des molécules anti-Alzheimer putatives ont été résolues. 183 Les états de réponse de sénescence de TcAChE inhibés par soman ont été structurellement caractérisés [Sanson 2009]. Nous avons mis en évidence la possibilité de l'intervention d'une molécule d'eau hautement conservée dans la polarisation de la liaison O-C.
Compte tenu de la résolution de la structure (1,9 Å), nous pensons que l'allongement de cette liaison est réel. Ainsi, le vieillissement rapide de TcAChE inhibé par le soman est plus compréhensible. Les structures de TcAChE avec un inhibiteur de site périphérique, AB1, ont été résolues.
Nous proposons également de résoudre la structure du complexe TcAChE/AB1 en utilisant l'enzyme mutée au niveau de Gln185 (par exemple Q185A). Nous avons postulé que le mode de liaison biologiquement pertinent est celui de la forme orthorhombique. A cette occasion nous avons tenté de résoudre la structure du complexe TcAChE avec AB1.



![Figure 4. Les différents états de l’eau refroidie ultra-rapidement. Reproduit de [Mishima & Stanley 1998]](https://thumb-eu.123doks.com/thumbv2/1bibliocom/463450.69214/24.892.274.673.528.970/figure-états-refroidie-ultra-rapidement-reproduit-mishima-stanley.webp)