Entomological
Research Bulletin
Entomological Research Bulletin
ISSN 1015-9916
한국곤충학회지 31권 1호 2015 (특별호)
The Entomological Society of Korea Research papers
Primary Seta No. 18 of the Larval Head of Stenopsyche Species (Trichoptera: Stenopsychidae) 1 Takao Nozaki and Noriyoshi Shimura
The Genus Hydroptila Dalman (Trichoptera: Hydroptilidae) in the Ryukyu Islands, Southwestern Japan 7 Tomiko Ito
A New Species of the Genus Adicella McLachlan (Trichoptera: Leptoceridae) from Japan 18 Nobuyuki Katsuma and Eiji Yamamoto
First Record of Esakia latonota Duc and Zettel (Hemiptera: Heteroptera: Gerridae) in Thailand, 22 with a Note on the Distribution of the Genus in Oriental Region
Benjamart Suksai, Narumon Sangpradub and Chutima Hanjavanit
Two-year Aftereffects of Tsunami on Abundance of Mosquitoes in Suburban Sendai Area 27 in Miyagi Prefecture, Japan in 2013
Kimio Hirabayashi, Masaaki Takeda and Yoshio Tsuda
Macroinvertebrate Community Response to Habitat Alteration in a Regulated Mountain Stream 32 in Doi Suthep-Pui National Park, Thailand
Phuchiwan Suriyawong, Decha Thapanya, Elizabeth A. Bergey and Porntip Chantaramongkol
A List of Myanmar Caddisflies (Trichoptera) including Recently Collected Data 41 Hnin Wityi, Takao Nozaki and Takeshi Fujino
Short communications
A Rare Mayfly Siphluriscus chinensis Ulmer (Ephemeroptera: Siphluriscidae) from Vietnam 56 Van Vinh Nguyen, Van Hieu Nguyen and Yeon Jae Bae
Mayfly Fauna of the Zijin National Park as Representatives of Eastern Chinese Headwater Ephemeroptera 58 Jun-Zhi Sun, Yi-Ke Han, Juan-Yan Luo and Chang-Fa Zhou
Contents Volume 31 Number 1 2015 (Special issue)
Entomological Resear ch Bulletin
Volume 31 Number 12015(Special issue)한국곤충학회지 편집위원 (2013~2015)
편집위원장 배연재(고려대 교수)
편 집 위 원 고기성(중앙대 교수) 김익수(전남대 교수) 문명진(단국대 교수) 민경진(인하대 교수) 이훈복(서울여대 교수) 장이권(이화여대 교수) 조기종(고려대 교수) 최세웅(목표대 교수) 한연수(전남대 교수)
편 집 간 사 황정미(고려대 연구교수)
한국곤충학회지
31권 1호 (특별호)
2015. 1. 15 인 쇄 2015. 1. 30 발 행
발행인: 배연재 편집인: 배연재
발행소: 한국곤충학회/ 한국곤충연구소 136-713 서울시 성북구 안암로145 고려대학교 생명과학대학 한국곤충연구소 Tel.: 02-3290-3813, Fax.: 02-3290-3623
인쇄소: 정행사
130-814 서울시 동대문구 난계로28길48 303호 Tel.: 02-2232-3281~2, Fax.: 02-2232-5874
Entomological Research Bulletin
Volume 31 Number 1 (Special issue)
Publisher: Yeon Jae Bae Editor-in-Chief: Yeon Jae Bae
Published by
The Entomological Society of Korea Korean Entomological Institute
College of Life Sciences and Biotechnology Korea University
145 Anam-ro, Seongbuk-gu, Seoul 136-713, Korea Tel.: +82 2 3290 3813, Fax.: +82 2 3290 3623 Homepage: http://www.korentsoc.org E-mail: enrbeditor@gmail.com
Printed by Junghaeng-Sa
#303, 48, Nangye-ro 28-gil, Dongdaemun-gu, Seoul 130-814, Korea
Tel.: +82 2 2232 3281~2, Fax.: +82 2 2232 5874 Homepage: http://www.junghaengsa.co.kr Online publishing system: http://www.jops.co.kr E-mail: s3213@korea.com / jhs3213@empas.com
한국곤충학회2015 Printed in Seoul, Korea 표지사진: 2nd BSA Excursion at Hobakso and Pyochungsa Temple in Milyang, Korea
Entomological Research Bulletin
Volume 31 Number 1 2015 (Special issue) 한국곤충학회지 31권 1호(특별호)
ISSN: 1015-9916
Entomological Research Bulletin
Volume 31 Number 1 2015 (Special issue)
Aquatic Ecosystem Assessment and Management:
from Molecules to Communities
Proceedings of the 2nd Symposium of Benthological Society of Asia (BSA) June 5- - 7, 2014
Busan, Korea
Edited by
Yeon Jae Bae
The 2nd Symposium of Benthological Society of Asia (BSA), Pusan National University, Busan, Korea June 5-7, 2014
Introduction
The genus Stenopsyche is one of three genera of Stenopsychi- dae and is a genus composed of more than 80 extant species known from the eastern Palearctic and Oriental regions, except for one Afrotropical species and a European fossil species (Morse 2014). Larvae of this genus are large in body size and often abundant in number among stream macroinvertebrates, consequently they are important animals in Asian lotic eco- systems (Tanida 2002). However, ecological studies of Stenopsyche species are limited (e.g., Nishimura 1966, Aoya and Yokoyama 1987, Kocharina 1989, Dudgeon 1996, Ismail et al. 1996, Laudee and Chantaramongkol 2003). Morpho- logical knowledge of the larval stage is also scarce (Kawai 1950, Ulmer 1957, Lepneva 1964, Dudgeon 1996, Ismail et al. 1996, Hoang and Bae 2007). Inability to distinguish the species in the larval stage is a serious impediment for species- level biological investigations. Thus, taxonomic study of the larval stage, especially for means to distinguish the species, is needed not only for basic ecological studies but also for applied research such as monitoring stream environments in East and Southeast Asia.
Dr. T. Fujino, Saitama University, is conducting a survey of the aquatic insect fauna of Southeast Asia, especially Myanmar, to facilitate monitoring stream environments (Fu- jino et al. unpublished). During the course of this study, the first author of the present study examined many larval spec-
imens belonging to the genus Stenopsyche and found a pair of curious setae near the posteroventral margin of their heads.
Although these setae have not yet been mentioned by resear- chers, their shape, size and location seemed to be useful char- acters for distinguishing Stenopsyche species.
In this study, we report the morphology of this interesting seta based on examination of three Japanese Stenopsyche species, S. marmorata Navás, 1920; S. schmidi Weaver, 1987; and S. sauteri Ulmer, 1907. We also discuss the func- tion of this character.
Materials and methods
Larval specimens preserved in about 80% ethanol were used.
Collection data of three Stenopsyche species are shown in Table 1. Stenopsyche marmorata and S. schmidi are distrib- uted in Kuzuha-gawa (western Kanagawa Prefecture) and Takasato-gawa (Okinawa Island), respectively (Nozaki 1986 Kitamura unpublished). Larvae of these two species from these locations were divided into five instars by their head lengths. Final instar larvae of S. sauteri were identified by the color pattern of the head and the shape of the forecoxae (Kawai 1950, Tanida 2005).
Heads were dissected from larvae and the position and shape of primary tactile setae were observed using a Olympus SZX-12 stereomicroscope. The lengths of each head capsule Entomological Research Bulletin 31(1): 1-6 (2015)
Primary Seta No. 18 of the Larval Head of Stenopsyche Species (Trichoptera, Stenopsychidae)
Takao Nozaki1,*and Noriyoshi Shimura2
1Midorigaoka 3-16-15, Ninomiya-machi, Naka-gun, Kanagawa 259-0132, Japan
2Daisansanhaimu 106, 2-38-14, Nagatsuta, Midori-ku, Yokohama-shi, Kanagawa 226-0027, Japan
Abstract
The paired no. 18 primary setae on the larval head of three Japanese Stenopsyche species were morphologically studied. These setae arise on the ventral side of the head about 1/4th to 1/3rd of the distance from the posterior margin in the first instar and move toward the posterior margin in successive instars. Also, they are bifurcated from their bases in the 4th and 5th instars. The position of the no. 18 setae near the posterior margin of the larval head in late instars may be a synapomorphy supporting an inference of monophyly for the genus Stenopsyche. The shape, size and position of this seta and its surrounding rugged area are useful characters for distinguishing larvae of Stenopsyche species. The tactile setae probably function as a sensor to measure water current flowing into the larval retreat for efficient respiration.
Key words: larva, tactile seta, diagnostic character, function, Stenopsychelarvae, Japan
Research paper
*Correspondence
Takao Nozaki, Midorigaoka 3-16-15, Ninomiya-machi, Naka-gun, Kanagawa 259-0132, Japan E-mail: takao.nozaki@nifty.com Received 6 September 2014 Accepted 2 December 2014
and its left no. 18 seta were measured under a stereomicro- scope using an ocular micrometer. Ultrastructural studies of primary seta no. 18 and its surrounding area were also con- ducted with a JEOL JSM-6010LV scanning electron micro- scope (SEM) after sputter coating with gold using a JEOL NeoCoater MP-19020NCTR. Setal nomenclature is based on that of Williams and Wiggins (1981).
Results
Primary tactile setae of larval heads
The arrangement of primary tactile setae on the larval heads of
S. marmorata is shown in Figs. 1A and 1B. The positions of most setae except for setae no. 15 and 18 were similar in all instars. A pair of setae no. 15 were situated midlaterally on the head in the 1st instar larva and moved to the ventral side with successive instars (Figs 1A, B). Simultaneously, setae no. 18 on the ventral side about 1/4th to 1/3rd of the distance from the posterior margin in the 1st instar larva moved pos- teriorly and was situated very close to the postoccipital ridge in the 5th instar (Figs 1B, 2A, B). Setae no. 18 were forked from their bases in 4th and 5th instars (Figs 1B, 2B). The api- cal part of each seta no. 18 was occasionally bifurcated in 2nd and 3rd instars (Fig 2C). Setae no. 18 of 4th and 5th instar larvae were inconspicuous because they were short and fine
2 Entomological Research Bulletin 31(1): 1-6 (2015)
T. Nozaki & N. Shimura
Table 1.Specimens of Stenopsychespecies
Species Instars Locality Latitude and Longitude Date
S. marmorata 1st-5th Kuzuha-gawa, Hadano-shi, Kanagawa, Honshu 35°22′57′′N, 139°13′12′′E 23.vii.2013 S. schmidi 1st-5th Takasato-gawa, Ogimi-son, Okinawa 26°42′32′′N, 128°10′04′′E 14.xi.2011 S. sauteri 5th Yamaguni-gawa, Nakatsu-shi, Oita, Kyushu 33°28′57′′N, 131°09′36′′E 8.v.2014
Figure 1.Primary tactile setae on larval heads of S. marmorata(A, B) and S. schmidi(C). A, dorsal view; B & C, ventral view. I-V: 1st to 5th instar.
Scales, 0.5 mm.
I II III IV V
A
B
C
relative to their head size (Fig. 2A).
The arrangement of the primary setae of each instar in S.
schmidi was similar to that in S. marmorata, and the bifurca- tion of seta no. 18 also usually occurred in 4th and 5th instars (Fig. 1C). The bifurcated setae of this species were distinct (Fig. 2D), and easily observed by a stereomicroscope at low magnification (x10). The setation of 5th instar larvae of S.
sauteri was similar to that of S. marmorata and S. schmidi.
Seta no. 18 of S. sauteri was short like that of S. marmorata, but the location was not as close to the posterior margin in
the 5th instar as in S. marmorata (Fig. 2G). Setae no. 18 di- rected their tips anteriorly in all three species (Fig. 2).
Surrounding area of seta no. 18
Fourth and 5th instar larvae of S. schmidi had a large semi- circular area surrounding each seta no. 18 on their head in 4th and 5th instar larvae (Figs 1C, 2D-F). The area was slightly depressed, and the rugged surface was covered with dense filamentous microorganisms (mostly actinomycetes)
Primary Seta No. 18 of StenopsycheLarval Head
Figure 2.Larval heads of Stenopsychespecies. A-C: S. marmorata. A, posterior half of the head of 5th instar larva, ventral; B, left no. 18 seta of 5th instar larva, ventral; C, right no. 18 seta of 3rd instar larva, ventral. D-F: S. schmidi. D, posterior half of the head of 5th instar larva, ventral; E, left no. 18 seta of 5th instar larva, ventral; F, same, filamentous microorganisms removed. G. H: S. sauteri. G, posterior half of the head of 5th instar larva, ventral; H, left no. 18 seta of 5th instar larva, ventral. Arrow showing no. 18 seta. Scales 500μm for A. D. G; 100μm for B. E; 50μm for F. H; 10μm for C.
A D G
B E
H
C F
(Figs 2E, F). Larvae of S. marmorata also had rugged sur- face around the base of each seta no. 18 in 4th and 5th instars, but the area was very small (Fig. 2B). Fifth instar larvae of S. sauteri did not have such structure surrounding seta no.
18, although the anterior part of the setal socket was slightly wrinkled (Fig. 2H).
Discussion
Larvae of three Japanese Stenopsyche species, S. marmorata, S. sauteri and S. schmidi, each have a pair of setae arising near the posteroventral margins of larval heads in 4th and 5th instars, and those setae are apparently the primary tactile setae no. 18 (Figs 1B, C). Lepneva (1964) illustrated a larval head of S. marmorata (as Stenopsyche griseipennis McLachlan) and labeled a number of tactile setae on it. However, the seta labeled as “18” must be no. 15. Furthermore, the seta num- bered as “15” by her is seta no. 13 which did not appear in her figure (Lepneva 1964, fig. 524). Since setae no. 18 of this species are minute and inconspicuous in late instars, she probably failed to notice them.
Larvae of S. schmidi have a large semicircular depression surrounding each no. 18 seta (Figs 1C, 2D-F), but this struc- ture is vestigial in S. marmorata and absent in S. sauteri (Figs 2B, H). Ismail et al. (1996) described and illustrated the same structure in Stenospsyche siamensis Martynov, 1931 as
“black, oval and roughened area” They did not mention any setae associated with this area, but, in fact, one strong seta (not bifurcated) arises from this area in this species (Nozaki unpublished). Hoang and Bae (2007) illustrated similar semi- circular structures for six Vietnamese species including S.
siamensis. Although they also did not mention any setae on the structure, no. 18 setae probably arise there. All Stenopsy- che larvae collected from Myanmar bear a pair of setae near the posteroventral margins of the heads, and many of them have the surrounding rugged areas (Nozaki et al. in prepara- tion). Setae no. 18 situated near the posterior margin of the larval head in late instars may be a synapomorphy in this genus, a hypothesis to be tested by examination of larvae of the other two genera of the family (Pseudostenopsyche Döhler 1915, and Stenopsychodes Ulmer 1916), for both of which the larvae remain unknown. The shape, size and position of this seta and its surrounding rugged area could be useful characters for distinguishing larvae of Stenopsyche species.
These characters of three Japanese species are summarized in Table 2.
Ismail et al. (1996) suggested that “the black, oval and roughened area” on the head of S. siamensis was concerned with stridulation, analogous to the file areas in hydropsychid larvae. In our study of S. schmidi larvae, however, this
“roughened area” is always thickly grown with actinomycetes without traces of stridulation (Fig. 2E). This structure is formed simultaneously with the modification of seta no. 18 in 4th and 5th instars (Fig. 1C) and its function is more prob- ably correlated with that of the tactile seta which detects a
4 Entomological Research Bulletin 31(1): 1-6 (2015)
T. Nozaki & N. Shimura
Table 2.Characters of no. 18 tactile seta on 5th instar larval heads of three Stenopsychespecies
Species n Position Seta Surrounding Seta length (mm) Head length (mm)
rugged area average±SD average±SD
S. marmorata 10 adjacent to postoccipital ridge bifurcate vestigial 0.10±0.02 5.2±0.28 S. schmidi 10 close to postoccipital ridge bifurcate distinct, semicircula 0.22±0.03 3.8±0.10
S. sauteri 7 apart from postoccipital ridge bifurcate absent 0.13±0.02 4.6±0.18
Figure 3.Larval head of Stenopsyche schmidi. Left lateral view. Arrow shows no. 18 seta.
mechanical signal such as speed and/or direction of stream flow. Stenopsyche larvae have a very elongate head, and its ventral side is flat (Fig. 3). We think that a pair of setae di- rected anterad from the posterior margin may be an effective way to measure water current flowing past the long flat head surface, and the surrounding rugged areas may increase the sensitivity of the setae.
Yuasa (1987) observed the structure and function of cap- ture net and retreat of S. marmorata larvae in an aquarium and noted that the capture net performs the functions not only of collecting organic particles but also of introducing water into the retreat for respiration (Yuasa 1987). Larvae of Steno- psyche species do not have any tracheal gills. Freshwater efficiently introduced into their tubular retreats is important, especially for late instar larvae which have large bodies.
Differentiation between the capture net and the retreat is weak in Stenopsyche larvae (Tanida 2002). The net projecting from the entrance of the retreat may originally have evolved as a means for improving respiratory efficiency in Stenopsyche larvae.
Most Stenopsyche species are distributed in tropical or sub- tropical zones, and respiratory efficiency is more important for larvae living in warm waters. Larvae of S. schmidi living in subtropical streams have a distinct pair of no. 18 setae and surrounding depressions on their heads, but those of S. mar- morata living in temperate and subarctic streams have short no. 18 setae and vestigial surrounding depressions. Although both species belong to the same S. marmorata Species Group (Weaver 1987), these characters may be less developed in S. marmorata living in cooler waters where oxygen concen- tration is greater and respiration is easier. Stenopsyche sauteri larvae also live in temperate streams and have short no. 18 setae and no roughened areas on their heads.
Acknowledgments
We express our cordial thanks to Dr. John C. Morse, Clemson University, for critical reading of our manuscript. We are also grateful to Associate Professor Takeshi Fujino, Saitama University, for giving us the chance to examine Stenopsyche larvae collected from Southeast Asia, to Mr. Takaaki Kita- mura, Hendona High School, for the gift of larvae of Stenopsyche schmidi, and to Dr. Hiroki Sato, Forestry and Forest Products Research Institute, for the identification of actinomycetes. SEM photographs were taken at the Research and Planning Division of Kanagawa Natural Environment Conservation Center.
References
Aoya K, Yokoyama N (1987) Life cycles of two species of Sten-
opsyche (Trichoptera: Stenopsycheidae) in Tohoku district.
Japanese Journal of Limnology 48: 41-53.
Döhler W (1915) Die aussereuropäischen Trichopteren der Leip- ziger Universitäts-Sammlung. Deutsche Entomologische Zeitschrift, pp. 397-412.
Dudgeon D (1996) Life history, secondary production and micro- distribution of Stenopsyche angustata (Trichoptera: Stenopsy- chidae) in a tropical forest stream. Journal of Zoology 238:
679-691.
Hoang DH, Bae YJ (2007) Vietnamese species of Stenopsyche McLachlan (Trichoptera: Stenopsychidae). Zootaxa 1624: 1- 15.
Ismail AR, Edington JM, Green PC (1996) Descriptions of the pupae and larvae of Stenopsyche siamensis Martynov, 1931 (Trichoptera: Stenopsychidae) with notes on larval biology.
Aquatic Insects 18: 241-252.
Kawai T (1950) Notes on the larvae of the Stenopsychidae (Trichoptera). Kontyû 18: 86-88. (in Japanese)
Kocharina S (1989) Growth and production of filter-feeding caddisfly (Trichoptera) larvae in a foothill streat in the Soviet Far East. Aquatic Insects 11: 161-179.
Laudee P, Chantaramongkol P (2003) Life history of Stenopsyche siamensis Martynov 1931 (Trichoptera: Stenopsychidae) from Doi Chiang Dao catchment area, Chiang Mai Province, Thai- land, Braueria 30: 23-25.
Lepneva SG (1964) Fauna SSSR, Rucheiniki, vol. 2, no. 1. Lichin- ki I kukolki podotryada kol’chatoshchupikovykh. Zoologich- eskii Institut Akademii Nauka SSSR, ns. 88 [in Russian].
(Transrated into English as: fauna of the U.S.S.R.; Trichoptera, vol. 2, no. 2. Larva and Pupae of Annulipalpia. Israel Program for Scientific Translations, 1971).
Martynov AV (1931) Report on a collection of insects of the order Trichoptera from Siam and China. The Proceedings of the United States National Museum 79: 1-20++pls. 1-4.
Morse JC (2014) Trichoptera World Checklist. Available from:
http://entweb.clemson.edu/database/trichopt/index.htm (Accessed 24 June 2014).
Navás L (1920) [1919]. Neurópteros (Ins.) del Japón. Revista de la Real Academia de Ciencias Exactas, Físicas y Naturales de Madrid 18: 157-164.
Nishimura N (1996) Ecological studies on the net-spinning cad- disfly, Stenopsyche griseipennis MacLachlan (Trichoptera, Stenopsychidae) 1. Life history and habit. Mushi 39: 103-114
+ +3 tables.
Nozaki T (1986) Trichoptera of Kanagawa Prefecture 1. Family Stenopsychidae. Aquatic Organisms of Kanagawa Prefecture 8: 65-70. (in Japanese)
Tanida K (2002) Stenopsyche (Trichoptera, Stenopsychidae): Ecol- ogy and biology of a prominent Asian caddisfly genus. In:
Mey W (ed). Nova Supplementa Entomologica 15 (Proceedings of the 10th International Symposium on Trichoptera). pp.
595-606.
Tanida K (2005) Stenopsychidae. In: Kawai T, Tanida K (eds).
Aquatic Insects of Japan: Manual with Keys and Illustrations.
Primary Seta No. 18 of StenopsycheLarval Head
Tokai University Press, Hadano, Kanagawa, pp. 456-458.
(in Japanese)
Ulmer G (1907) Trichopteren. Part. 1. Collection of Zoologiques du Baron Edm. de Selys Longchamps, Fasc 6: 1-102.
Ulmer G (1916) Results of Dr. E. Mjöbergs Swedish scientific expeditions to Australia 1910-1913. 10. Trichoptera. Arkiv für Zoologi 10(13): 1-23.
Ulmer G (1957) Köcherfliegen (Trichopteren) von den Sunda- Iseln. Teil III. Archiv für Hydrobiologie, Supplement 23: 109- 470.
Weaver JSIII (1987) New species of Stenopsyche from the north- eastern Orient (Trichoptera: Stenopsychidae). Aquatic Insects 9: 161-168.
Williams NE, Wiggins GB (1981) A proposed setal nomenclature and homology for larval Trichoptera. In: Moretti GP (ed).
Proceedings of the 3rd International Symposium on Trichop- tera. Dr. W. Junk Publishers, The Hague. pp. 421-429.
Yuasa Y (1987) Higenaga-kawa-tobikera no suzukuri [Construc- tion of nets and retreats of Stenopsyche marmorata larvae].
Hyôgo Fresh-water Biology 26: 3-7. (in Japanese)
6 Entomological Research Bulletin 31(1): 1-6 (2015)
T. Nozaki & N. Shimura
Introduction
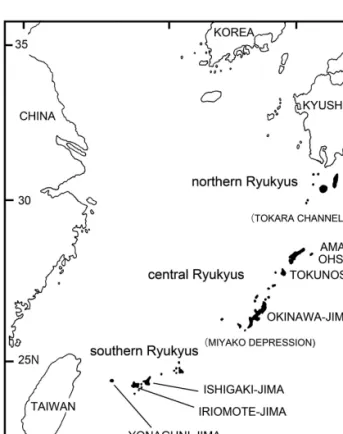
The Ryukyu Islands are the chain of islands between Taiwan and Kyushu, the southernmost of the major islands of Japan (Fig. 1), and are known to have high rates of endemism in several animal groups (Nishida et al. 2003). The islands are divided into three parts, northern, central and southern Ryu- kyus by the Tokara Channel and the Miyako Depression.
Characteristically, the fauna differs in the three part, the fauna of the northern Ryukyus is relatively similar to that of Kyushu; the southern Ryukyus have components in common with Taiwanese and southeastern Asia; and the central islands have the highest rate of the three parts (Nishida et al. 2003).
The caddisflies of the islands have been studied by many Trichoptera workers since the initial work by Akagi (1974) and Tanida (1974), and an overall account of the Trichoptera fauna of the islands was complied by Tanida (1997). He listed 44 caddisfly species and indicated high levels of end- emism, about 70%, in the central and southern Ryukyus.
Since Tanida’s study, many caddisfly groups of the islands have described new endemic species and published new records of Taiwanese and southeast Asian species (Kurani- shi 1997, Ito 2005b, 2006, Nozaki & Tanida 2006, Shimura 2010, Nishimoto 2011, Torii & Nishimoto 2011, Ito et al.
2011, 2012, 2013, Ito & Ohkawa 2012, Nozaki & Shimura 2013, Shimura et al. in press). Totally, more than 50 named caddis species and many undetermined ‘species’ have been known in this area.
This paper reviews the hydroptilid genus Hydroptila in the Ryukyus. Four species in this genus were recorded from the islands by Ito et al. (2011, Table 1). Among available mate-
rial from the central and southern Ryukyus, many new records of known species are added and three new species are described. A Taiwanese species, Hydroptila seirene Mal- icky and Chantaramongkol, 2007 is redescribed for compa- Entomological Research Bulletin 31(1): 7-17 (2015)
The Genus Hydroptila Dalman (Trichoptera: Hydroptilidae) in the Ryukyu Islands, Southwestern Japan
Tomiko Ito*
Hokkaido Aquatic Biology, Hakuyo-cho, 3-3-5, Eniwa, Hokkaido, 061-1434 Japan
Abstract
The genus Hydroptila Dalman (Trichoptera, Hydroptilidae) in the central and south- ern Ryukyus, southwestern Japan, is reviewed with descriptions of three new spe- cies; Hydroptila pseudseirene, H. spiralis and H. yaeyamensis. Four among the total of seven species are endemic to the Ryukyu Islands. For comparisons a Taiwanese species, Hydroptila seirene Malicky and Chantaramongkol, 2007 is redescribed. A clear difference is detected in the species composition of the Hydroptila fauna bet- ween the central and southern Ryukyus.
Key words: Hydroptila, Japan, new species, Ryukyu, Taiwan, Trichoptera
Research paper
*Correspondence
Tomiko Ito, Hokkaido Aquatic Biology, Hakuyo-cho, 3-3-5, Eniwa,
Hokkaido 061-1434, Japan E-mail: tobikera@siren.ocn.ne.jp Received 29 October 2014 Accepted 15 December 2014
Figure 1.Map of the Ryukyu Islands.
rison with one of the new species.
Materials and methods
Male and female genitalia were figured after treatment in about 10% KOH. Morphological terms mainly follow Mar- shall (1979) and Wells (1979). The type series of the new species are deposited in the collections of the Natural His- tory Museum and Institute, Chiba, Japan (CBM-ZI). Other specimens are deposited in the collection of the author, unless otherwise indicated in parentheses. All specimens are pre- served in 70-80% ethyl alcohol. Collectors and collection are abbreviated as follows: TI, T. Ito; M, Malaise trap: S, sweep-netting; L, light trapping using 20 watt ultra-violet light; P, light pan trap using 4 watt black light.
Species description and records
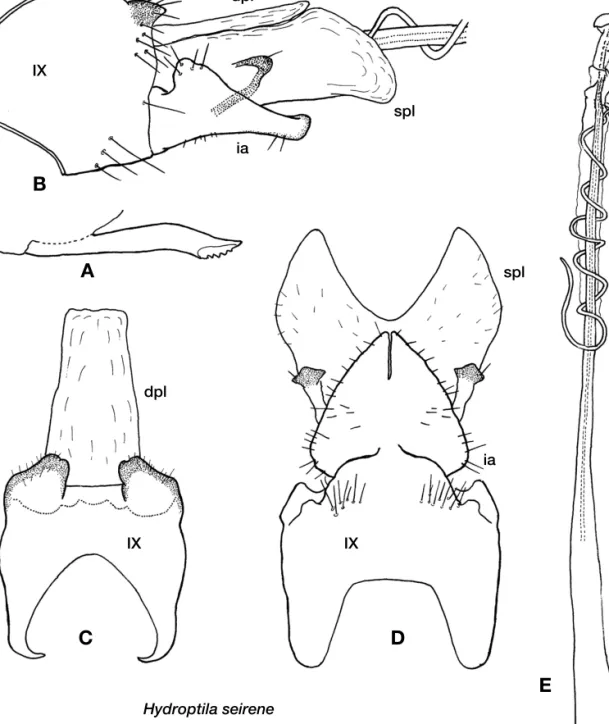
Hydroptila seirene Malicky and Chantaramongkol, 2007 (Fig. 2, Table 1)
Hydroptila seirene Malicky and Chantaramongkol, 2007, 1022, 1068. Holotype male, Taiwan.
Adult. Wings dark brown with several small lighter mark- ings. Antennae brown.
Male.Length of forewing, hind wing and body 2.2 mm, 2.0 mm and 2.3 mm, respectively (n==1). Segment number and length of antenna unknown due to specimen condition. Ven- tral process of sternite VII stout and elongate, apex with rugose sculpturing. Segment IX in lateral view large, pro- truded largely and roundly at anterior margin, with large sclerotized projections at dorso-caudal edges in ventral view.
Dorsal plate membranous and semi-transparent, often not visible, elongate rectangular in dorsal view, bar-like in late- ral view. Subgenital plate membranous, stout and rectangu- lar in lateral view, in ventral view greatly expanded laterally with wide deep excision; pair of bar-like processes near mid- dle of lateral margin, each with strongly sclerotized lozenge- shaped apex and a short seta. Phallic apparatus very elon- gate with long, spiral subapical process (not subapical in position but probably homologous with subapical process in Wells (1979)) and titillator; subapical process slender, direc- ted posteriad and twined around aedeagus, titillator directed anteriad and turned more than 5 times around aedeagus.
Inferior appendages without any hooks or appendages, fused at basal 2/3; in ventral view widest at base, gradually nar- rowed, subacute apically; in lateral view broad at base, large dorsal convexity at basal 2/5 and gradually tapered in apical half to sub-rectangular distal part.
Female.Unknown.
Specimens examined. TAIWAN: 1 male, Ilan Country,
8 Entomological Research Bulletin31(1): 7-17 (2015)
T. Ito
Table 1.Geographical distribution of Hydroptilaspecies in Ryukyu Islands, southeastern Japan, and neighbouring regions Ryukyu Islands†Other regions of Japan SpeciesAsian Far East TaiwanYonaguni-Iriomote-Ishigaki-Okinawa-TokunoAmami- KyushuShikokuHonshuHokkaidoTsushimaOgasawaraSakhalinRemarks (continental part) jimajimajimajimashimaôshima asymmetrica○○○○○○○○○○○ nanseiensis○○○endemic to Ryukyus oguranis○○○○○○endemic to Japan pseudseirene○○endemic to southern Ryukyus spiralis○○endemic to central Ryukyus thuna○○○○○ yaeyamensis○○endemic to South Ryukyus seirene○ botosaneanui○○○ chinensis○○○ coreana○○○○○ dampfi○○○ kakidaensis○○endemic to Japan ogasawaraensis○endemic to Ogasawara lslands phenianica○○○○○○○ spinosa○○○○○ Total number155313659831 †No species of Hydroptilahave been recorded in other islands.
1.5 km west of Migachyr, 1100 m, 1.XI.1996, I. Sivec (Para- type).
Remarks.This species is characterized by a very large sub- genital plate with flange-like lateral processes and broadly triangular, basally fused inferior appendages. In the original description, dorsal plate and subgenital plate are not distin- guished correctly, probably due to the almost complete trans-
parency of dorsal plate (Malicky & Chantaramongkol 2007).
Distribution.Taiwan.
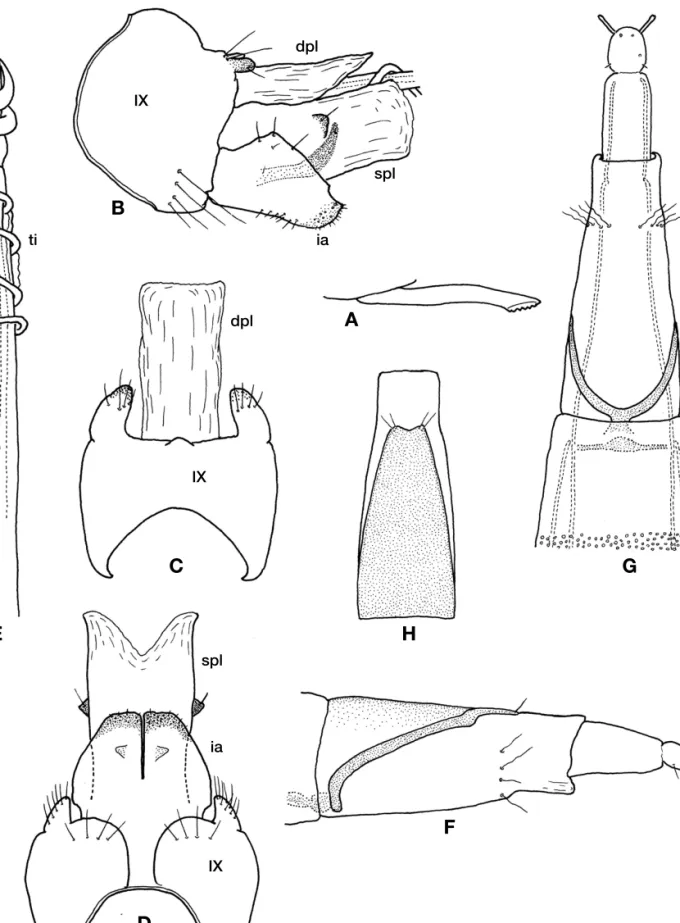
Hydroptila pseudseirene sp. nov. (Fig. 3, Table 1)
Diagnosis.The male of this species resembles a Taiwanese species, H. seirene Malicky and Chantaramongkol, 2007 in
Hydroptilid Caddisflies in the Ryukyu Islands
Figure 2.Hydroptila seireneMalicky and Chantaramongkol, 2007, male: A. ventral process of sternite VII; B. genitalia, lateral; C. same, dorsal; D.
ventral; E. phallic apparatus. Abbreviations: IX. abdominal segment IX; dpl. dorsal plate; spl. subgenital plate; ia. inferior appendage; ti. titillator.
Hydroptila seirene
C D
IX IX
ia IX
B
dpl
ia
spl
dpl
ti
A
splE
10 Entomological Research Bulletin31(1): 7-17 (2015) T. Ito
Figure 3.Hydroptila pseudseirenesp. nov. A-E, male; A. ventral process of sternite VII; B. genitalia, lateral; C. same, dorsal; D. ventral; E. phallic apparatus. F-H, female; F. abdominal segment VIII-X, lateral; G. same, ventral; H. abdominal segments VII, dorsal. Abbreviations as in Fig. 2.
Hydroptila pseudseirene
D
F
E H
C G
IX IX
dpl
spl
B
dpl
A
ti ia
spl
ia
IX
having the long phallic apparatus and broad, partly fused inferior appendages but is clearly distinguished from the latter as follows: inferior appendages in ventral view sub- rectangular apically in contrast to subtriangular in H. seirene, and subgenital plate in ventral view parallel-sided not greatly expanded laterally as in H. seirene.
Adult. Wings dark brown with several small lighter mark- ings. Antennae brown.
Male.Length of forewing, hind wing and body 1.9-2.3 mm, 1.6-2.0 mm and 1.8-2.4 mm, respectively (n==6). Antennae 27-32 segmented; length 1.0-2.7 mm (n==6). Ventral pro- cess of sternite VII stout and elongate, apex with rugose sculpturing. Segment IX large and round with round protu- berances postero-laterally, in lateral view with dorsal margin longer than ventral margin. Dorsal plate completely mem- branous and semi-transparent, often invisible, elongate- rectangular in dorsal view, parallel-sided in basal half and gradually tapered in apical half in lateral view. Subgenital plate elongate rectangular, apical half membranous and semi- transparent, often invisible, apical margin deeply and widely excised in ventral view; pair of small triangular sclerotized processes at middle of lateral margin, each with a short seta.
Phallic apparatus very elongate with spiral subapical process and long spiral titillator; subapical process strap-like and twisted around aedeagus 2 times, titillator directed anteriad, twisted around aedeagus 3-4 times. Inferior appendages broad without any hooks or appendages, fused in basal half;
in lateral view short and broad; in ventral view wide at base, gradually narrowed, sub-rectangular apically.
Female.Length of forewing, hind wing and body 2.2-2.4 mm, 1.8-2.2 mm and 2.7-3.0 mm, respectively (n==2). Ant- ennae 26-27 segmented; length 1.1-1.2 mm (n==2). Sternite VI bearing small acute ventral process. Segment VIII length greater than twice width a anterior margin; tergite weakly sclerotized, broad at basal half, gradually tapered to quad- rate apex, with shallow triangular cleft and 2 pairs of setae apically; sternite membranous with 4 pairs of sunuous setae subapically.
Holotype: Japan, Ryukyu Islands, Ishigaki-jima:1 male, Shiramizu, Nagura-gawa, unnamed tributary (24°24′44′′N, 124°11′11′′E, 95 m), 26.X.2012, TI, P (CBM-ZI 151123).
Paratypes:1 male, type locality, 27.XI.2013, TI, S (CBM- ZI 151124).
Other specimens examined. Ryukyu Islands, Ishigaki- jima:1 female, type locality, 26.X.2012, TI, P; 1 male, Arakawa, 28.XI.2013, TI, S; 1 male, Mt. Omoto-dake, 18- 19.III.2002, I. Oshima et al., L; 1 female, ibid., 26.X.2012, TI, P. Iriomote-jima: 1 male, Urauchi-gawa, mountain stream near uppermost part of Kampira-no-taki, 29.X.2012, TI, S; 1 male, Nishi-funatsuki-bashi, 23.III.1999, TI & A.
Ohkawa, S & L.
Etymology.The species name refers to similarity to H. sei- rene.
Distribution.Japan (Ryukyu Islands: Ishigaki-jima, Iriomote- jima). Endemic to southern Ryukyus.
Habitat.Adults were collected from beside streams.
Japanese name.Nise-taiwan-hime-tobikera.
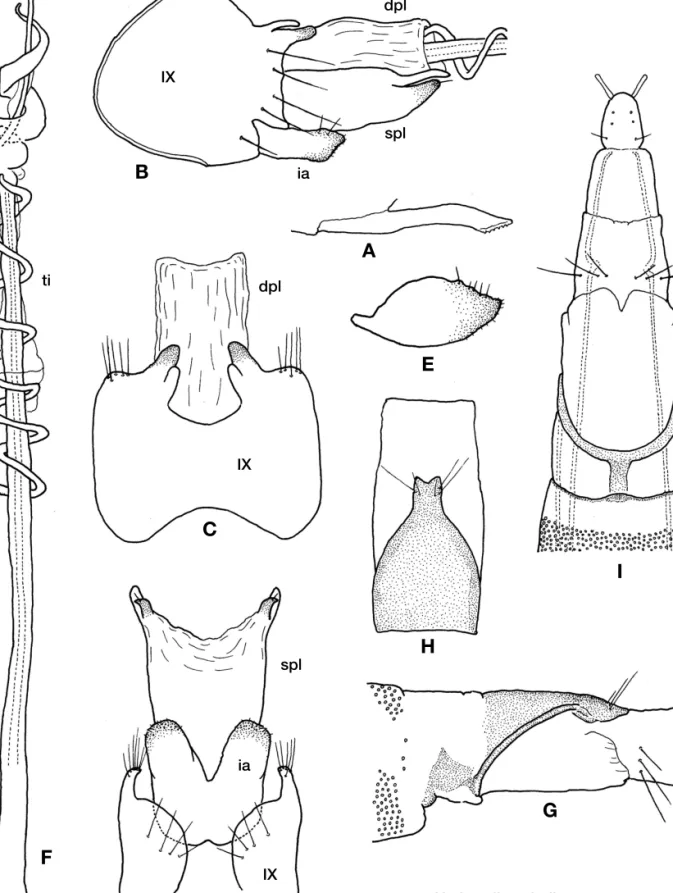
Hydroptila spiralis sp. nov. (Fig. 4, Table 1)
Diagnosis.The male of this species resembles a Taiwanese species, H. seirene Malicky and Chantaramongkol, 2007 and H. pseudseirene in having a long phallic apparatus and broad inferior appendages but is clearly distinguished from both as follows: in ventral view the apical half of inferior appen- dages is separated widely and deeply, not aligned closely as in H. seirene and H. pseudseirene, and each apico-lateral lobe of the subgenital plate is subdivided apically in this spe- cies, but not in the other two.
Adult.Wings dark brown with several small paler markings.
Antennae brown.
Male.Length of forewing, hindwing and body 2.1-2.5 mm, 1.9-2.0 mm and 2.2-2.5 mm (n==5), respectively. Antennae 27-28 segmented; length 1.2-1.3 mm (n==4). Ventral pro- cess of sternite VII stout and elongate, with fine sculptures apically. Dorsal plate completely membranous and trans- parent, often not visible, elongate-rectangular in dorsal and lateral views. Subgenital plate elongate rectangular, apical half membranous and semi-transparent, often not visible, apical margin deeply and widely concave in ventral view; in lateral view apical 1/3 subdivided, lower blade strongly sclerotized, small triangular with short apical seta. Phallic apparatus elongate with strap-like subapical process and long titillator; subapical process twined around aedeagus 2 times, titillator directed anteriad, twisted around aedeagus 5-6 times. Inferior appendages broad, relatively short, with- out any hooks or appendages, fused in basal half; apical half directed latero-caudad, spices round.
Female. Length of forewing, hindwing and body 2.5-2.8 mm, 2.2-2.3 mm and 2.8-3.3 mm, respectively (n==3). Ant- ennae 24-25 segmented; length 1.0-1.1 mm (n==2). Sternite VI bearing small acute ventral process. Segment VIII rela- tively long; tergite weakly sclerotized, broad in basal half, gradually tapered to trancate apices, with triangle cleft and 2-3 pairs of setae apically; sternite membranous with 3 pairs of setae subapically.
Holotype: Japan, Ryukyu Islands, Okinawa-jima:1 male, Kunigami-son, Yona, Yona-gawa, Heigi-hashi (26°45′41′′N, 128°13′09′′E, 33 m), 10.IV.2011, TI, S (CBM-ZI 151125).
Paratypes:3 males, 2 females, same data as holotype (CBM- ZI 151126-151130).
Other specimens examined. Ryukyu Islands, Okinawa- jima:60 males, 3 females, same data as holotype; 11 males, 15 females, Kin-cho, Okukubi, Okukubi-gawa, headwater, 1.
III.2011, R. B. Kuranishi (deposited in the collection of R. B.
Kuranishi). 1 female, Kunigami-son, Nishime-dake, Zatsu-
Hydroptilid Caddisflies in the Ryukyu Islands
12 Entomological Research Bulletin31(1): 7-17 (2015) T. Ito
Figure 4.Hydroptila spiralissp. nov. A-F, male; A. ventral process of sternite VII; B. genitalia, lateral; C. same, dorsal; D. ventral; E. inferior appendage, variation, lateral; F. phallic apparatus. G-I, female; G. abdominal segments VIII-X, lateral; H. same, dorsal; I. abdominal segment VIII, ventral. Abbreviations as in Fig. 2.
Hydroptila spiralis
C
D
ti
F
IX
ia dpl
IX
E
H
dpl
spl
B
iaG
I A
spl IX
gawa, headwater, 2.VII.1997, R. B. Kuranishi (deposited in the collection of R. B. Kuranishi).
Amami-oshima: 1 female, Uken-son, Kawauchi-gawa, 50 m, 26.III.2007, TI, L; 1 male, Yamato-son, Materia-no-taki, 27.III.1998, H. Nishimoto.
Etymology.Specific name refers to the shape of the phallic apparatus.
Distribution.Japan (Ryukyu Islands: Amami-oshima, Oki- nawa-jima). Endemic to central Ryukyus.
Habitat.Many adults were collected from rocky fast-flowing streams.
Japanese name.Rasen-hime-tobikera.
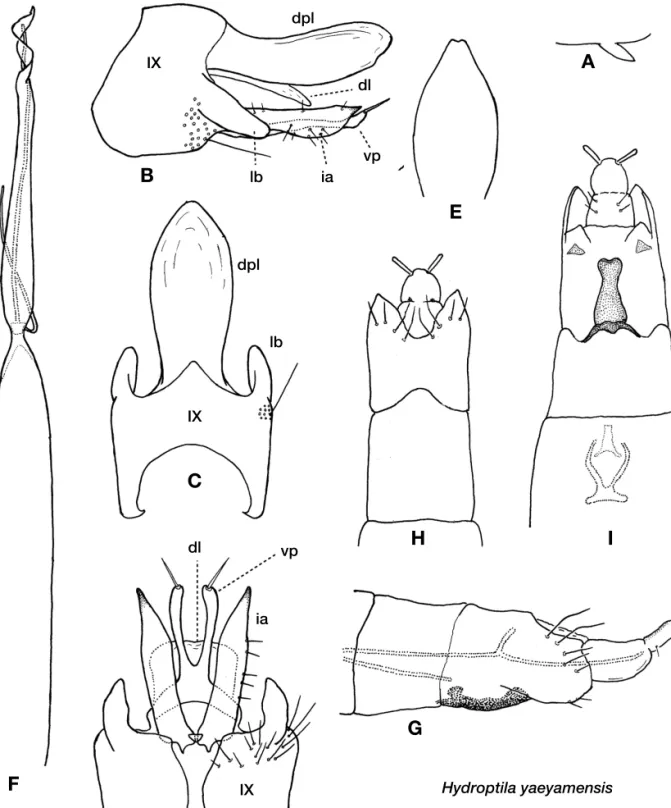
Hydroptila yaeyamensis sp. nov. (Fig. 5, Table 1)
Diagnosis.The male of this species is similar to that of Hyd- roptila pythia Malicky and Chantaramongkol, 2007, des- cribed from Sumatra, having the dorsal plate ovoid, but is clearly distinguishable from the latter by the divided ventral process of subgenital plate.
Adult.Wings dark brown with several small light markings.
Antennae brown.
Male.Length of forewing, hind wing and body 1.7-2.0 mm, 1.4-1.6 mm and 1.9-2.2 mm (n==4), respectively. Antennae 27-segmented; length 1.0 mm (n==1). Ventral process of ster- nite VII small, acute apically. Segment IX sub-rectangular in ventral view, with large semi-membranous lateral lobes, lobes directed ventro-caudad apices subacute. Dorsal plate semi-membranous, oval in dorsal view, with very small exci- sion apically in some specimens. Subgenital plate consisting of membranous dorsal lobe and weakly sclerotized ventral processes; dorsal lobe much shorter than dorsal plate, thin bar-like in lateral view, sub-rectangular in ventral view; ven- tral processes almost as long as dorsal plate, deeply divided at basal 1/3 with a conspicuous seta apically. Phallic appa- ratus very elongate, twisted twice apically, short titillator curled once around aedeagus and directed caudad. Inferior appendages longate, straight, almost as long as ventral pro- cess of subgenital plate, parallel-sided in basal 2/3 and gra- dually tapered in apical 1/3, apices acute.
Female.Length of forewing, hind wing and body 1.6-2.1 mm, 1.4-1.8 mm and 1.8-2.3 mm, respectively (n==4). Ant- ennae 19-20 segmented; length 0.6-0.7 mm (n==2). Sternite VI bearing small acute ventral process. Tergite VIII weakly sclerotized, large with deep wide concavity at distally bor- dered by 5 pairs of marginal setae, 2 median pairs longer than others. Ventral sclerite of segment VIII heavily sclero- tized, elongate-rectangular with gently concave lateral mar- gins, wide anterior margin and roundish posterior margin.
Holotype: Japan, Ryukyu Islands, Ishigaki-jima:1 male, Shiramizu, Nagura-gawa, unnamed tributary (24°24′44′′N, 124°11′11′′E, 95 m), 26.X.2012, TI, P (CBM-ZI 151131).
Paratype:1 male, Ishigaki-jima, Shiramizu, Nagura-gawa,
8 m, unnamed bridge, 25.X.2012, TI, P (CBM-ZI 151132).
Other specimens examined. Ryukyu Islands, Ishigaki- jima:1 male, Ishigaki-jima, Shiramizu, Nagura-gawa, tri- butary, unnamed bridge, 8 m, 25.X.2012, TI, P; 1 female, Shiramizu, small tributary, 11-13.IV.2011, TI, P; 1 female, Miyara-gawa, upper reach of Maezato-dam, 27.X.2012, TI, P. Iriomote-jima: 1 male, Aira-gawa, 3 m, 28-30.X.2012, TI, P; 1 female, Omija-gawa, Omija-bashi, 29.X.2012, TI, P; 1 female, Nishi-funatsuki-gawa, Nishi-funatsuki-bashi, 28.XI.2013, TI, S.
Etymology.Specific name derived from the name of the islands where this species were collected.
Distribution. Japan (Ryukyu Islands: Ishigaki-jima, Irio- mote-jima). Endemic to southern Ryukyus.
Habitat.Adults were collected from beside streams.
Japanese name.Yaeyama-hime-tobikera.
Hydroptila thuna Oláh, 1989 (Table 1)
Hydroptila thuna Oláh, 1989, 281-282, fig. 26, male, Viet- nam.
Hydroptila triangularis Wells and Dudgeon, 1990, 168- 169, figs. 14-16, male, female, Hong Kong; synonymi- zed by Wells and Malicky (1997).
Hydroptila apiculata Yang and Xue, 1992, 26-27, male, China; Arefina, 2004, 210-211, figs. 6-14, male, female, Russia (Khabarovsk, Primorye); synonymized by Malicky and Chantaramongkol (2007).
Hydroptila molione Malicky, 2004b, 293-294, fig. 3, male, Nepal; synonymized by Malicky and Chantaramongkol (2007).
Hydroptila phenianica: Shimura, 2010, 49, Japan (Yonaguni Island), misidentification.
Hydroptila thuna: Ito et al. 2011, 4-5, male, female, Japan.
Specimens examined.Records additional to those of Ito et al. (2011). Ryukyu Islands: Ishigaki-jima: 1 male, 2 fe- males, Shiramizu, Nagura-gawa, Tôre-bashi, 25.X.2012, TI, L; 11 males, 33 females, Shiramizu, Nagura-gawa, un- named bridge, 8 m, 25.X.2012, TI, P; 1 female, Shiramizu, Nagura-gawa, tributary, 18 m, 26.X.2012, TI, P; 1 female, Shiramizu, Nagura-gawa, small tributary, 11-13. IV.2011, TI, L; 1 male, 77 females, Miyara-gawa, dam-ue, 55 m, 27.
X.2012, TI, P; 31 males, 703 females, Miyara-gawa, dam shita, 40 m, 27.X.2012, TI, P. Iriomote-jima: 1 female, Aira-gawa, 3 m, 11-13.IV.2011, TI, P.
Distribution. Japan (Ryukyu Islands: Yonaguni-jima, Ishigaki-jima, Iriomote-jima), Russian Far East (continental parts), China, Taiwan, Hong Kong, Raos, Thailand, Viet- nam, Nepal, Sumatra. New to Iriomote-jima.
Japanese name.Ajia-hime-tobikera.
Hydroptila phenianica Botosaneanu, 1970 (Table 1) Hydroptila phenianica Botosaneanu, 1970, 290-291, pl. 12,
Hydroptilid Caddisflies in the Ryukyu Islands
male, female, Korea: Arefina et al., 2002, 99, Russian Far East (continental parts); Shimura et al., in press, Japan (Ryukyu Islands: Amami-oshima).
Hydroptila matsuii Kobayashi, 1974, 67-68, pl. 25, male,
Japan (Honshu): Ito, 2005a, 441, 446, figs. 7-8, male;
synonymized by Nozaki and Tanida (2007).
Distribution.Japan (Hokkaido, Honshu, Shikoku, Kyushu,
14 Entomological Research Bulletin31(1): 7-17 (2015)
T. Ito
Figure 5.Hydroptila yaeyamensissp. nov. A-F, male; A. ventral process of sternite VII; B. genitalia, lateral; C. same, dorsal; D. ventral; E. dorsal plate, variation, dorsal; F. phallic apparatus. G-I, female; G. abdominal segments VII-X, lateral; H. same, dorsal; I. abdominal segments VIII-X, ven- tral. Abbreviations: dl. dorsal lobe of subgenital plate; vp. ventral process of subgenital plate; lb. lateral lobe of segment IX; others as in Fig. 2.
Hydroptila yaeyamensis
IX
A
ti
B
lb iadl
vp
IX
C
dl
E
H I
F
D
G
IX dpl
lb
vp
ia dpl
Tsushima Islands, Ryukyu Islands: Amami-oshima). Korea, Russian Far East (continental parts).
Japanese name.Matsui-hime-tobikera.
Hydroptila oguranis Kobayashi, 1974 (Table 1)
Hydroptila oguranis Kobayashi, 1974, 68-69, pl. 25, male, Japan; Ito et al., 1998, figs. 3-4, larva, case; Ito, 2005a, 441, 446, fig. 21, male; Ito et al. 2011, 8-11, male, female, Japan.
Hydroptila sp. Tanida et al., 1999, 394.
Distribution.Japan (Hokkaido, Honshu, Shikoku, Kyushu, Tsushima Islands, Ryukyu Islands: Amami-oshima). Ende- mic to Japan.
Japanese name.Ogura-hime-tobikera.
Hydroptila nanseiensis Ito, 2011 (Table 1)
Hydroptila nanseiensis Ito et al. 2011, 14-15, male, female, Japan.
Specimens examined.Additional records to Ito et al. (2011).
Ryukyu Islands: Okinawa-jima: 59 females, Nago-shi, Genka-kawa, Hogen-hashi, 8.IV.2011, TI, L & P; 4 males, 33 females, ibid., 17-19.X.2014, TI, L & P; 4 femlaes, Nago-shi, Genka-kawa, tributary, 8.IV.2011, TI, P; 31 females, Kunigami-son, Okuma-gawa, 12 m, 9.IV.2011, TI, P; 1 female, Kunigami-son, Shinrin-koen, pond, 10.IV.2011, TI, L; 1 female, Kunigami-son, Yona-gawa, Heigi-hashi, 10.IV.2011, TI, S; 2 males, 53 females, Kunigami-son, Yona- gawa, Nakafukuji-hashi, 18.X.2014, TI, L. Ishigaki-jima:
255 males, 477 females, Shiramizu, Nagura-gawa, tributary, 8 m, 25.X.2012, TI, P; 7 males, 34 females, Shiramizu, Nagura-gawa, tributary, 28 m, 25.X.2012, TI, L; 2 females, ibid., 27.XI.2013, TI, P; 14 males, 39 females, ibid., 26- 28.XI.2013, TI, L; 29 males, 88 females, Shiramizu, Nagura- gawa, small tributary, 15 m, 11-13.IV.2011, TI, L, P & S; 3 males, 1 female, Nagura-gawa, tributary, 95 m, 26.X.2012, TI, P; 4 males, 2 females, ibid., TI, S; 3 males, 6 females, Nagura-gawa, dam ue, 70 m, 11.IV.2011, TI, P; 13 males,11 females, ibid., 27.XI.2013, TI, L; 17 males, 8 females, Miyara-gawa, Maezato dam shita, 40 m, 27.X.2012, TI, P &
S; 2 females, Miyara-gawa, Maezato dam ue, 55 m, 27.X.
2012, TI, P; 3 males, Omoto-dake, 155-195 m, 26.X.2012, TI, P. Iriomote-jima: 57 males, 309 females, Aira-gawa, 3 m, 28-30.X.2012, P; 3 males, ibid., 28.X-2.XII.2013, TI &
R. Saito, M; Aira-gawa, about 35 m, 1.XII.2013, TI, P; Aira- gawa, 44 m, 1.XII.2013, TI, L & S; 2 males, 8 females, Omija-gawa, Omija-bashi, 29.X.2012, TI, P; 2 males, 4 females, ibid., 30.XI.2013, TI, S & L.
Distribution. Japan (Ryukyu Islands: Okinawa-jima, Ishigaki-jima, Iriomote-jima). Endemic to Ryukyu Islands.
Most abundant hydroptilid species in the Ryukyus.
Habitat.Adults were collected alongside streams in moun-
tain and lowland areas.
Japanese name.Nansei-hime-tobikera.
Hydroptila asymmetrica Kumanski, 1990 (Table 1)
Hydroptila asymmetrica Kumanski, 1990, 50-52, figs. 56- 63, male, female, Korea; Arefina et al., 2002, 97, Russian Far East (continental parts); Nozaki and Tanida, 2007, 245, Japan (Honshu); Ito et al. 2011, 15-18, male, female, Japan; Shimura in press, Japan (Ryukyu Islands: Amami- oshima).
Specimens examined.Records additional to those of Ito et al. (2011). Ryukyu Islands: Amami-oshima: 2 males, 7 females, Uken-son, Kawauchi-gawa, 50 m, 24.X.2011, TI, P; 63 males, 203 females, 20.X.2014, TI, L & P; 1 male, Yamato-son, Materia-no-taki, 25-26.X.2011, TI, S; 199 males, 212 females, Materia-gawa, upper reach, 21.X.2014, TI, L & P. Tokunoshima: 1 female, Amagi-cho, Nishi- agina, 13.V.2014, K. Tojo, L. Okinawa-jima: 83 males, 131 females, Nago-shi, Genka-kawa, Hogen-hashi, 8.IV.2011, TI, L & P; 21 males, 10 females, ibid., 17-19.X.2014, TI, L &
P; 3 males, 7 females, Nago-shi, Genka-kawa, tributary, 8.
IV.2011, TI, P; 11 males, 6 females, Nago-shi, Genka-kawa, upper reach, 26.V.1997, R. B. Kuranishi (deposited in the collection of R. B. Kuranishi); 1 male, 4 females, Kunigami- son, Okuma-gawa, 12 m, 9.IV.2011, TI, P; 1 male, 11 females, Kunigami-son, Yona-gawa, Heigi-hashi, 10.IV.
2011, TI, P; 41 males, 37 females, Kunigami-son, Yona- gawa, Nakafukuji-hashi, 18.X.2014, TI, L.
Distribution.Japan (Hokkaido, Honshu, Shikoku, Kyushu, Tsushima Islands, Ryukyu Islands: Amami-oshima, Okinawa- jima, Ishigaki-jima, Iriomote-jima), Korea, Russian Far East (continental parts).
Habitat.Adults were collected from beside streams in moun- tain and lowland areas.
Japanese name.Migi-hime-tobikera.
Hydroptila sp.
This appears to be an undescribed species collected in Okinawa-jima and will be dealt with when more material becomes available.
Specimens examined. 1 male, Okinawa-jima, Nago-shi, Genka-kawa, Hogen-hashi, 8.IV.2011, TI, L.
Discussion
Seven species of the genus Hydroptila are now known in the central and southern Ryukyus (Table 1). Four of these, H. nanseiensis, H. pseudseirene, H. spiralis and H. yaeya- mensis, are endemic to the islands. Close relatives of the
Hydroptilid Caddisflies in the Ryukyu Islands
four are found in southeastern Asia including Taiwan, sug- gesting that ancestor of the four species may have derived from there. Clear differences are apparent in the Hydroptila fauna of between central Ryukyus (H. spiralis) and southern Ryukyus (H. thuna, H. pseudseirene, H. yaeyamensis), although two species, H. nanseiensis and H. asymmetrica, are commonly distributed in the islands of both areas. H.
thuna found in both southern Ryukyus and Taiwan.
High endemism among the Trichoptera of central and southern Ryukyus has been observed in 9 caddisfly genera;
Hydropsyche Pictet (Hydropsychidae; 1 species endemic to Ryukyus/2 species distributed in Ryukyus in total; Tanida 1986a, 1986b, 1987), Lepidostoma Ramber (Lepidosto- matidae; 8/10; Ito 2005b), Goera Stephens (Goeridae; 2/4;
Nozaki & Tanida 2006), Molanna Curtis (Molannidae; 1/1;
Ito 2006), Eoneureclipsis Kimmins (Psychomyiidae; 2/2;
Torii & Nishimoto 2011), Paduniella Ulmer (Psychomyi- idae; 1/2; Nishimoto 2011), Adicella McLachlan (Leptoce- ridae; 1/2; Ito et al. 2012), Ugandatrichia Mosely (Hydrop- tilidae; 1/2; Ito & Ohkawa 2012), and Oxyethira Eaton (Hyd- roptilidae; 1/1; Oláh & Ito 2013). There exist high faunal differences between central and southern Ryukyus in Tri- choptera genera. Although some species in central Ryukyu are common to the adjacent regions, a number of species in southern Ryukyus are endemic to the islands as in the genus Hydroptila; 7 of the genera have species in common with Taiwan, Eoneureclipsis and Hydropsyche in the southern Ryukyus being the exceptions. Characteristics of the Hyd- roptila fauna are almost completely in accord with those shown in previous studies.
Acknowledgments
I am sincerely grateful to Alice Wells, Australian Biolo- gical Resources Study, Canberra, Australia, for her kind advice on the study and critical reading of my manuscript.
For loan or gift of valuable specimens my thanks are also due to Hans Malicky, Austria; Ryoichi B. Kuranishi, Natural History Museum and Institute, Chiba; Koji Tojo, Shinshu University; and Hiroyuki Nishimoto, Komaki-shi, Aichi;
and for their help on my collecting trips I am grateful to Takaaki Kitamura, Okinawa Hentona High School, and Rie Saito, Shinshu University. I am thankful to Takao Nozaki, Ninomiya-machi, Kanagawa, for his help on making figure files.
References
Akagi I (1974) A larva of Goera from Amami-ohshima Island.
Annuals of Kansai Natural Science Association 26: 18-19 [in Japanese].
Arefina TI (2004) A new species of the genus Stactobiella Mar- tynov with reassignment of Stactobiella tshistjakovi (Arefina et Morse, 2002) and new records of micro-caddisflies (Tri- choptera: Hydroptilidae) from the Russian Far East. Euro- asian Entomological Journal 3(3): 209-211.
Arefina TI, Vshivkova TS, Morse JC (2002) New and interesting Hydroptilidae (Insecta: Trichoptera) from the Russian Far East. In: Mey W (ed), Proceedings of 10th International Symposium of Trichoptera (Nova Supplementa Entomologica 15), pp. 96-106. Goecke & Evers, Keltern.
Botosaneanu L (1970) Trichoptères de la République Démocra- tique-Populaire de la Corée. Annales Zoologici 27: 275-359.
Ito T (2005a) Hydroptilidae. In: Kawai T, Tanida K (eds), Aqua- tic Insects of Japan: Manual with Keys and Illustrations. pp.
440-446. Tokai University Press, Kanagawa [in Japanese].
Ito T (2005b) Checklist of the family Lepidostomatidae, Trichop- tera, in Japan 2. In: Tanida K, Rositer A (eds) Proceedings of the 11th Trichoptera Symposium, pp. 199-206. Tokai Uni- versity Press, Kanagawa.
Ito T (2006) A new species of the genus Molanna Curtis (Tri- choptera, Molannidae) from the Yaeyama Islands, the south- ernmost part of Japan. Limnology 7: 205-211.
Ito T, Hayashi Y, Shimura N (2012) The genus Anisocentropus McLachlan (Trichoptera, Calamoceratidae) in Japan. Zootaxa 3157: 1-17.
Ito T, Kamei H, Ohkawa A, Kuhara N, Nishimoto H (1998) Cad- disfly fauna of the Shibetsu District and the Shiretoko Pass, easternmost part of Hokkaido, northern Japan. Biology of Inland Waters 13: 1-17 [in Japanese with English abstract].
Ito T, Kuhara N, Katsuma N (2013) The genus Adicella McLach- lan (Trichoptera, Leptoceridae) from Japan. Zootaxa 3635:
27-39.
Ito T, Ohkawa A (2012) The genus Ugandatrichia Mosely (Tri- choptera, Hydroptilidae) in Japan. Zootaxa 3394: 48-58.
Ito T, Ohkawa A, Hattori T (2011) The genus Hydroptila (Tri- choptera, Hydroptilidae) in Japan. Zootaxa 2801: 1-26.
Kobayashi M (1974) On two new species of Hydroptilidae from Japan (Insecta: Trichoptera). Bulletin of the Kanagawa Pre- fectural Museum (Natural Science) 7: 67-70, pls. 25-26.
Kumanski K (1990) Studies on the fauna of Trichoptera (Insects) of Korea. I. Superfamily Rhyacophiloidea. Historia naturalis bulgarica 2: 36-59.
Kuranishi RB (1997) The genus Rhyacophila of the Ryukyu Archipelago, Part I (Trichoptera: Rhyacophilidae). In: Hol- zenthal R, Flint OS Jr (eds) Proceedings of the 8th Interna- tional Symposium on Trichoptera, pp. 265-269. Ohio Biolo- gical Survey, Ohio.
Malicky H (2004) Neue Köcherfliegen (Trichoptera) aus dem Bardia Nationalpark, Nepal. Denisia 13: 291-300.
Malicky H, Chantaramongkol P (2007) Beiträge zur Kenntnis asiatischer Hydroptilidae (Trichoptera). Linzer biologische Beitrage 39: 1009-1099.
Marshall JE (1979) A review of the genera of the Hydroptilidae (Trichoptera). Bulletin of the British Museum (Natural His-
16 Entomological Research Bulletin31(1): 7-17 (2015)
T. Ito
tory) 39: 135-239.
Nishida M, Shikatani N, Shokita S (eds) (2003) The flora and fauna of inland waters in the Ryukyu Islands. Tokai Univer- sity Press, Tokyo [in Japanese].
Nishimoto H (2011) The genus Paduniella (Trichoptera: Psycho- myiidae) in Japan. Zoosymposia 5: 381-390.
Nozaki T, Shimura N (2013) Two polycentropodid caddisflies (Trichoptera, Insecta) collected from Yonaguni-jima, west- ernmost Japan. In: Tanida K, Tojo K, Nozaki T (eds) Biology of Inland Water, Supplement 2 (Proceedings of the 1st Sym- posium of the Benthological Society of Asia), pp. 101-108.
Scientific Research Society of Inland Water Biology, Kana- gawa.
Nozaki T, Tanida K (2006) The genus Goera Stephens (Trichop- tera: Goeridae) in Japan. Zootaxa 1339: 1-29.
Nozaki T, Tanida K (2007) The caddisfly fauna of a huge spring- fed stream, the Kakida River, in central Japan. In: Bueno- Soria J, Barba-Ávlarez R, Armitage B (eds) Proceedings of the 12th International Symposium on Trichoptera, pp. 243- 255. The Caddis Press, Ohio.
Oláh J (1989) Thirty-five new hydroptilid species from Vietnam (Trichoptera, Hydroptilidae). Acta Zoologica Hungarica 35:
255-293.
Oláh J, Ito T (2013) Synopsis of Oxyethira flavicornis species group with new Japanese Oxyethira species (Trichoptera, Hydroptilidae). Opuscula Zoologica, Budapest 44: 23-46.
Shimura N (2010) Collection record of Ephemeroptera, Plecop- tera and Trichoptera from Yonaguni-Island, the westernmost part of Japan. Hyôgo Freshwater Biology 61/62: 45-54 [in Japanese].
Shimura N, Yoshinari G, Morimoto S, Tokoro T, Komori C. Col- lection records of aquatic invertebrates from Amami Island, Kagoshima Prefecture. Hyôgo Freshwater Biology [in Japan-
ese]. In press.
Tanida K (1974) Preliminary reports on the aquatic insects of the Ryukyus with some ecological remarks I. Ecological Studies on Nature Conservation of Ryukyu Islands, 1: 161-174 [in Japanese].
Tanida K (1986a) A revision of Japanese species of the genus Hydropsyche (Trichoptera, Hydropsychidae) I. Kontyû, Tokyo 54: 467-484.
Tanida K. (1986b) A revision of Japanese species of the genus Hydropsyche (Trichoptera, Hydropsychidae) II. Kontyû, Tokyo 54: 624-633.
Tanida K (1987) A revision of Japanese species of the genus Hydropsyche (Trichoptera, Hydropsychidae) III. Kontyû, Tokyo 55: 59-70.
Tanida K (1997) The Trichoptera fauna of the Ryukyu Islands:
taxonomic and ecological prospect. In: Holzenthal R, Flint, OS Jr (eds) Proceedings of the 8th International Symposium on Trichoptera, pp. 445-451. Ohio Biol. Survey, Ohio.
Torii T, Nishimoto H (2011) Discovery of the genus Eoneurec- lipsis Kimmins (Trichoptera: Psychomyiidae) from Japan.
Zoosymposia 5: 453-464.
Wells A (1979) A review of the Australian species of Hydroptila Dalman (Trichoptera: Hydroptilidae) with descriptions of new species. Australian Journal of Zoology 26: 745-762.
Wells A, Dudgeon D (1990) Hydroptilidae (Insecta: Trichoptera) from Hong Kong. Aquatic Insects 12: 161-175.
Wells A, Malicky H (1997) The micro-caddisflies of Sumatra and Java (Trichoptera: Hydroptilidae). Linzer biologische Beitrage 29: 173-202.
Yang L, Xue Y (1992) Six new species of Hydroptilidae (Insecta:
Trichoptera) from China. Entomotaxonomia 14(1): 26-34 [in Chinese with English description of the species].
Hydroptilid Caddisflies in the Ryukyu Islands
Introduction
The genus Adicella McLachlan, 1877 (Trichoptera, Lepto- ceridae) is a genus composed of about 140 species from the Oriental, Afrotropical and West and East Palearctic biogeo- graphic regions (Morse 2014). In Japan, Ito et al. (2013) revi- ewed Japanese Adicella and recognized 4 species (A. makaria Malicky and Chantaramongkol, 2002, A. trichotoma Ito and Kuhara, 2013, A. paludicola Ito and Kuhara, 2013 and A.
strigillataKatsuma and Ito, 2013) for Japanese fauna. On the other hand, the second author of this paper found a lepto- cerid males, and it’s wing venation is very similar to that of males of the genus Erotesis. After precise study of this inter- esting species, we recognized that this species belongs to the genus Adicella. In this paper, we describe the male and female of a new species from Japan.
Materials and methods
Male and female genitalia were illustrated after treatment in hot 10% KOH. Morphological terminology follows that of Yang and Morse (2000). The type specimens are deposited in the collection of the Natural History Museum and Institute of Chiba (CBM). Other specimens are deposited in my own collection (NK). Abbreviations as follows of Fig. 1. Male, IX and X: abdominal segment IX, and X, lo X: lower part of tergum X, up X: upper part of tergum X, pre ap: preanal appendage, inf app: inferior appendage, phb: phallobase, phc:
phallicata. Female, VIII, IX and X: abdominal segment VIII, IX and X; pr ap: preanal appendage; go pl: gonopod plate;
vl go pl: vertical lobe of gonopod plate; ap go pl: apicomes- sal process of internal gonopod plate; sp sc: spermathecal sclerite.
Taxonomic accounts Family Leptoceridae Genus Adicella
Adicella odamiyamensis sp. nov.
Japanese name. Odamiyama-ko-higenaga-tobikera.
(Figs 1-3)
Material examined. Holotype. (in alcohol), Japan. Oda- miyama-keikoku, Uchiko-cho, Ehime (132°53′37′′E, 33°31′ 49′′N alt. 800 m), 8.ix.2000, E. Yamamoto (CBM-ZI151133).
Paratypes. 1 (in alcohol), Japan. Yurano, Kumakogen-cho, 10.vii.2007, E. Yamamoto (CBM-ZI151135), 1♀(in alco- hol), same data as holotype (CBM-ZI151134). 1 (in alco- hol), Odamiyama-keikoku, Uchiko-cho, alt. 800 m, 15.viii.
2000, E. Yamamoto (CBM-ZI151136). Other specimens examined. Ehime: Japan. 1♀, Odamiyama-keikoku, Uchiko- cho, 27.viii.2000, E. Yamamoto (NK), 3♀, ditto, 2.ix.2000, E. Yamamoto (NK), 1♀, ditto, 17.ix.2000, E. Yamamoto (NK), 1 , Namakusa-dani, Odamiyama, Uchiko-cho, alt.
1200 m, 8.viii.2000, E. Yamamoto (NK), 1♀, ditto, 8.ix.
2000, E. Yamamoto (NK), 1♀, ditto, 17.ix.2000, E. Yama- moto (NK).
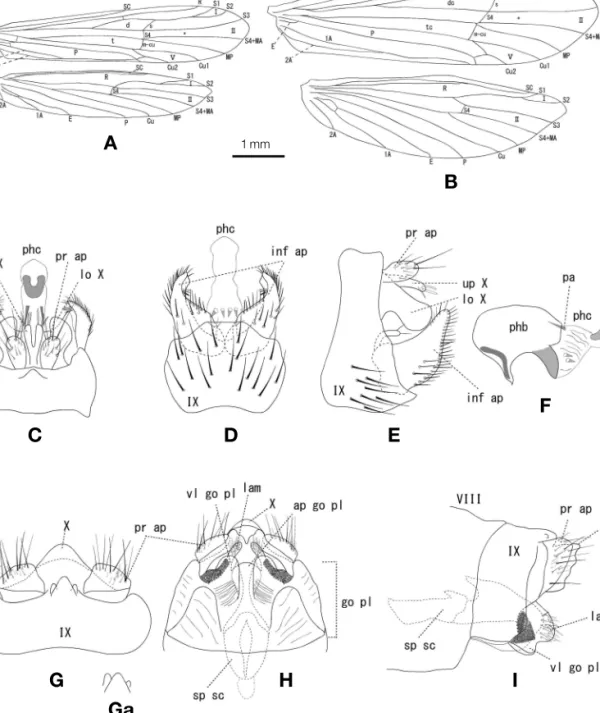
Description. Adult.Head and body tawny. Forewings brown, length 4.1-5.0 mm (n==4) in male, 6.3-6.8 mm (n==6) in female covered with brown hairs but almost rubbed in alco- hol. In male forewings, venation similar to that of genus Erotesis, anastomosis not in a continuous oblique line such as those of other Japanese Adicella species (Fig. 1A). Anten- nae long and slender, about twice as long as forewings.
Male genitalia(Figs 1C-F). Segment IX subtrapezoidal in lateral view, with small triangular lobe posterodorsally; pos- teroventral margin convex in ventral view. Preanal append-
A New Species of the Genus Adicella McLachlan (Trichoptera:
Leptoceridae) from Japan
Nobuyuki Katsuma1,*and Eiji Yamamoto2
1Environmental Research Center Co. Ltd., 3-1, Hanare, Tsukuba, Ibaraki 305-0857, Japan
2Yamamoto Institute of Forest Biology, 1127, Yoshinokawa, Uchiko-cho, Ehime 791-3511, Japan
Abstract
A new leptocerid caddisfly Adicella odamiyamensis sp. nov. is described from Japan.
Key words: taxonomy, caddisfly, East Palearctic, Adicella eratoSpecies Group
Research paper
*Correspondence
Nobuyuki Katsuma, Environmental Research Center Co. Ltd., 3-1, Hanare, Tsukuba, Ibaraki 305-0857, Japan E-mail: n_katsuma@erc-net.com Received 1 September 2014 Accepted 2 December 2014
Entomological Research Bulletin 31(1): 18-21 (2015)
2015 The Authors. Entomological Research Bulletin 2015 The Entomological Society of Korea
ages oval in dorsal and lateral view, with many long and short setae. Upper part of tergum X single, clavate, slightly shorter than lower part of tergum X, deeply divided, each tip with 3-4 short spines-like setae. Lower part of tergum X tall, hood-like, deeply excised into two vertical lobes, each lobe with broad and round apex in lateral view. Inferior append- ages triangular in lateral view, each with many setae, internal margin concaved, with short 3-4 spines; a pair of tiny lobe, clavate, arising from internal base of the inferior appendages,
each tip with few setae. Phallobase short, bulbous, ventro- lateral margin dark pigmented; phallicata almost as long as phallobase, with two pairs of wedge-like spines ventrally, with pair of short paramere spines dorsally; phallotremal sclerite U-shaped in dorsal view.
Female genitalia(Figs 1G-I). Segment IX rectangular in lateral view, fused with segment X, with triangular lobe pos- terodorsally; triangular lobe usually with paired papillae, but shape of papillae variable, sometimes asymmetrical in some
A New Species of the Genus Adicella
Figure 1.Adicella odamiyamensissp. nov. A, B. right wings, A: male, B: female; C-F, male genitalia, C: dorsal, D: ventral, E: lateral; F: phallus, lateral; G-I, female genitalia, G: dorsal, Ga: variation of tergum IX; H: ventral; I: lateral.
A
B
F
C D E
G H I
Ga
1 mm
0.5 mm