PAULO LEONARDO ARAÚJO DE GÓIS MORAIS
PROJEÇÃO RETINIANA, CARACTERIZAÇÃO CITOARQUITETÔNICA E
NEUROQUÍMICA DA ZONA INCERTA DO MOCÓ (Kerodon rupestris)
Dissertação de Mestrado submetida ao Programa de Pós-graduação em Psicobiologia da Universidade Federal do Rio Grande do Norte.
PAULO LEONARDO ARAÚJO DE GÓIS MORAIS
PROJEÇÃO RETINIANA, CARACTERIZAÇÃO CITOARQUITETÔNICA E NEUROQUÍMICA DA ZONA INCERTA DO MOCÓ (Kerodon rupestris)
Dissertação de Mestrado submetida ao Programa de Pós-graduação em Psicobiologia da Universidade Federal do Rio Grande do Norte.
Orientador: Prof. Dr. Expedito Silva do Nascimento Júnior
TÍTULO: PROJEÇÃO RETINIANA, CARACTERIZAÇÃO CITOARQUITETÔNICA E NEUROQUÍMICA DA ZONA INCERTA DO MOCÓ (Kerodon rupestris)
AUTOR: PAULO LEONARDO ARAÚJO DE GÓIS MORAIS
DATA DA DEFESA: 27/03/2014 EXAMINADORES:
__________________________________________ Prof. Dr. Fausto Pierdoná Guzen
__________________________________________ Prof. Dr. Jeferson de Souza Cavalcante
AGRADECIMENTOS
Agradeço primeiramente a minha família, Ana Maria, José Orlando e Paula Giovanna, base de tudo, que é diretamente responsável por tudo de bom que acontece comigo e que sei que posso contar quando as coisas ruins acontecem. Amo vocês! Agradecimento especial para o Prof Expedito Jr, meu orientador e grande líder, que fez tudo e mais um pouco para me ajudar a trabalhar.
Aos professores Judney, Jeferson, Miriam, Celcimar e Ruthnaldo, pela compahia, duvidas esclarecidas e sugestões, tantos para os experimentos e trabalhos quanto para outras questões cotidianas.
A Todos os alunos que passaram pelo Labneuro, pelos momentos alegres, ajuda nos experimentos, compahia nos diversos lugares, desde os almoços de confraternização até nos congressos e simpósios da vida, em especial para Karen, André, Melquisedec, Mariana, Nelyane, Fladjane, Helder, Wilqui, Renata, Nayra, Felipe, Joacil, Twyla, Rovena, e um agradecimento especial a Dona Regina, responsável pela manutenção da lei, da ordem e do funcionamento geral do Laboratorio.
Aos meus grandes amigos da biologia e agregados, Rodolfo, Rafael, George, Ivon Edipo, Huguinho, Guido, Cleanto, Stanlley, Fernando, Daniel, Bozena, Alf, Marilia, Vanessa, Aline, Helton e mais uma galera grande, pelos momentos de alegria, conhecimento, e angustias compartilhados desde o período de graduação.
A galera do ABC Leozinho, Joao Paulo, Romario, Oreia, Duble, Michel, Idamilton, Marquinhos, Bia, Junior e Nessah, excelentes compahias em jogos, viagens e em vários outros momentos unidos por uma paixão em comum pelo ABC.FC.
E o que dizer das galeras dos cursos de Geofisica 2013.1 e Fisioterapia 2013.1 e .2, que convivi pouco tempo, mas já os considero pacas? Amizades surgidas a pouco tempo pela loucura de querer cursar outra graduação durante o mestrado, mas também importantes para conhecer outros ares e áreas do conhecimento.
LISTA DE ABREVIATURAS 3v – terceiro ventrículo
5-HT – Serotonina CB – Calbindina ci – Capsula interna CR – Calretinina CS – Colículo superior DA - Dopamina f - Fornix
FIG – folheto intergeniculado GABA -ácido gama aminobutírico GFAP - Proteína acídica fibrilar glial GLD – Núcleo Geniculado lateral dorsal GLV – Núcleo Geniculado lateral ventral HRP – horseradish peroxidase
IR- - Imunoreativo
MD - núcleo médio dorsal do tálamo ml – lemnisco medial
mt – tracto mamilotalâmico NO - óxido nítrico
NPY - Neuropeptídeo Y NSO – núcleo supraóptico NSQ – Núcleo supraquiasmático PV – Parvalbumina
PVT – núcleo paraventricular do tálamo Rt – núcleo reticular do tálamo
rts – radiação talâmica superior TF – tampão fosfato
TH – Tirosina hidroxilase
VIP - Polipeptídeo intestinal vasoativo VP – Vasopressina
VM - núcleo ventral póstero medial ZI – Zona Incerta
LISTA DE FIGURAS
Tabela 1: Especificação das substâncias, bem como seus fabricantes e diluições, utilizadas nos procedimentos de imunohistoquímica.
Figura 1: Área de distribuição do mocó. Figura 2: mocó (Kerodon rupestris).
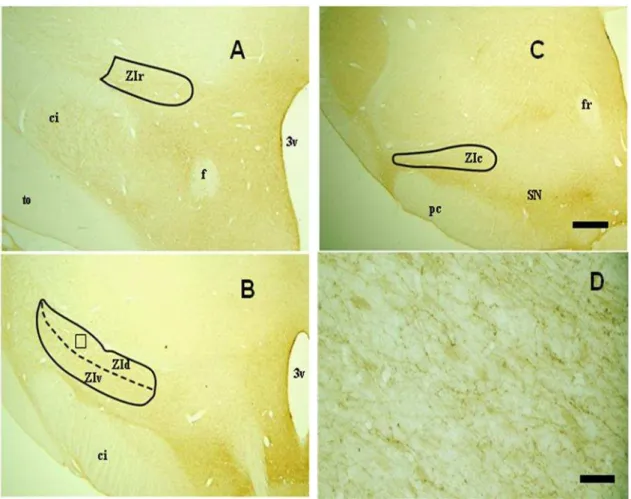
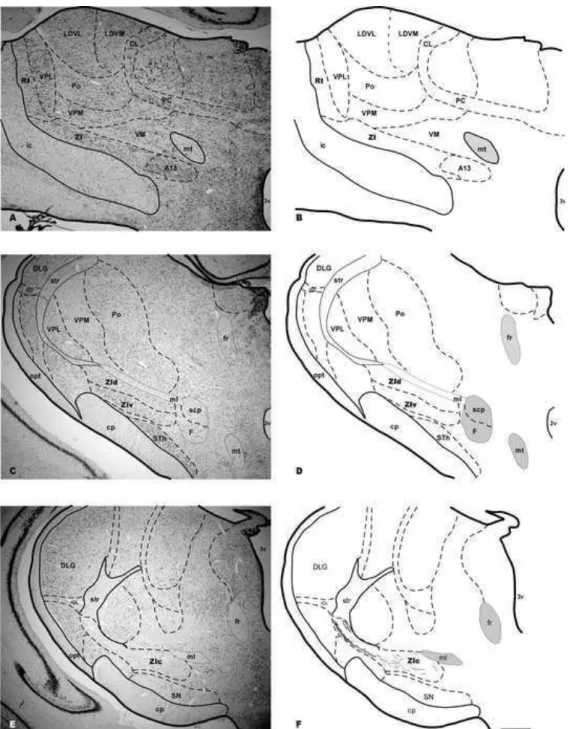
Figura 3: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó coradas pelo método de Nissl nas subdivisões da ZI.
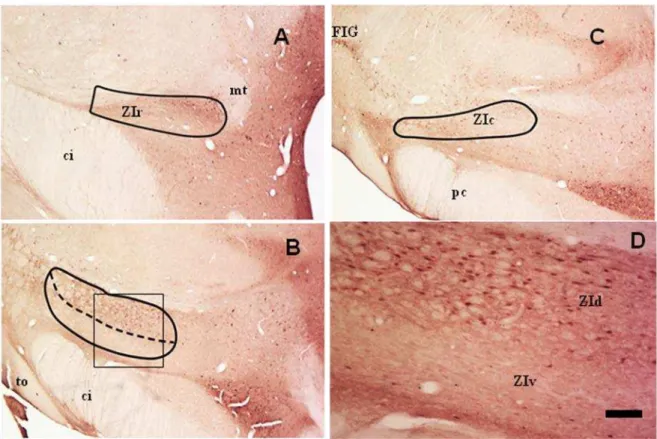
Figura 4: Em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra CTb, evidenciando a projeção retiniana para a ZIc.
Figura 5: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra TH.
Figura 6: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra NOS.
Figura 7: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra 5-HT.
Figura 8: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra GFAP.
Figura 9: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra CB.
Figura 10: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra CR.
Figura 11: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó, mostrando a imunorreatividade contra PV.
SUMÁRIO
RESUMO 10
ABSTRACT 11
1. INTRODUÇÃO 12
1.1 Zona Incerta (ZI) 12
1.2. Subdivisões da ZI 12
1.3 Hodologia da ZI 13
1.4 Aspectos Funcionais da ZI 14
1.5 Projeções retinianas 16
1.6 Projeções retinianas para núcleos classicamente não visuais 17
1.7 Sujeito experimental 18
1.8 Justificativa 20
2.OBJETIVOS 21
2.1 Objetivos gerais 21
2.2 Objetivos específicos 21
3. METODOLOGIA 22
3.1 Anestesia 22
3.2 Injeção intra ocular 22
3.3 Perfusão 23
3.4 Remoção dos Encéfalos 23
3.5 Microtomia 23
3.6 Método de Nissl 24
3.7 Imunohistoquímica 24
3.8 Análise das imagens 26
4. RESULTADOS 27
4.1 Citoarquitetura Nissl 27
4.2 Projeção retiniana 29
TH 31
NOS 32
5-HT 33
GFAP 34
CB 35
CR 36
PV 37
5 DISCUSSÃO 39
5.1 Citoarquitetura 39
5.2 Projeção retiniana 41
5.3 Neuroquímica 44
TH 44
NOS 46
5-HT 47
GFAP 48
CB 48
CR 49
PV 50
5.4 Considerações do funcionamento global da ZI 51
6. CONCLUSÕES 53
7. REFERÊNCIAS BIBLIOGRÁFICAS 54
11 RESUMO
A Zona Incerta (ZI) é um grupamento neuronal embriologicamente derivado do tálamo ventral, em continuidade com o núcleo reticular do tálamo. Diversos estudos com traçadores retrógrados e anterógrados revelaram a conexão da ZI com diversas estruturas do sistema nervoso central. Dados moleculares e citoquímicos revelaram que a ZI é um dos grupamentos neuronais com maior diversidade neuroquímica e citoarquitetônica do diencéfalo, e estudos hodológicos e neuroquímicos permitiram considerar o envolvimento da ZI em diversas funções, as quais se destacam a nocicepção, atenção, estado de alerta, controle e manutenção da postura e controle da atividade visceral. Este trabalho tem por objetivo caracterizar a citoarquitetura e o conteúdo neuroquímico da ZI do mocó (Kerodon rupestris), bem como a aferência óptica presente neste núcleo nesta espécie. A técnica de Nissl é eficiente para a delimitação e caracterização citoarquitetônica da ZI do mocó; A ZIc recebe projeção da retina contralateral, apresentando fibras Classe II ou modulator, sugerindo um caráter modulatório da informação fótica; A ZI do mocó, assim como em outros roedores e primatas, é caracterizada por uma complexa rede neuroquímica, sobretudo na porção medial da ZIr, onde encontramos imunorreatividade de todas as substâncias neuroativas investigadas, além de que A IR-NOS, GFAP e CR auxiliaram a delimitação da ZI no nível médio em ZId e ZIv. Contudo, somente fibras IR 5-HT estão presentes em todas as subdivisões da ZI. Esses dados demonstram a grande riqueza neuroquímica da ZI do mocó, auxiliando para explicar o envolvimento em um amplo repertorio funcional.
Palavras chave: Zona Incerta, neuroquímica, citoarquitetura, projeção retiniana,
12 ABSTRACT
The Zona Incerta (ZI) is embryologically derived from the ventral thalamus, in continuity with the reticular nucleus of the thalamus. Studies usingneural tracers technics have allowed identify a complex connectional map including the ZI. Futhermore, cytochemical, molecular and functional data have shown abundant variability in the neurochemical contend in the ZI, as well as,the involvement of the ZI in the modulation of nociception, attention, alertness, control and maintenance of posture and control of visceral activity. This work aims to characterize the cytoarchitecture, neurochemical content of the ZI in the rock cavy (Kerodon rupestris), and a direct retinal-ZI pathway present in this species. The Nissl staining is effective for the delineation and characterization of ZI citoarchitecture. ZIc receives a contralateral retinal projection showing varicosities, suggesting a modulatory character of photic information. The ZI in the rock cavy, as in others rodents and primates, is characterized by a complex neurochemical signature. The ZI neurochemistry presents great diversity, especially in the medial portion of ZIr, where we have found immunoreactivity of all neuroactive substances investigated, and that NOS-IR, GFAP and CR helped the delimitation of middle ZI in ZId and ZIv. Nevertheless, just 5-HT-IR fibers are present in all subdivisions of the ZI. These data demonstrate the great wealth of the neurochemistry of rock cavy’s ZI and a direct retinal modulation in the ZI, helping to explain it’s broad functional repertory.
Keywords: Zona Incerta, neurochemistry, cytoarchitecture, retinal projection,
13
1. INTRODUÇÃO
1.1 Zona Incerta (ZI)
A ZI é um grupamento neuronal embriologicamente derivado do tálamo ventral (Jones, 2007), em continuidade com o núcleo reticular do tálamo (rt) (Kolmac e Mitrofanis, 1999). Desde sua primeira descrição, feita por Forel em 1877, diversos estudos com traçadores retrógrados e anterógrados revelaram a conexão da ZI com diversas estruturas do sistema nervoso central. Dados moleculares e citoquímicos revelaram que a ZI é um dos grupamentos neuronais com maior diversidade neuroquímica e citoarquitetônica do diencéfalo (Mitrofanis, 2005; Nicolelis et al, 1995). Esses estudos hodológicos e neuroquímicos permitiram considerar o envolvimento da ZI em diversas funções, as quais se destacam a atenção, estado de alerta, controle e manutenção da postura e controle da atividade visceral (Mitrofanis, 2005) e nocicepção (Petronilho et al, 2012).
1.2. Subdivisões da ZI
14
1.3 Hodologia da ZI
As principais fontes aferentes para a ZI são: retina, núcleo central do complexo amigdalóide, núcleo sensitivo do trigêmio, área hipotalâmica posterior e lateral, núcleo ventromedial do hipotálamo, rt, núcleo talâmico posterior, núcleo geniculado lateral ventral (GLV), complexo ventrobasal, área pretectal, núcleo peripeduncular, as camadas profundas e intermédia do colículo superior (CS), colículo inferior, núcleo coclear, substância cinzenta periaquedutal, complexo parabraquial, pars reticulata da substância negra (SN), áreas perirubrais, núcleo interpósito do cerebelo, área tegmentar ventral, núcleo reticular pontino, núcleo grácil, núcleo cuneiforme, núcleo intersticial de Cajal, núcleo do oculomotor, núcleo da coluna dorsal, núcleo do trato espinal do trigêmio, núcleo supraquiasmático (NQS), folheto intergeniculado (FIG) e núcleo tegmental pedunculopontino. Várias áreas corticais também se projetam para a ZI: córtex cingulado, motor primário, pré frontal, sensório motor primário e secundário, occipital, neocórtex temporal, piriforme, tubérculo olfatório e núcleo septal medial (Roger e Cadusseau, 1985; Mitrofanis e Mikuletic, 1999; Mitrofanis, 2004; May et al, 1997; Shaw e Mitrofanis, 2001, Lechner et al, 1993; Barthó et al, 2002, Çavdar et al, 2006, Leak e Moore, 2001; Power et al, 2001, Mitrofanis, 2005; Mitrofanis et al, 2004; Gaillard et al, 2013; Morin, 2013).
15
1.4 Aspectos Funcionais da ZI
Os vários estudos sobre a neuroquímica e as conexões auxiliaram os pesquisadores a propor prováveis funções para ZI, e a partir disso as pesquisas mais recentes detalham sobre como a ZI atua no processamento de diversas informações, através de estudos farmacológicos, eletrofisiológicos e de lesão. Mitrofanis (2005) faz uma revisão geral sobre os estudos sobre a ZI, e aponta quatro funções gerais desempenhadas pela ZI: Controle da atividade visceral, manutenção da postura e locomoção, Atenção e excitação. Outros estudos ressaltam a importância da ZI no processamento da informação nociceptiva.
O envolvimento da ZI em funções como a ingestão alimentar e hídrica, comportamento sexual e mudanças na função cardiovascular exemplificam bem a sua importância sobre a atividade visceral (Mitrofanis, 2005). Estudos de lesão, bem como injeções de lidocaína na ZI, induzem um aumento de ingestão de comida e água, sugerindo que a ZI atua em mecanismos de inibição dos sinais de saciedade (Tonelli e Chiaraviglio, 1993). Em (1995), esses mesmos autores também verificaram que a ativação do receptor D2 de dopamina (DA) na ZI inibe a ingestão alimentar e hídrica. A ZI é alvo de diversos sistemas neuronais, entre eles GABA, DA serotonina (5-HT), orexina, hormônio de concentração de melanina, que estão envolvidos com a liberação de gonadotrofina (Siddiqui et al, 2004). Também há relatos que a injeção de L-glutamato na ZI diminui a frequência cardíaca e a pressão arterial (Spencer et al, 1988). Estudos eletrofisiológicos e farmacológicos, estimulando ou inibindo esses sistemas, detectaram que os neurônios da ZI apresentam características intrínsecas de membrana que geram disparos rítmicos espontâneos (Trageser e Keller, 2006). A ZI se projeta para o bulbo olfatório de ratos, estrutura fundamental para a percepção do ambiente dos roedores em geral. Especula-se que a ZI, que recebe projeção do rt, serve como um intermediário que modula a atividade dos neurônios do bulbo olfatório, alterando seu padrão de atividade elétrica (Uemura-Sumi et al, 1985).
16 com funções motoras, como o trabalho de Supko et al (1991), que verificaram que a injeção unilateral de L-glutamato age especificamente em receptores AMPA e cainato, e provocam aumento de mobilidade, seguido de mudança postural. Em contrapartida, a ativação de receptores de GABA na ZI diminui a atividade locomotora (Wardas et al, 1988). A ZI também apresenta conexão com o CS, atuando na modulação dos movimentos dos olhos e da cabeça (Moschovakis, 1996). Esta modulação ocorre através de células GABAérgicas localizadas na porção ventral da ZI, corroborado por estudos eletrofisiológicos, que detectaram que a inibição da células da ZI sobre as células coliculares, e farmacológicos, onde a administração de um agonista (mucimol) ou de um antagonista (bicuculina) provocam movimentos irregulares (“tilts”) nos olhos e na cabeça (Murer e Pazo, 1993). Outro importante estudo demonstrou evidências de que a ZI é organizada somatotopicamente (Nicolelis et al, 1992), e outros que a ZI possui uma conexão com os núcleos da base, núcleos fundamentais para a execução de movimentos voluntários (Heise e Mitrofanis, 2004). Ainda no que tange a sensibilidade, verificou-se que a ZI apresenta ativação neuronal rítmica, e que esse padrão de disparo é muito importante para a sensibilidade, regulando, por exemplo, a atividade dos músculos das vibrissas em ratos da linhagem Long Evans, apresentando queda significativa na taxa de disparo espontâneo, alterações estas também documentadas em células corticais, em indivíduos com lesão unilateral da ZI (Shaw et al, 2013).
17
1.5 Projeções retinianas
No sistema nervoso central dos mamíferos, a entrada de luz proveniente da retina alcança dois sistemas funcionais distintos: o sistema formador de imagem, responsável pela formação das imagens do ambiente, e o sistema não formador de imagem, responsável pela integração visuomotorae da geração e regulação dos ritmos biológicos. Outro grupo de fibras oriundas da retina, projetam-se para áreas classicamente não-visuais, envolvidas com diversas outras funções e dentre elas a ZI, região de interesse neste estudo.
O sistema formador de imagem, classicamente denominado de sistema visual primário, é composto por vários conjuntos neuronais localizados no diencéfalo e mesencéfalo são responsáveis pela formação de imagens. O núcleo geniculado lateral dorsal (GLD) é principal estação de retransmissão do sinal visual (Jones, 2007). Já o GLV está envolvido em funções como estimulação visual e visuomotora (Jones, 2007). O complexo pré tectal um conjunto de núcleos localizados na transição entre o mesencéfalo e o diencéfalo, sendo responsável pelo controle do reflexo pupilar (Kaas e Huerta, 1988). Este complexo foi alvo de estudos no macaco-de-cheiro (Saimiri sciureus), que o dividiu em cinco núcleos: Pré tectal anterior, medial e posterior, núcleo o trato óptico e núcleo olivar pré tectal, todos eles recebendo projeção da retina (Hutchins e Weber, 1985). Outro componente do sistema formador de imagem é o CS. Este apresenta uma estrutura laminar, geralmente contendo sete camadas, sendo as camadas superficiais as que recebem aferência retiniana. O CS está envolvido com integração vísuomotora e o controle dos movimentos dos olhos e da cabeça (Kaas e Huerta, 1988).
18 temporização circadiana é o FIG, que se trata de uma lâmina celular intercalada entre o GLD e o GLV, que recebe projeção bilateral da retina. (Moore e Lenn, 1972; Moore, 1973; Morin e Allen, 2006; Cavalcante et al, 2006; Morin, 2013).
1.6 Projeções retinianas para núcleos classicamente não visuais
Com o desenvolvimento de técnicas utilizando traçadores mais sensíveis, os pesquisadores começaram a detectar projeções retinianas para áreas classicamente não visuais. Diversas áreas talâmicas consideradas não visuais foram descritas como retino-recipientes, como os núcleos anterodorsal e lateral posterior (Kudo et al., 1988; Youngstrom et al., 1991; Martinet et al., 1992; Fite e Janusonis, 2001). No tronco encefálico, projeções retinianas foram detectadas no núcleo dorsal da rafe (Foote et al., 1978; Shen e Semba, 1994; Fite et al., 1999; Fite e Janusonis, 2001), no núcleo parabraquial (Fite e Janusonis, 2002; Engelberth et al., 2008), como também no CS (Itaya e Van Hoesen, 1982). Projeções retinianas também foram descritas nos núcleos intralaminares e da linha média (Cavalcante et al.. 2005), no núcleo habenular lateral (Qu et al., 1996.). O tubérculo olfatório, o complexo amigdalóide no prosencéfalo basal e o córtex piriforme são outros exemplos de terminais retinianos não envolvidos com o sistema formador de imagem (Cooper et al, 1993; Martinet et al, 1992; Mick et al, 1993; Herbin et al, 1994; Elliot et al, 1995).
19
1.7 Sujeito experimental
O mocó (Kerodon rupestris) é um roedor pertencente à família caviidae, nativo da caatinga do nordeste brasileiro, também com presença registrada no norte do estado de Minas Gerais (Cabrera, 1961) e com introdução bem sucedida na ilha de Fernando de Noronha em 1967 (Oren, 1984). Animais dessa família são dotados de grande capacidade de adaptação aos mais diversos tipos de ambiente, entretanto o mocó exibe uma preferência por habitar regiões áridas de pedreiras com várias fendas e rachaduras, onde costuma ficar alojado. Considerando a classificação filogenética baseada em aspectos morfológicos e comportamentais, o gênero kerodon, a que pertence ao mocó, é agrupado monofileticamente com os gêneros Cavia, Galea e Microcavia, compondo a família Caviidae (Cabrera, 1961; Lacher, 1981), entretanto, considerando aspectos genéticos, o gênero kerodon é incluído na família hydrochaeridae, a mesma família da capivara, pertencente ao gênero Hydrochaeris (Rowe e Honeycutt, 2002).
Figura 1: Área de distribuição do mocó
20 O mocó apresenta coloração cinza clara, castanho ferruginoso na região caudal e no dorso, um pouco acastanhada nas patas e branco na região cervical, podendo atingir 30cm de altura e pesar 1kg quando adulto (Moojen, 1952; Carvalho, 1969). As patas dotadas de coxins calosos e unhas rígidas lhe confere ótima habilidade saltatória e para escalar, apesar de não possuir garras e cauda, adaptações comuns em animais com essas habilidades. Tem olfato e audição bastante aguçados, o que lhes permite detectar a presença de seus predadores a uma longa distância (Carvalho, 1969).Sua alimentação composta principalmente de cascas de árvores, com preferência para o mufumbo (Cobretum leprosum), Parreira Brava (Cissampelos pareira) e a faveleira (Cnidoscolus phyllacanthus). Na ausência dessas árvores, o mocó alimenta-se de gramíneas em geral (Lacher, 1981; Carvalho, 1969). O mocó tem um período médio de gestação de 65 dias, com número médio de 1,2 filhotes por gestação. Apesar da baixa quantidade de filhotes gerada por gestação, a reprodução ocorre durante o ano todo, exceto durante os meses de abril a junho, e o baixo período gestacional garantem uma elevada produção de filhotes durante o ano (Lacher, 1981).
21
1.8 Justificativa
Considerando a importância da ZI no controle e modulação de diversas funções do organismo, bem como de sua atual importância clínica em relação à alternativas de tratamentos para doenças neurodegenerativas, como esclerose múltipla e mal de Parkinson, é de fundamental conhecer e esclarecer características neuroanatômicas e neuroquímicas da ZI. Também vale destacar que, mesmo com os avanços dos estudos, pouco se sabe sobre como a ZI interage com tantas regiões do sistema nervoso, que desempenham funções tão distintas, indo desde locomoção até controle hormonal.
22
2. OBJETIVOS
2.1Objetivos gerais
Caracterizar a citoarquitetura e o conteúdo neuroquímico da ZI do mocó (Kerodon rupestris), bem como a aferência óptica presente neste núcleo nesta espécie.
2.2 Objetivos específicos
• Definir citoarquitetonicamente as subdivisões da ZI do mocó.
• Mapear e descrever os aspectos morfológicos das projeções retinianas marcadas positivamente para CTb na ZI do mocó.
23
3. METODOLOGIA
3.1 Anestesia
Os 4 animais utilizados neste trabalho foram anestesiados conforme o seguinte protocolo: inicialmente foi utilizado como medicação pré-anestésica o sulfato de atropina na dosagem de 0,04 mg/Kg, via subcutânea, e 2 mg/Kg de Tramadol, via intramuscular; passados 15 minutos, os animais receberam ketamina e xilazina, administradas após serem misturados na mesma seringa, na dosagem de 20 mg/Kg e 2mg/Kg respectivamente. O plano anestésico, bem como sua manutenção, foi realizado por meio de máscara com oxigênio e isoflurano.
3.2 Injeção intra ocular
Após a completa anestesia de um deles, este foi submetido a uma injeção intraocular unilateral de 70μl de uma solução aquosa da CTb (List Biological Laboratories, Inc., Campbell, CA)a 5%, contendo dimetilsulfóxido (DMSO) a 10% com a finalidade de aumentar a permeabilidade e, consequentemente, melhorar a captação do traçador. A injeção ocorreu no olho esquerdo, conforme protocolo do laboratório, utilizando-se uma agulha calibre 30 (8,00mm X 0,3mm), lentamente, sob pressão, com auxílio de uma microbomba propulsora, a qual impulsiona a solução em um fluxo de 1μl/minuto. Essa agulha foi introduzida na junção esclera-corneal, atingindo o corpo vítreo em um ângulo de aproximadamente 45º. Após o término do fluxo da solução, a agulha foi mantida por mais 15 minutos, visando diminuir um possível refluxo da solução.
24
3.3 Perfusão
Atingido o plano anestésico, cada animal foi submetido à perfusão transcardíaca, que compreende os seguintes passos:
1 –Posicionamento do animal em decúbito dorsal sobre tela de arame e sob ponto de água.
2 –Toracotomia, com incisão de pele, músculos e arco costal, sendo estes removidos em bloco, para exposição do coração.
3 –Cardiopunção no ventrículo esquerdo, utilizando uma agulha de dimensões 17mm x 1,5 mm, a qual é direcionada para o cone arterioso, seguindo-se uma incisão no átrio direito. A agulha foi conectada a uma bomba peristáltica (Cole-Parmer), passando-se 300ml de solução salina a 0,9% em tampão fosfato 0,1M, pH 7,4 com heparina (Parinex, Hipolabor, 2ml/1000 ml de solução salina) durante um tempo estimado de seis minutos.
3.4 Remoção dos Encéfalos
Finalizada a etapa de perfusão, os animais foram posicionados no aparelho estereotáxico para roedores. Depois de se fazer uma incisão longitudinal na pele e rebatê-la lateralmente, fez-se a limpeza da superfície óssea, facilitando a visualização do bregma e do lambda, os quais ficaram nivelados na mesma altura dorsoventral, ajustando-se a barra dos incisivos. Em seguida, os ossos da calota craniana foi removido com o uso de broca e trocater, expondo-se o encéfalo. Estes foram retirados delicadamente para evitar danos, preservando os olhos e nervos ópticos (uma vez que estes animais foram também utilizados em outra pesquisa). Logo após esta etapa, os três blocos foram armazenados em uma solução contendo sacarose a 30% em tampão fosfato (TF) 0,1M, pH 7,4, a 4 ºC, até serem submetidos à microtomia.
3.5 Microtomia
25 maneira cíclica e alcoóis cada. Os cortes de um compartimento foram imediatamente montados em lâminas de vidro gelatinizadas e submetidas à coloração pelo método de Nissl para permitir uma melhor demarcação das estruturas. Os cortes dos demais compartimentos foram transferidos para solução anticongelante e conservados a -20 ºC para utilização posterior em procedimentos de imunohistoquímica.
3.6 Método de Nissl
A coloração pelo método de Nissl é constituída por uma série de etapas que se iniciam com a desidratação do tecido passando-o em concentrações crescentes de alcoóis etílicos (70% - 1 vez, por 1h, 95% - 2 vezes, 3 minutos cada, 100% - 2 vezes, 3 minutos cada). Posteriormente são deslipidificados em dois recipientes com xilol, por 3 e 30 minutos, nessa ordem. Na seqüência, o tecido é reidratado em concentrações decrescentes de 25lcoóis (mesmo processo anterior, mas no sentido inverso), submergido em água destilada por 2 minutos, colocado 1 minuto na solução de thionina a 0,25%. Por fim os cortes foram rapidamente submergidos e emergidos por 1 minuto em água destilada e novamente desidratados e deslipidificados como descrito anteriormente, sendo acrescentada apenas uma etapa (antes da desidratação em álcool 100%). Os cortes foram deslipidificados em xilol em dois recipientes (2 e 4 minutos, respectivamente) e finalmente cobertos com lamínula utilizando como meio demontagem o ERV-Mount.
3.7 Imunohistoquímica
Para iniciar a imunohistoquímica, foi feito um pré-tratamento (incubação em peróxido de hidrogênio (H2O2) a 0,3% em TF 0,1M, pH 7,4) por 20 minutos, com a finalidade de abolir artefatos causados pela liberação de peroxidases endógenas. No início, entre as soluções e ao final desta fase, os cortes foram submetidos a 5 lavagens de 5 minutos cada em TF 0,1 M, pH 7,4.
26 Tabela 1: Especificação das substâncias, bem como seus fabricantes e diluições, utilizadas nos procedimentos de imunohistoquímica.
Antígeno Anticorpo primário Anticorpo secundário Soro normal/ BSA
CTb Cabra [1:5000] List Biological
Asno [1:1000]
Sigma
Asno [1:50] TH Camundongo [1:10000] Sigma Cabra [1:1000] Jackson BSA
NOS Coelho [1:1000] Sigma Cabra [1:1000] Jackson BSA CB Coelho [1:1000] Sigma Cabra [1:1000] Jackson BSA CR Coelho [1:1000] Sigma Cabra [1:1000] Jackson BSA PV Coelho [1:5000] Sigma Cabra [1:1000] Jackson BSA 5-HT Coelho [1:5000] Sigma Jumento [1:1000] Jackson BSA GFAP Camundongo [1:1000] Sigma Cabra [1:1000] Jackson BSA
Em seguida, os cortes passaram por 5 lavagens em TF 0,1 M em agitador orbital e depois colocados na solução do complexo avidina-biotina (Protocolo ABC, Kit elite da Vector), numa diluição de 1:100 em Triton X-100 a 0,4%, contendo NaCl, por 90 minutos à temperatura ambiente, sob agitação lenta, em rotor. Terminada esta fase, as secções foram novamente submetidas a 5 lavagens em TF 0,1 M em agitador orbital.
27
3.8 Análise das imagens
As secções do mesencéfalo, coradas pelo método de Nissl e/ou submetidas à imunohistoquímica foram examinadas ao microscópio óptico (Olympus BX41) em campo claro. Imagens digitais foram obtidas de secções representativas usando uma videocâmara digital (Nikon DXM1200) e Motican 5.0 M camera. As imagens foram analisadas, corrigidas minimamente para brilho e contraste e os desenhos esquemáticos foram montados no software Canvas X® (ACD Systems, Victoria, BritishColumbia, Canada). Os resultados foram documentados em fotomicrografias e esquemas construídos a partir das mesmas no Power Point.
28
4. RESULTADOS
4.1 Citoarquitetura Nissl
30
4.2 Projeção retiniana
32
4.3 Neuroquímica
TH
Na ZI do mocó, células e fibras IR-TH foram detectadas na ZIr, sendo possível visualizar poucas células marcadas (Figura 5A). Estes neurônios apresentam formato triangular e irregular, com axônios de pequeno tamanho (Figura 5B).
33
NOS
A ZI do mocó apresenta neurônios IR-NOS em todas as subdivisões da ZI, com formato ovoide e fusiforme, com pequenas varicosidades (Figura 6D). Na ZIr são visualizadas células IR-NOS concentradas na porção medial (Figura 6A). A ZIv possui mais células marcadas em comparação com a ZId, localizados principalmente na região lateral, próximo ao rt (Figura 6B). Na ZIc, neurônios IR-NOS concentram-se na região lateral, com poucas células presentes na região medial (Figura 6C).
34
5-HT
Na ZI do mocó identificamos a marcação de fibras e varicosidades IR-5-HT em todas as subdivisões (Figura 7 A-C). Na ZId e ZIv, uma maior quantidade de fibras e varicosidades foram detectadas (D).
35
GFAP
Células IR-GFAP foram encontradas na ZIr (Figura 8A), ZId (Figura 8B) e ZIc (Figura 8C), enquanto que na ZIv a marcação foi negativa (Figura 8B). Células estreladas com pequenas radiações estão presentes nas 3 regiões marcadas, mas na ZIr também detectamos células maiores, de formato ovoide (Figura 8D).
36
CB
Células IR-CB foram detectadas na ZIr (Figura 9A), apresentando formato alongado e irregular (Figura 9B). Na ZId, ocorre uma grande quantidade de neurópila, enquanto a ZIv há uma menor quantidade (Figura 9B). Na ZIc, ocorre uma concentração de neurópila na porção lateral, mesma região onde encontra-se a projeção retiniana (Figura 9C).
37
CR
Neurônios IR-CR apresentam formato ovoide, localizados na ZIr, ZId e ZIc. Na ZIr, ocorre uma concentração desses neurônios próximos ao mt (Figura 10A). Na ZId detectamos grande marcação por toda sua extensão. Em contrapartida, a ZIv não apresenta nenhuma imunorreatividade para CR (Figura 10B e D). Na ZIc, os neurônios estão esparsamente distribuídos em toda sua extensão (Figura 10C).
38
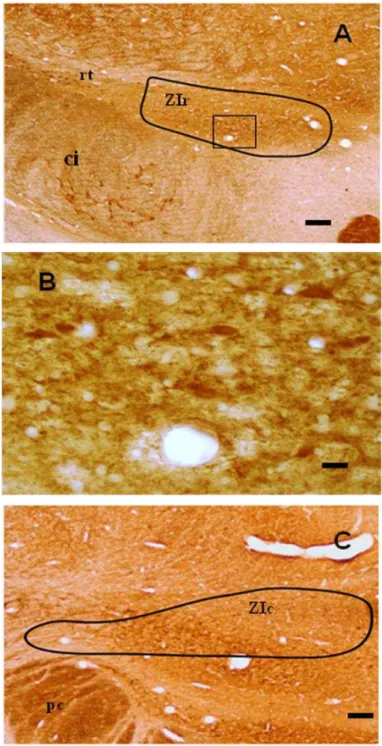
PV
Células e neurópila IR-PV foram detectadas na região medial da ZIr (Figura 11A) e intermédia da ZIc (Figura 11C), apresentando formato ovoide e irregular (Figura 11B).
39 Tabela 2: Resumo esquemático da distribuição das substâncias neuroativas nas subdivisões da ZI do mocó.
Substância/Divisão ZIr ZId ZIv ZIc
Projeção da retina(CTb) - - - +
TH + - - -
NOS + - + +
5-HT (fibras) + + + +
GFAP + - + -
CB + - - -
CR + + - +
40
5. DISCUSSÃO
5.1 Citoarquitetura
A ZI do mocó está dividida em 4 porções, de acordo com a análise citoarquitetônica observada em nossos resultados. Devido a grande diversidade celular da ZI e algumas dificuldades em delimitar claramente suas subdivisões, o número destas varia de autor para autor: de 2 a 6 em ratos, entretanto, os estudos mais recentes consideram apenas 4, de 2 a 4 em macacos e superior a 5 em gatos (Mitrofanis, 2005).
Em rato, a ZI é classicamente subdividida em quatro regiões: rostral, dorsal, ventral e caudal. Na região rostral corre uma predominância de células fusiformes, com raras células ovoides. A região dorsal possui células ovoides com tamanho médio, enquanto que na ventral as células são densamente agrupadas e com formato fusiforme. Por fim, a região caudal apresenta células de tamanho pequeno a médio com formatos fusiforme, ovoide e multipolar, podendo também apresentar células multipolares com grande tamanho (Kolmac e Mitrofanis, 1999; Power et al, 2001) Em um estudo que focou somente as porções dorsal e ventral, foram detectados neurônios com morfologia bipolar e fusiforme, com grandes dendritos (Trageser e Keller, 2006).
Na ZI do cachorro, foram delimitadas 5 regiões, rostral, dorsal, ventral, caudal, e uma extensão do nível rostral, denominada porção dorsolateral. Esta porção dorsolateral é típica em carnívoros, apesar de não ter sido identificada no furão. Os neurônios têm diversos formatos e tamanhos em todas as porções, mas com prevalência de neurônios de tamanho intermediário e formato fusiforme na porção rostral, formato ovoide de pequeno tamanho na porção dorsal, formato triangular de tamanho intermediário na porção ventral, e nas porções dorsolateral e caudal não houve uma prevalência de um tamanho ou formato específico (Gorbachevskaya e Chivileva, 2008).
42
5.2 Projeção retiniana
Os estudos sobre projeções retinianas fornecem uma melhor definição do trajeto axonal, além de possibilitar uma visualização detalhada dos terminais, algo fundamental para compreensão do funcionamento de diversos sistemas que utilizam a informação visual (Rouiller e Welker, 2000). A CTb, a subunidade B da toxina colérica, é um traçador axonal, altamente sensível, que atual tanto retrogradamente como anterogradamente (Angelucci et al, 1996; Gaillard et al, 2013). Sua utilização como traçador das vias retinianas tem sido amplamente empregada em diversos animais, como pombo (Schimizu et al, 1994), primatas (Nakagawa et al, 1998; Costa et al, 1999; Mitrofanis et al, 2004; Cavalcante et al, 2005; Engelberth et al, 2008; Lima et al, 2012) e roedores (Youngstrom, 1991; Abrahamson e Moore, 2001;Mitrofanis, 2005; Gaillard et al, 2013). No mocó, vários alvos retinianos já foram detectados utilizando CTb, como o NSQ e FIG (Nascimento Jr et al, 2010a), MD (Sousa, 2013; Nascimento Jr et al, 2010b), PVT (Nascimento Jr et al, 2008), trabalhos estes que comprovam a eficácia da CTb para estudos de traçados axonais nesta espécie. Em relação à ZI, detectamos a presença de fibras e terminais IR-CTb na porção lateral da ZIc.
43 achado, sugere-se uma participação da ZI no processamento visual, especificamente em relação aos movimentos dos olhos e da cabeça e de comportamentos sócio-sexuais (Power et al, 2001). No rato do Nilo (Arvicanthis niloticus), uma pequena projeção retiniana foi encontrada na porção dorsolateral da ZI (Gaillard et al, 2013). No sagui, fibras e terminais IR-CTb foram detectadas na região ventral da porção caudal da ZI, apresentando varicosidades (Lima, 2008).
44 No contexto das características sobre como as informações aferentes chegam em seus alvos, diversos estudos demonstraram que a morfologia das fibras axonais e dos terminais sinápticos apresentam diferenças importantes, e que essas diferenças também refletem em diferenças funcionais, sobre como as informações são transportadas e processadas. Fibras Classe I, ou Drivers, possuem terminais de grande tamanho, com alta capacidade de potenciais excitatórios pós-sinápticos (PEPS) e ativam receptores ionotrópicos, e no sistema visual, estas fibras são presentes em núcleos talâmicos específicos para a formação de imagens, como o GLD, enquanto que Fibras classe II, ou modulators, possuem terminais de pequeno tamanho, baixa capacidade de disparo de PEPS e que ativam receptores glutamatérgicos metabotrópicos (Shermam e Guillery, 2011). As fibras retinianas da ZI do mocó são de pequeno calibre, e mais de 60% de suas varicosidades encontram-se no eixo dessas fibras, ou seja, essa projeção retiniana é composta por fibras do tipo II, com predominância de varicosidades do tipo botões de passagem. Esses dados demonstram que a ZI recebe informação direta da retina, mas não está envolvida com a formação de imagem, usando esta informação para outras finalidades.
45
5.3 Neuroquímica
TH
A DA é uma substância resultante da ação da enzima DOPA-descarboxilase sobre a di-hidroxi-fenilalanina (DOPA), a qual deriva da tirosina por ação da TH (Marín et al., 2005; Chen et al., 2008). É importante considerar que, nos últimos anos, a expressão de TH em amostras encefálicas tem sido amplamente utilizada como marcador molecular de DA neuronal, embasado por evidências provenientes de estudos com ferramentas fisiológicas, hodológicas, farmacológicas, clínicas e de biologia molecular, permitem assegurar que os grupamentos neuronais imunorreativos a TH no mesencéfalo, diencéfalo, telencéfalo e retina são constituídos por neurônios produtores de DA e, portanto, a imunorreatividade a TH pode ser considerada um marcador molecular confiável para identificação de grupamentos dopaminérgicos (Prakash e Wurst, 2006).
Neste estudo, foram visualizados neurônios IR-TH na região A13, que está medialmente à ZIr, e também de poucos neurônios IR-TH na ZIr. Esses neurônios possuem formato triangular e irregular (Figura 5A e B). De acordo com Abrahamson e Moore (2001), a ZI é considerada um grupamento dopaminérgico (região A13), que recebe projeção do PVT, ou região A14. Todas essas regiões apresentam conectividade e propriedades farmacológicas dos sistemas dopaminérgicos nigroestriatal e mesolímbico (Tonelli e Chiaraviglio, 1995). Outros estudos sugerem que esses neurônios são responsáveis pelo controle de diversas funções hormonais, como a inibição da liberação de prolactina, a secreção do hormônio luteinizante e do hormônio de concentração de melanina, além de ser modular a ingestão hídrica e alimentar (Sita et al, 2003). Alguns trabalhos mostram que a ZI apresenta conectividade dopaminérgica com regiões hipotalâmicas laterais através de estudos com fluorescência (Bjorklund et al, 1975) e que ratos com lesão na ZI comem menos que ratos controle (McDermott e Grossman, 1980). Assim, é provável que neurônios IR-TH da ZI possam estar envolvidos no controle de ingestão alimentar.
46 existe uma hiperatividade da ZI em ratos parkinsonianos (Perier et al, 2000). Além disso, a estimulação de alta frequência da ZI melhora o tremor em pacientes com esclerose múltipla, bem como a acinesia e bradicinesia em Parkinsonianos. Esses resultados iniciais indicam que esta é uma região chave na geração de circuitos de movimentos, e que a sua atividade anormal contribui para os sintomas da doença de Parkinson. No entanto, há muito que não entendemos desse circuito emcasos normais, sendo assim necessário um maior conhecimento acerca de ser funcionamento (Heise e Mitrofanis, 2004), e expandir o conhecimento acerca das células IR-TH, bem como suas características, citoarquitetura e projeções, é fundamental.
47
NOS
A NOS é a enzima responsável pela conversão de L-arginina em óxido nítrico (NO), que tem ação em processos inflamatórios e, no sistema nervoso, atua como um neurotransmissor, mesmo sem preencher todos os requisitos básicos que caracterizam um neurotransmissor convencional. De natureza gasosa e lipofílica, o NO no sistema nervoso atua na modulação de transporte das monoaminas e na nitrosilação de receptores glutamatérgicos N-Metil-D-Aspartato, ou NMDA (Pierucci et al, 2011), influencia a síntese e liberação de outros neurotransmissores, desempenha um importante papel na potenciação a longo prazo, na depressão de longo prazo e na secreção neuroendócrina (Simon et al, 2013). Na ZI do mocó, encontramos células IR-NOS na ZIr, ZIv e ZIc, e sem imunorreatividade na ZId. Em ratos, células IR-IR-NOS são abundantes na ZId e ausentes nas demais regiões (Kolmac e Mitrofanis, 1999; Mitrofanis, 2005), mais uma vez mostrando variabilidade, agora no contexto neuroquímico, entre esses roedores.
48
5-HT
Estudos apontam que a ZI recebe informações de diversos sistemas neuronais, que influenciam a liberação de gonadotrofina, entre eles, o sistema serotonérgico. Verificou-se que a ativação de receptores serotonérgicos 5-HT1A e 5-HT7 nos neurônios IR-5-HT da ZI exercem ação inibitória sobre a liberação do hormônio luteinizante ocasionando uma diminuição de sua concentração no plasma sanguíneo (Siddiqui et al, 2004). A rafe, mais especificamente o núcleo dorsal e o núcleo magno, enviam projeção para a ZI em ratos, ambos atuando na modulação de impulsos fóticos durante o dia subjetivo, ocasionando avanço de fase na atividade circadiana dos neurônios (Medanic e Gillete, 1992). Considerando exclusivamente a ação de fibras serotonérgicas, estas agem alterando o limiar de disparo das células ou ativando segundo-mensageiros, ou seja, atuam diretamente na modulação da atividade dos neurônios (Chapin e Andrade, 2001). Vale relembrar que a ZI desempenha uma grande gama de funções e informações sobre o funcionamento do organismo, então é importante considerar a existência de um sistema de modulação serotonérgica a nível celular para coordenar essas atividades.
49
GFAP
A proteína acídica fibrilar glial, GFAP, é expressa em astrócitos, constituindo um marcador seletivo para essas células gliais. Os astrócitos participam do funcionamento da barreira hematoencefálica, uma vez que, alguns de seus prolongamentos reforçam as junções oclusivas das células que formam a parede dos capilares cerebrais regulando a troca de substâncias dentro do sistema nervoso central (Abbott, 2002). Células gliais apresentam outras funções, como originar populações específicas de neurônios durante o desenvolvimento (Kriegstein e Gotz, 2003), além de um papel neuroprotetor após danos no sistema nervoso central em decorrência de traumas (Anderson et al, 2003), produção de fatores neurotróficos, nutrição e migração neuronal, e estudos mais recentes demonstraram que os astrócitos tem papel ativo na transmissão sináptica, como responder ao aumento da concentração livre de cálcio após a transmissão sináptica (Eysseric et al, 2000), e que existem células gliais que fazem sinapse com neurônios (Bergles et al, 2000), e posteriormente essas sinapses foram encontradas em diversas áreas do sistema nervoso, como hipocampo (Bergles et al, 2000; Mangin et al, 2008) e cerebelo (Lin et al, 2005, Bergles et al, 2010).
Este é o primeiro estudo que detecta células IR-GFAP na ZI, apresentando marcação positiva na ZIr, ZId e ZIc, e negativa na ZIv. A detecção de células da glia na ZI é bastante relevante tendo em vista o grande enfoque na importância dessas células na nutrição e atividade dos neurônios, ainda mais quando se trata de uma região com grande riqueza neuroquímica, funcional e de conexões com outras regiões do sistema nervoso.
CB
50 Em ratos, células IR-CB apresentam pequenos somas, com formato ovoide ou redondo, e compõe entre 5 e 10% do número total de células na região rostral e medial, enquanto que a formação de 20-25% das células no setor caudal (Kolmac e Mitrofanis, 1999). Outro estudo demonstrou uma maior população de células IR-CB na ZIc, maior que na ZId e ZIv (Watson et al, 2013). Em (2004), Mitrofanis et al detectaram a presença de CB em todas as porções da ZI em Macaca fascicularis, distribuída de forma homogênea. No sagui, células IR-CB ocorreram em todas as porções, com maior concentração na porção caudal, e poucas células marcadas nas porções rostral e ventrolateral (Lima, 2008). Outros estudos em sagui detectaram células IR-CB com intensidade de marcação variável, com grandes somas e longos processos axonais na ZIv, enquanto que na ZId poucas células foram coradas, sem a detecção dos processos axonais (FitzGibbon et al, 2000), resultado semelhante ao encontrado por Watson et al (2013), que encontrou mais células na ZIc em comparação às ZId e ZIv.
CR
A CR pertence a uma subclasse de proteínas de gatilho, ou seja, está mais envolvida na modulação de mudanças transitórias de cálcio do que no simples tamponamento intracelular (Dowd et al, 1992), inclusive com diferenças estruturais – apresenta 6 domínios, 5 deles que podem se ligar ao cálcio - relevantes em comparação com a PV – que tem apenas 3 domínios, dois que podem se ligar ao cálcio (Kuznicki et al, 1996). No mocó, a região mais rica em células IR-CR foi a ZId e na região medial da ZIr.
51 Considerando a riqueza neuroquímica e importância funcional da ZI, é esperado que esta possua diversos mecanismos de controle da atividade celular, afim de otimizar o fluxo de informações recebidas, processadas e emitidas para suas conexões pelo sistema nervoso, e o controle da concentração intracelular de cálcio mediado pelas proteínas ligantes de cálcio é fundamental. Vários trabalhos detectaram a presença dessas proteínas na ZI em primatas (Mitrofanis, 2004; Lima, 2008) e roedores (Nicolelis et al, 1992; Trageser et al, 2006; Kolmac e Mitrofanis, 1999), mostrando conservação desse mecanismo durante a evolução desses clados.
PV
Células IR-PV foram encontradas na ZIr e ZIc (Figura 10 D e F), enquanto que na ZId e ZIv não ocorrem células. Isso é uma novidade em relação aos dados registrados na literatura, que destacam que células IR-PV é presente apenas na porção ventral da ZI (Mitrofanis, 2005, Nicolelis et al, 1992; Trageser et al, 2006).
A PV pode ser encontrada em diversas regiões, como o cérebro, músculos, osso e glândulas do sistema endócrino (Jones e Hendry, 1989). Alguns estudos mostram que o tipo celular que contém PV é dominante na ZI ventral (Kolmac e Mitrofanis, 1999; Nicolelis et al, 1992, Mitrofanis, 2005). Em estudos com sagui, células IR-PV também predominaram na ZI ventral, com grande densidade e células de grande tamanho (FitzGibbon et al, 2000). Lima (2008) também verificou uma forte marcação na porção ventral, mas com neurônios mais esparsamente distribuídos. Em Macaca fascicularis, IR-PV apresentam tamanho médio e formato ovoide, presente em todas as regiões da ZI, mas com predominância na porção ventral (Mitrofanis, 2004). Entretanto, a marcação de IR-PV da ZI do mocó mostra que é justamente a ZIv e ZId que não apresenta marcação imunohistoquímica
Verificou-52 se que este comunica-se com a ZI, através de estudos com traçadores anterógrados e com imunohistoquímica para PV na ZI, e que essa conexão era fundamental para que a ZI suprimisse a atividade do núcleo posterior durante um estímulo nociceptivo (Trageser et al, 2006).
5.4 Considerações do funcionamento global da ZI
53 da ZI, ou até mesmo a conjecturar outro modelo. A projeção da retina diretamente para a ZI, bem como a característica dessas fibras (modulatórias) fornecem informações acerca da ausência/presença de luz no ambiente, fundamental para auxiliar e coordenar sua atividade.
54
6. CONCLUSÕES
A técnica de Nissl é eficiente para a delimitação e caracterização citoarquitetônica da ZI do mocó;
A ZIc recebe projeção da retina contralateral, apresentando fibras Classe II e varicosidades do tipo Passagem (maior parte), Terminais e em Cluster, sugerindo um caráter modulatório da informação fótica;
A ZI apresenta células gliais na ZI, sendo o primeiro trabalho a fazer um registro sobre essas celulas em roedores;
A ZI apresenta grande diversidade neuroquímica, sobretudo na porção medial da ZIr, onde encontramos imunorreatividade de todas as substâncias neuroativas investigadas;
A IR-NOS, GFAP e CR auxiliam na delimitação da ZI no nível médio em ZId e ZIv;
55
7. REFERÊNCIAS BIBLIOGRÁFICAS
Abbott, N. J. 2002. Astrocyte-endotelial interactions and blood brain barrier permeability. Journal of Anatomy 200: 629-638.
Abrahamson, E.E., Moore, R.Y,. 2001. The posterior hypothalamic area: chemoarchitecture and afferent connections. Brain Research 889: 1–22.
Alusi, S.H., Worthington, J., Glickman, S., Bain, P.G. 2001. A study of tremor multiple sclerosis. Brain 124: 720 – 730.
Anderson, M.F., Blomstrand, C., Blomstrand, F., Eriksson, P.S., Nilssom, P. 2003. Astrocytes and stroke networking for a survival? Neurochem Res 28: 293-305.
Angelucci, A., Clasca, F., Sur, M. 1996. Anterograde axonal tracing with the subunit B of cholera toxin: a highly sensitive immunohistochemical protocol for revealing fine axonal morphology in adult and neonatal brains. J Neurosci Methods 65:101–112.
Arai, R., Jacobowitz, D.M., Deura, S. 1994. Distribuition of calretinin, calbidin D – 28 and Parvalbumin in the rat thalamus. Brain Res Bull 33:595-614.
Aumann, T.D., Rawson, J.A., Pichitpornchai, C., Horne, M.K., 1996. Projections from the cerebellar interposed and dorsal column nuclei to the thalamus in the rat: a double anterograde labelling study. J Comp Neurol 368: 608–619.
Barthó, P., Freund, T. F., Acsády, L. 2002. Selective GABAergic innervation of thalamic nuclei from zona incerta. European Journal of Neuroscience, Vol. 16, pp. 999 - 1014.
56 Bjorklund, A., Lindvall, O., Nobin, A. 1975. Evidence of a n incerto-hypothalamic
dopamine neuron system in the rat.Brain Research, 89: 29-42.
Blanks, R.I., Clarke, R.J., Fausto, L., Giolli, R.A., Phan, S.V., Torige, Y. 1995. Projections of the lateral terminal acessory optic nucleus of commom marmoset (Callitrhix jacchus) J comp Neurol 354:511-532.
Bergles, D.E., Roberts, J.D.B., Somogyi, P., Jahr, C.E. 2000. Glutamatergic synapses on oligodendrocyte precursor cells inthe hippocampus. Nature 405, 187–191.
Blomstedt, P., Sandvik, U., Linder, J., Fredricks, A., Forsgren, L., Hariz, M. I. 2011. Deep brain stimulation of the subthalamic nucleus versus the zona incerta in the treatment of essential tremor.Acta Neurochir 153:2329–2335.
Cabrera, A. 1961 Catálogo de los mamiferos de America del sur. Ver. Mus, Argentino Cien. Nat. Bernardo Rivadavia. 1-732.
Carvalho, J.C.M. 1969. Notas de viagem de um zoólogo á região das caatingas e áreas limítrofes. Imprensa universitária do Ceará.
Catzeflis, F., Patton J., Percequillo, A., Bonvicino, C. & Weksler, M. 2008. Kerodon rupestris. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.1. <www.iucnredlist.org>. Downloaded on 17 August 2013.
Cavalcante, J.S., Costa, M.S.M.O., Santee, U.R., Britto, L.R.G., 2005. Retinal projections to the midline and intralaminar thalamic nuclei in the common marmoset (Callithrix jacchus).Brain Res. 1043, 42-47.
57 Cavalcanti, J.R.L.P. 2011. Os núcleos dopaminérgicos do mocó (Kerodon rupestris): Caracterização citoarquitetônica e por imunoistoquímica para tirosina-hidroxilase. Programa de pós-graduação em Psicobiologia, UFRN, Natal RN. Dissertação de Mestrado.
Celio, M.R. 1990. Calbindin D-28K and parvalbumin in the rat nervous system. Neuroscience 35: 375-475.
Chapin, E.M., Andrade, R.A. 2001. 5-HT (7) receptor mediated depolarization in the anterodorsal thalamus I. Pharmacological characterization. J Pharmacol. Exp. Ther. 297: 395-402.
Chen, J.C., Chen, P.C., Chiang, Y.C. 2009. Molecular mechanisms psychostimulant addiction. Chang Gung Med. J. 32: 148-154.
Çavdar, S,. Onat, F., Çakmak Y.O,. Saka, E,. Yananli, H.R,. Aker, R,. 2006. Connections of the zona incerta to the reticular nucleus of the thalamus in the rat. J Anat 209: 251-258.
Cooper, H.M., Herbin, M., Nevo, E., 1993. Visual system of a naturally microopthalmic mammal: the blind mole rat, Spalax ehrenbergi. Comp. Neurol. 328: 313–350.
Costa, M.S.M.O., Santee, U.R., Cavalcante, J.S., Moraes, P.R.A., Santos, N.P., Britto, L.R.G. 1999. Retinohypothalamic projections in the common marmoset (Callithrix jacchus): a study using cholera toxin subunit B. J. Comp. Neurol. 415: 393–403.
Da Silva, A.V., Torres, K. R., Haemmerle, C.A., Céspedes, I.C., Bittencourt, J.C. 2013. The Edinger-Westphal nucleus II: Hypothalamic afferents in the rat. J Chem Neuroanat.54, 5 -19.
58 Dowd , D.R., MacDonald, P.N., Komm , B.S., Haussler, M.R., Miesfeld , R.L. 1992. Stable expression of the calbindin- D28k complementary DNA interferes with the apoptotic pathway in the lymphocytes. Mol Endocrinol 6: 1843-1848.
Ebner, F.F., Armstrong-James, M., Diamond, M. E. 1995. Changes in receptive field properties of rat barrelfield neurons following thalamic lesions. Soc. Neurosci. Abstr. 15: 1194-1200.
Edwards, D. A., Isaacs, S. 1991. Zona incerta lesions: effects on copulation, partner-preference and other socio-sexual behaviors. Behav Brain Res 1: 45-50.
Elliott, A. S., Weiss, M. L., Nunez, A.A. 1995. Direct retinal communication with the periamygdaloid area. Neuroreport. 6: 806–808.
Engelberth, R.C.G.J., Pontes, A.L.B., Nascimento, R.B.S.N., Lima, R.R.M.L., Lima, R.R., Toledo, C.A.B., Costa, M.S.M.O., Britto, L.R.G., Cavalcante, J.S. 2008. Discrete retinal input to the parabrachial complex of a new-world primate, the common marmoset (Callithrix jacchus). Neurosci Lett. 443: 99–103.
Eysseric, H., Gonthier, B., Souberyan, A., Richard, M.J., Develoose, D., Barret, L. 2000. Effects of chronic ethanol exposure of acetaldehyde and free radical production by astrocytes in culture. Alcohol 21: 117-125.
Ficalora, A.S., Mize, R.R. 1989. The neurons of the substantia nigra and zona incerta wich project to the cat superior colliculus are GABA imunorreactive. A double label study using GABA immunocytochemistry and lectin retrograde transport. Neuroscience 29: 567-58.
59 Fite, K.V., Janusonis, S. 2002. Optic afferents to the parabrachial nucleus. Brain Res.
943: 9-14.
Fite, K.V., Janusonis, S., Foot, W., Bengston, L. 1999. Retinal afferents to the dorsal raphe nucleus in rats and Mongolian gerbils, J. Comp. Neurol. 414: 469–484.
FitzGibbon, T,. Solomom, S.G., Goodchild, A. K. 2000. Distribuition of calbindin, parvalbumin and calretinin immunoreactivity in the reticular thalamic nucleus of the marmoset: evidence for a medial leaft of incertal neurons. Exp Neurol 164: 371-383.
Foote, W. E., Taber-Pierce, E., Edwards, L. 1978. Evidence for a retinal projection to the midbrain raphe of the cat. Brain Research, 156: 135-140.
Fytagoridis, A., Sjöberg, R.L., Åström, M., Fredricks, A., Nyberg, L., Blomstedt, P. 2013. Effects of Deep Brain timulation in the Caudal Zona Incerta on Verbal Fluency. Stereotact Funct Neurosurg 91: 24 – 29
Gaillard, F., Karten, J. H., Sauve, Y. 2013. Retinorecipient Areas in the Diurnal Murine Rodent Arvicanthis niloticus: A Disproportionally Large Superior Colliculus. The Journal of Comparative Neurology. 521: 1699–1726.
Giber, K., Slezia, A., Bokor, H., Bodor, A., Katona, A., Acsady, L., 2008. Heterogeneous Output Pathways Link the Anterior Pretectal Nucleus With the Zona Incerta and the Thalamus in Rat. The Journal of Comparative Neurology 506: 122–140.
Giolli, P. L. A., Blanks, R.H.I., Lui, F. 2006. The acessory optic system: basic organization with an update on connectivity, neurochemistry and function. Prog Brain Res 151: 407-440.
60 Gorbachevskaya, A. I., Chivileva, O. G. 2008. Structural Organization of the Zona Incerta of the Dog Diencephalon. Neuroscience and Behav Physiology 6: 573-578.
Heise, C., Mitrofanis, J. 2004. Evidence for a Glutamatergic Projection from the Zona Incerta to the Basal Ganglia of Rats. The Journal of Comparative Neurology 468: 482–495.
Herbin, M., Reperant, J., Cooper, H. M. 1994. Visual system of the fossorial molelemmings, Ellobius talpinus and Ellobius lutescens. J. Comp. Neurol. 346: 253-275.
Huntchins, B., Weber, J.T. 1985. The pretectal complex of the monkey: a reinvestigation of the morphology and retinal projections, J comp neurol 232: 425-442.
Itaya, S.K., Van Hoesen, G.W. 1982. Retinal innervation of the inferior colliculus in rat and monkey. Brain Research. 233: 45-52.
Jones, E.G. 2007. The Thalamus VII, Cambridge University Press p 1679.
Kaas, J.H., Huerta, M. F. 1988. The subcortical visual system of primates, Comp. Primate Biol. 4: 327–391.
Kawana, E., Watanabe, K., 1981. A cytoarchitectonic study of zona incerta in the rat. J. Hirnforsch. 22, 535-541.
Kim, U., Gregory, E., Hall, W.C., 1992. Pathway from the zona incerta to the superior colliculus in the rat. J comp Neurol 321: 555-575.
61 Kolmac, C.L., Mitrofanis, J. 1999. Distribution of various neurochemicals within zona incerta: an immunocytochemical and histochemical study. Anat Embryol 199: 265–280.
Kriegstein, A.R., Gotz, M. 2003. Radial glya diversity: a matter of cell fate. Glia 43: 37-43.
Krout, K.E., Kawano, J., Mettenleiter, T.C., Loewy, A.D. 2002. CNS inputs to the suprachiasmatic nucleus of the rat. Neuroscience 110: 73–92.
Kudo, M., Nakamura, Y., Moriizumi, T., Tokuno, H., Kitao, Y. 1988. Direct retinal projections to the lateroposterior thalamic nucleus (LP) in the mole. Neuroscience Letters. 93: 176-180.
Lacher Jr, T. E. 1981. The comparative social behavior of kerodon rupestris and galea spixii and the evolution of behavior in the caviidae. Bulletin of carnegie Museus Natural History. 17: 1-71.
Leak, R, K., Moore, R, Y. 2001. Topographic organization of supraquiasmatic nucleus projection neuron. J Comp Neurol 433: 312-334.
Ling, C., Schneider, G.E., Jhaveri, S., 1998. Target-specific morphology of retinal axon arbors in the adult hamster. Vis Neurosci. 15, 559–579.
Lima, R,R. 2008. A Zona Incerta no Sagui (Callithrix jacchus): Projeção retiniana e análise neuroquímica. Programa de pós-graduação em psicobiologia, UFRN, Natal RN. Dissertação de Mestrado.
62 Lin, S.C., Huck, J.H., Roberts, J.D., Macklin, W.B., Somogyi, P., Bergles, D.E. 2005. Climbing fiber innervation of NG2-expressing glia inthe mammalian cerebellum. Neuron 46: 773–785.
Lima, R.R., Pinato, L., Nascimento, R.B., Engelberth R,C., Nascimento Jr, E.S., Cavalcante, J.C., Britto, L.R., Costa, M.S., Cavalcante, J.S. 2012. Retinal projections and neurochemical characterization of the pregeniculate nucleus of the common marmoset (Callithrix jacchus). J Chem Neuroanat. 44: 34-44.
Marín, F,. Herrero, M.T,. Vyas, S., Puelles, L. 2005. Ontogeny of tyrosine hydroxylase mRNA expression in mid- and forebrain: neuromeric pattern and novel positive regions. Dev. Dyn. 234: 709–717.
Martinet, L., Serviere, J., Peytevin, J. 1992. Direct retinal projections of the “non-image forming” system to the hypothalamus, anterodorsal thalamus and basal telecephalon of mink (Mustela vision) brain. Experimental Brain Research. 89: 373-382.
Mangin, J.M., Kunze, A., Chittajallu, R., Gallo, V. 2008. Satellite NG2
progenitor cells share common glutamatergic inputs withassociated interneurons in the mouse dentate gyrus. J. Neurosci. 28: 7610–7623.
Matteau, I., Boire, D., Ptito, M. 2003. Retinal projections in the cat: a colera toxin B subunit study. Visual neurosci 20: 481-493.
May, P. J., Sun W., Halls, W.C.1997. Reciprocal connections betwenn the zona incerta and the pretectum and superior coliculus of the cat. Neuroscience 77:1091-1114.
63 Merker, B. 2004. The liabilities of mobility: a selection pressure for thetransition to
consciousness in animal evolution. Conscious Cogn,in press.
Meyer, E.L., Harrington, M.E., Rahmani, T. 1993. A phase-response curve to the benzodiazepine chlordiazepoxide and the effect of geniculo-hypothalamic tract ablation. Physiol Behav 53: 237-43.
Mick, G., Cooper, H., Magnin, M. 1993. Retinal projections to the olfactory tubercle and basal telencephalon in primates, J. Comp. Neurol. 327: 205–219.
Mihailoff, G.A. 1995. Orthograde axonal transport studies of projections from the zona incerta and pretectum to the basilar pontine nuclei in the rat. J Comp Neurol 360: 301–318.
Mitrofanis, J. 2002a. Distinctive patterns of connectivity between the zona incerta and the red nucleus of rats. Anat Embryol 205: 283–289.
Mitrofanis, J. 2002b. Evidence for an auditory subsector within zona incerta of rats. Anat Embryol 205: 453–462.
Mitrofanis, J. 2005. Some certainty for the “zona of uncertainty?” exploring the function of the zona incerta. Neuroscience 130: 1–15.
Mitrofanis, J., Mikuletic, L. 1999. Organisation of the cortical projection to the zona incerta of the thalamus. J Comp Neurol 412: 173–185.
Mitrofanis, J., Ashkan, K., Wallace, B. A., Benabid, A. 2004. Chemoarchitectonic heterogeneities in the primate zona incerta: Clinical and functional implications. Journal of Neurocytology 33: 429–440.
64 Moga, M. M., Moore R.Y. 1997. Organization of neural inputs to the supraquiasmatic
nucleus in the rat. J comp Neurol 389: 508-534.
Mok, D., Mogenson, G, J. 1986. Contribution of zona incerta to osmotically indecud drinking in rats. Am J Physiol-Reg I 251: 823-832.
Morin, L. P,. 2013. Neuroanatomy of the extended circadian rhythm system. Experimental Neurology 243: 4–20.
Morin, L. P., Allen, C,N. 2006. The circadian visual system. Brain Res Rev 51: 1–60.
Moore, R.Y., Eichler, V. B. 1972. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 42: 201–206.
Moschovakis, A.K. 1996. The superior colliculus and eye movement control. Curr Opin Neurobiol 6: 811–816.
Mullen, R.J,. Buck, C.R,. Smith, A. M. 1992. NeuN, a neuronal specific nuclear protein in vertebrates. Development 116: 201-211.
Murer, M.G., Pazo, J.H. 1993. Circling behaviour induced by activation of GABAA receptors in the subthalamic nucleus. Neuroreport 4: 1219–1222.
Nakagawa, S., Mizuma, M., Kuchiwa, S. 1998. The retinal projections to the ventral and dorsal divisions of the medial terminal nucleus and mesencephalic reticular formation in the Japanese monkey (Macaca fuscata): a reinvestigation with cholera toxin B subunit as an anterograde tracer. Brain Res. 809: 198-203.
65 Nascimento Jr, E. S., Duarte, R. B., Silva, S. F., Engelberth R. C., Toledo, C.A., Cavalcante, J.S., Costa, M.S.M.O. 2008. Retinal projections to the thalamic paraventricular nucleus in the rock cavy (Kerodon rupestris).Brain Res. 1241: 56-61.
Nascimento Jr, E.S., Sousa, A.P., Duarte, R.B., Magalhães, M.A., Silva, S.F., Cavalcante, J.C., Cavalcante, J.S., Costa, M.S.M.O. 2010a. The suprachiasmatic nucleus and intergeniculate leaflet in the rock cavy (Kerodon rupestris): retinal projections and immunohistochemical characterization. Brain Res. 1320, 34-46.
Nicolelis, M.A., Chapin, J.K., Lin, R.C. 1992. Somatotopic maps within the zona incerta relay parallel GABAergic somatosensory pathways to the neocortex, superior colliculus and brainstem. Brain Res 577: 134–141.
Nicolelis, M.A,. Chapin, J.K,. Lin, R.C. 1995. Development of direct GABAergic projections from the zona incerta to the somatosensory cortex of the rat. Neuroscience 65: 609–631.
Oliveira, F.G. 2013. Aspectos anatômicos do olho e neuroquímicos da retina do mocó (Kerodon rupestris. Programa de pós-graduação em psicobiologia, UFRN, Natal RN. Tese de doutorado.
Paxinos, G, Watson, C. 2007. The rat brain in stereotaxic coordinates. 2ª Ed. Academic Press, San Diego.
Prakash, N., Wurst, W. 2006. Development of dopaminergic neurons in the mammalian brain. Cell. Mol. Life Sci. 63: 187-206.