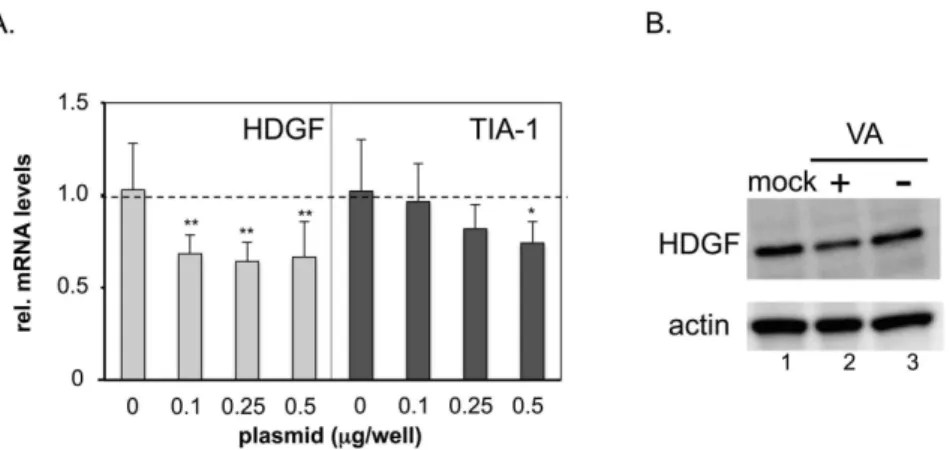
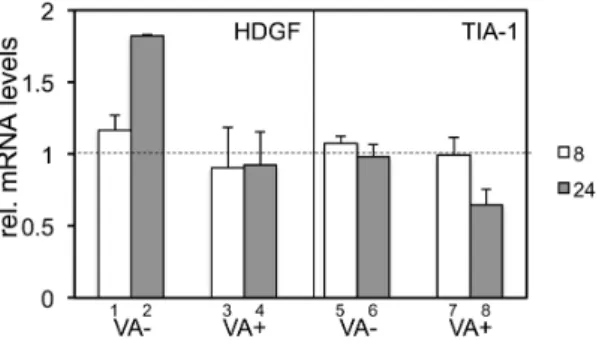
Adenovirus-encoding virus-associated RNAs suppress HDGF gene expression to support efficient viral replication.
Texto
Imagem

Documentos relacionados
Among other immune parameters we have assessed cytokine secretion both at mRNA level by real-time RT-PCR (gene expression) and at protein level by LUMINEX technology. However,
Expression analysis of transmembrane mucins MUC1 and MUC4 at RNA and protein level in various bladder carcinoma cell lines and tabular representation of mucin expression as
(A) protein expression normalized by ACTB and calibrated by the level of a reference sample; (B) the ratio of NPM1 protein expression between tumor and matched non-neoplastic
Using this RNA interference system with in vitro cell studies, the siRNA recombinant retroviruses were screened for efficient knockdown of BCRP/ABCG2 mRNA and protein expression
The study confirmed at the transcriptional level that the constitutive expression of ramA was directly associated with increased expression of multidrug efflux pump AcrAB-TolC
Changes in the expression of ubiquitin, both at the gene and protein level, were investigated in the developing Monodelphis following spinal cord transection, together with
Relative gene expression for all six genes measured by RT-qPCR was compared with RNA-Seq data on days 2 and 5 for the July experiment.. In the first comparison relative expression
With the increase of matrine concentration, the relative expression level of Bcl-2 protein in CNE-2 cells was decreased, and the expression level of Caspase-3 protein was
In accordance with data presented in drug screening, AK-4 compound C39H50O5Si2 displayed strong cytotoxic capability against the growth of two glioblastoma cell lines LN229 and SNB19
