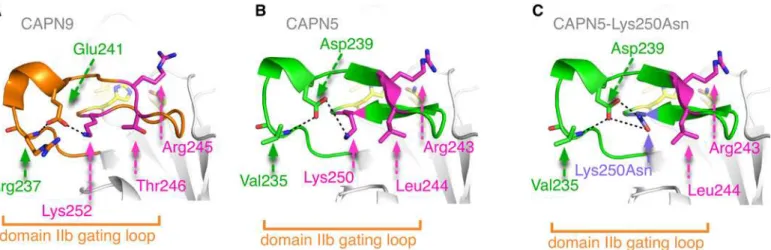
Structural modeling of a novel CAPN5 mutation that causes uveitis and neovascular retinal detachment.
Texto
Imagem
Documentos relacionados
Analisar a relação entre ambiente da prática de enfermagem e os resultados assistenciais, a segurança dos pacientes e o Burnout entre profissionais em unidades
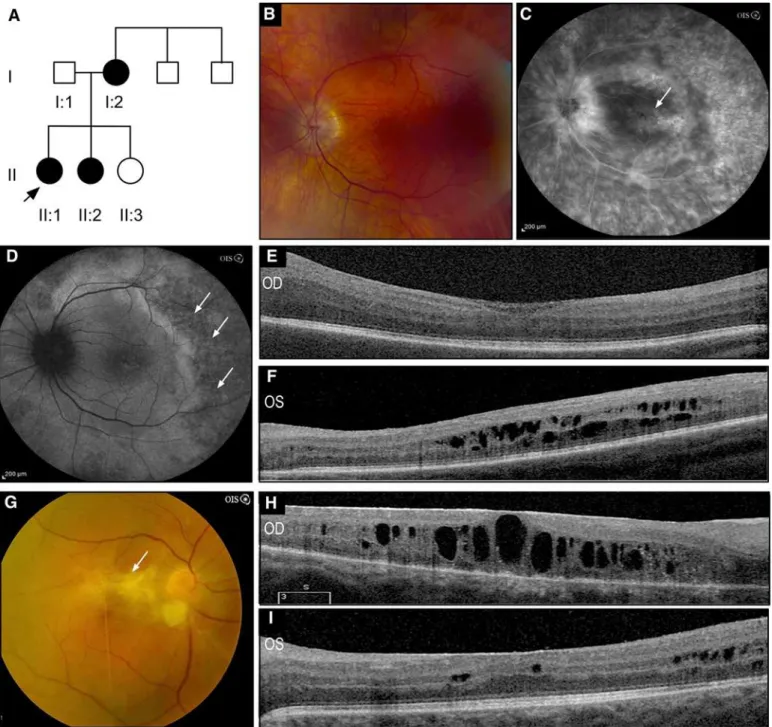
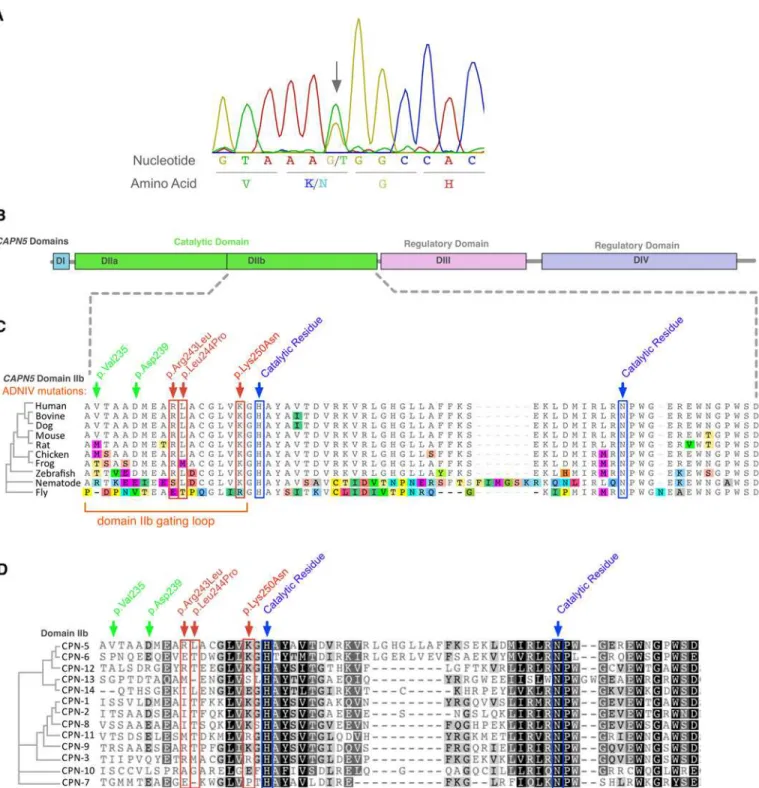
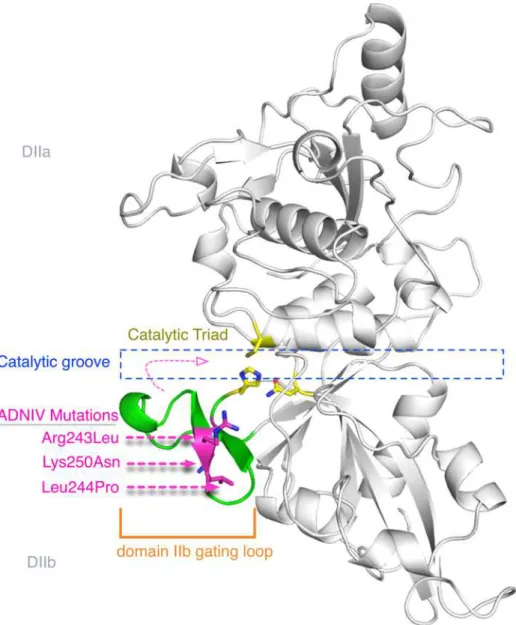
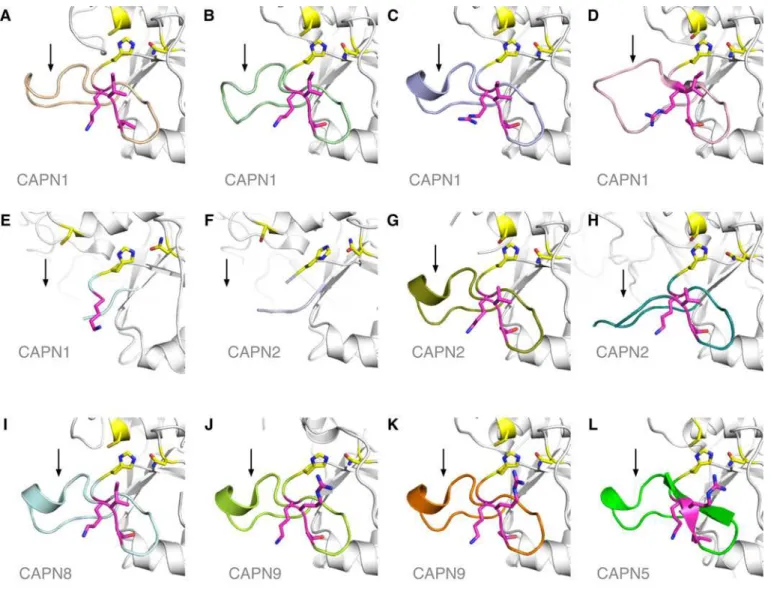
In the 55 unrelated patients screened, four different mutations were identified, of which three are novel: one missense and one nonsense substitutions, and one mutation in the
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
In a recent study on the mutation spectrum of the GALNS gene, it was shown that the three most frequent mutations, p.R386C, p.G301C and p.I113F account for only 20% of the
73 Tabela 4.40 - Matriz de confusão global para o caso binário com as features Amplitude com exclusão das features correlacionadas entre si e Kernel Linear para as
O presente capítulo discorre sobre os fundamentos teóricos do trabalho colaborativo em rede e sobre a ferramenta wiki, busca instrumentos de forma a que se possa compreender o
The main aim of the research project “On the Contribution of Schools to Children’s Overall Indoor Air Exposure” is to study associations between adverse health effects, namely,
Essa variação na taxa de juros influencia nos investimentos em ações, pois quando há aumento da taxa de juros, os investidores da bolsa vendem suas ações para investir em ativos
