Mammary fat of breast cancer: gene expression profiling and functional characterization.
Texto
Imagem
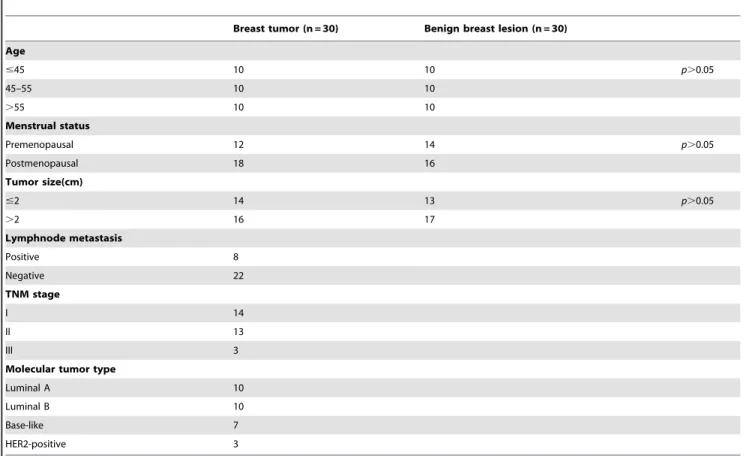
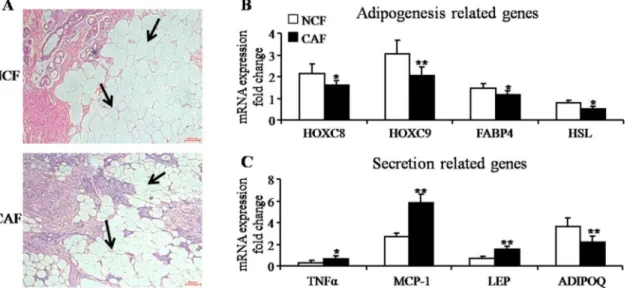
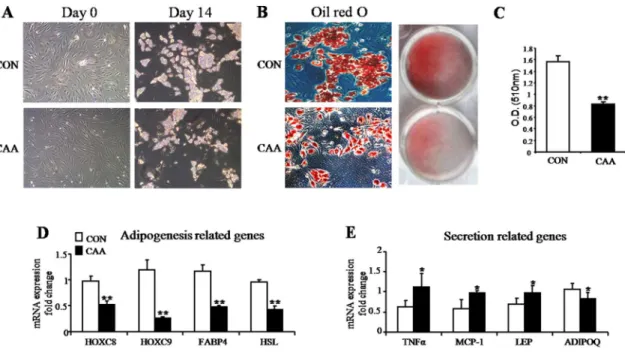
Documentos relacionados
We compared the proliferation of the human breast cancer cell line MDA-MB-231 and of the non-cancer human mammary epithelial cell line MCF-10A in different experimental
2- Ativar o inalador pressionando o botão superior, na posição vertical 3- Esvaziar completamente os pulmões com a boca fora do bucal 4- Colocar os lábios em torno do bucal com
Differential expression of 4 of 6 genes found to be differentially expressed by cDNA microarray analysis in the CSIII tumor versus normal tissue com- parison was validated
nels (VGSC) in the progression of different tumors, such as prostate cancer, small cell lung cancer, breast cancer and others, linking VGSC to the invasion capacity of tumor cells (4
Quantitative gene expression of laminin-332 γ2 in samples of canine mammary cancer (black columns) and in a pool of normal mammary tissues (zero or 0)... A controversial role
Neste sentido, conclui-se que os alunos atribuíram maior importância a motivos intrínsecos do que extrínsecos para se manterem motivados para as aulas de Educação Física,
of breast cancer associated with certain metastatic sites exhibit a characteristic genetic expression, and the immunohistochemical expression of the human epidermal growth factor
In the present study, we identified mutations in exons 9 and 20 of the PIK3CA gene in primary breast tumors from Brazilian breast cancer patients, and we analyzed the
