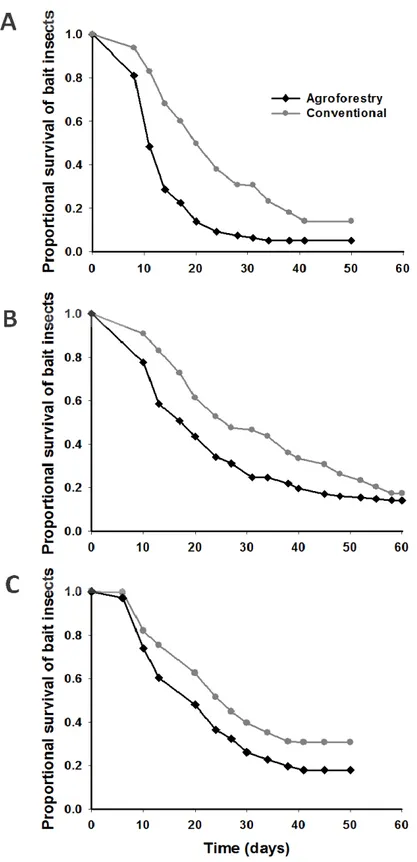
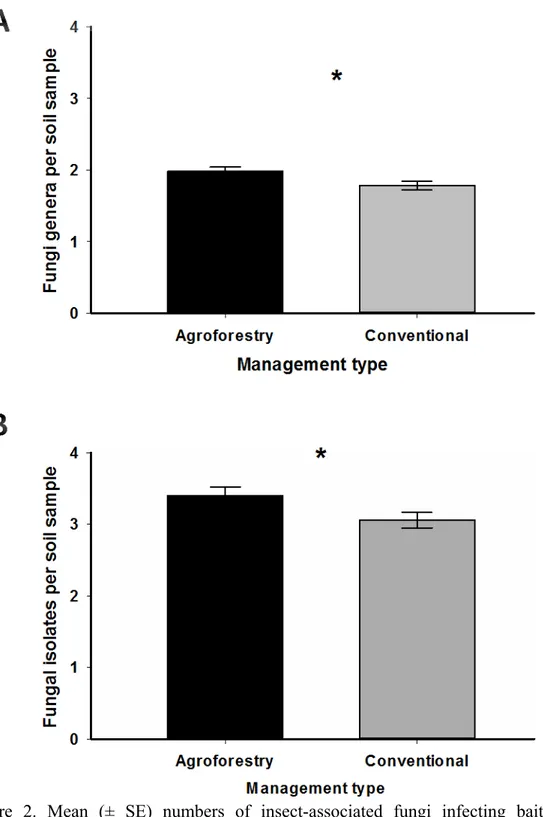
Suppressiveness mediated by entomopathogens in agroforestry and conventional soils
Texto
Imagem

Documentos relacionados
Extinction with social support is blocked by the protein synthesis inhibitors anisomycin and rapamycin and by the inhibitor of gene expression 5,6-dichloro-1- β-
Considerando que aspectos morais são atribuições racistas ao outro “diferente”, e que são estes aspectos que congregam a possibilidade de discriminações de toda ordem, a
As diferenças específicas do Conselho Europeu em relação ao Conselho da União Europeia previsto pelos artigos 202.º e seguintes do TCE são consideráveis: (a)
No caso e x p líc ito da Biblioteca Central da UFPb, ela já vem seguindo diretrizes para a seleção de periódicos desde 1978,, diretrizes essas que valem para os 7
É importante destacar que as práticas de Gestão do Conhecimento (GC) precisam ser vistas pelos gestores como mecanismos para auxiliá-los a alcançar suas metas
O primeiro aspecto é que as diferenças sexuais não podem ser vistas apenas como biologicamente determinadas na acepção meramente genital, antes, são social e
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
