Fungos micorrízicos arbusculares em rizosfera de pinhão-manso
Texto
Imagem
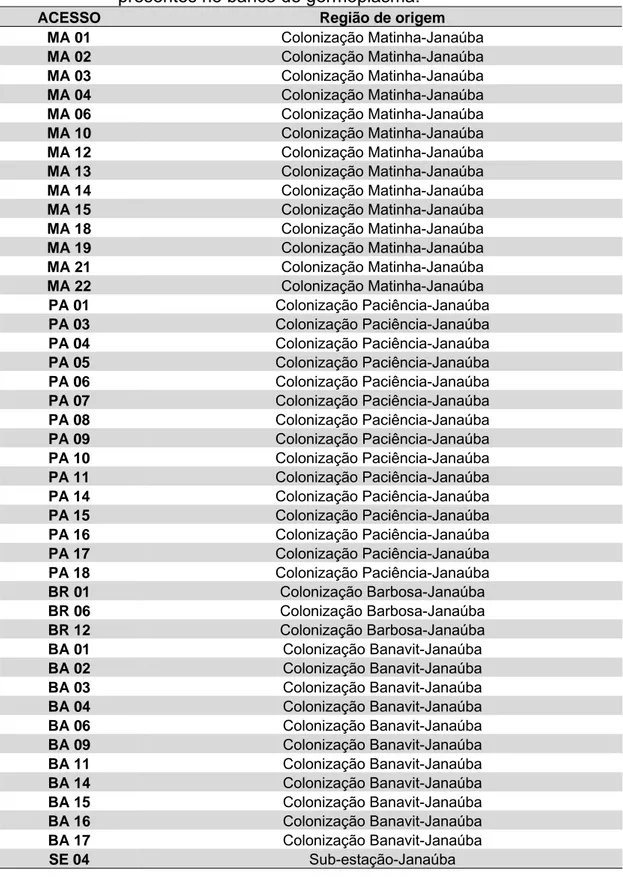
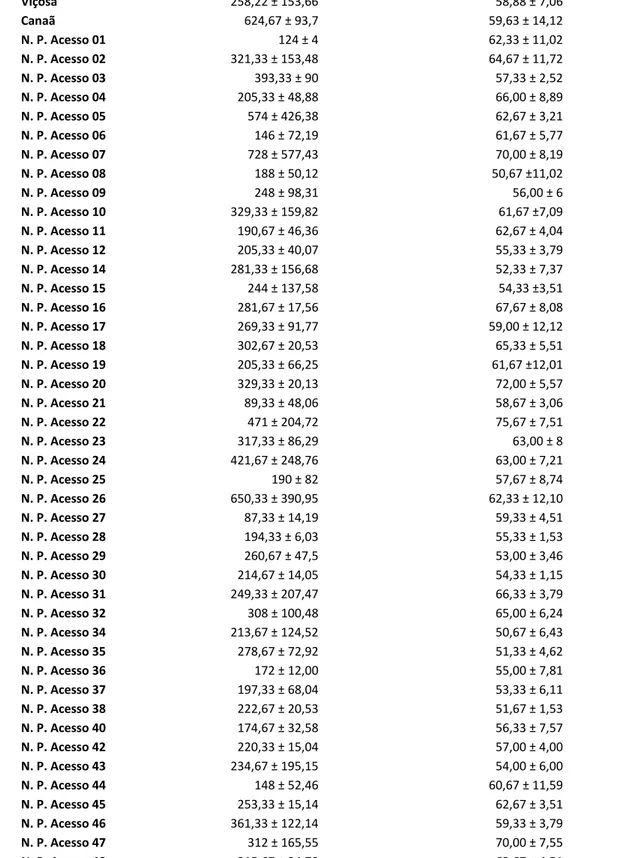

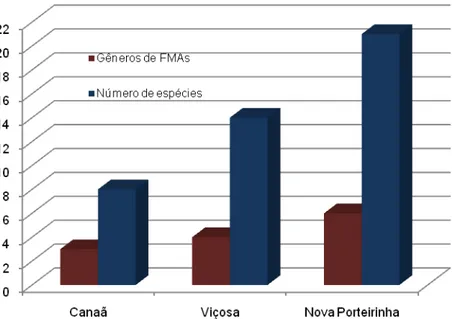

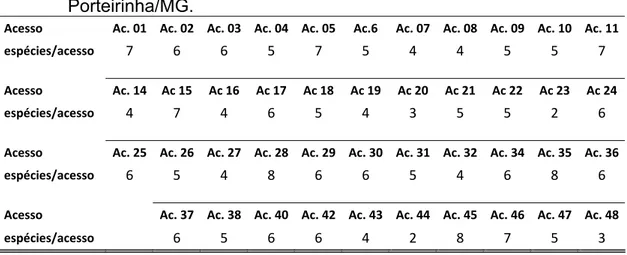


Documentos relacionados
Inspecção Visual Há inspeccionar não só os aspectos construtivos do colector como observar e controlar a comutação (em
A gestão do processo de projeto, por sua vez, exige: controlar e adequar os prazos planejados para desenvolvimento das diversas etapas e especialidades de projeto – gestão de
Como um todo eu frizo e repito: precisamos de um conjunto , primeiro das autoridades, a nossa parte estamos fazendo, para que SIF a carne de sol de Picuí, pra que
Ao final da utilização do sensor, este é removido e os dados armazenados no monitor são transferidos para um receptor (Figura 3) permitindo que os dados (Figura 4) sejam
Konrad Hesse contrapondo a Lassale, naquilo que ele denomina de negação do direito constitucional, compreende que, na verdade, não há como negar a força
O Ideb foi criado no governo Lula em 2007, pelo Instituto Nacional de Estudos e Pesquisas Educacionais Inep como um dos eixos principais do Plano de Desenvolvimento da Educação PDE
Energética no Vale do Jequitinhonha: Neste mesmo ano, em convênio com o IPEF, foram iniciados os primeiros experimentos de manejo da regeneração, onde procurou-se estudar os efeitos
O presente estudo foi realizado com o objetivo de descrever o perfil das mulheres que tiveram parto na maternidade do Hospital Universitário Polydoro Ernani de São Thiago em
