A novel network profiling analysis reveals system changes in epithelial-mesenchymal transition.
Texto
Imagem

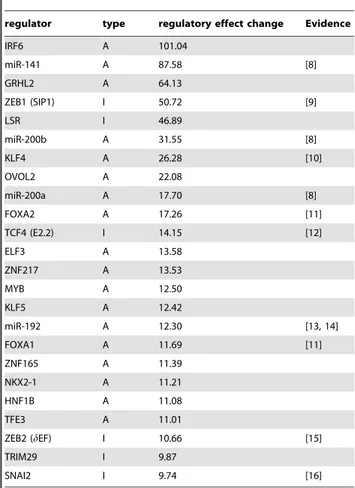
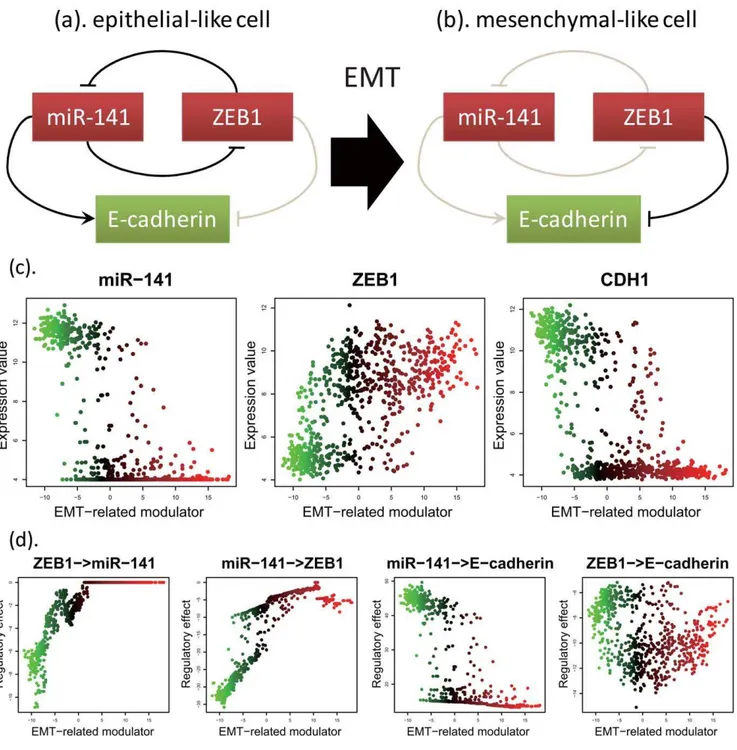
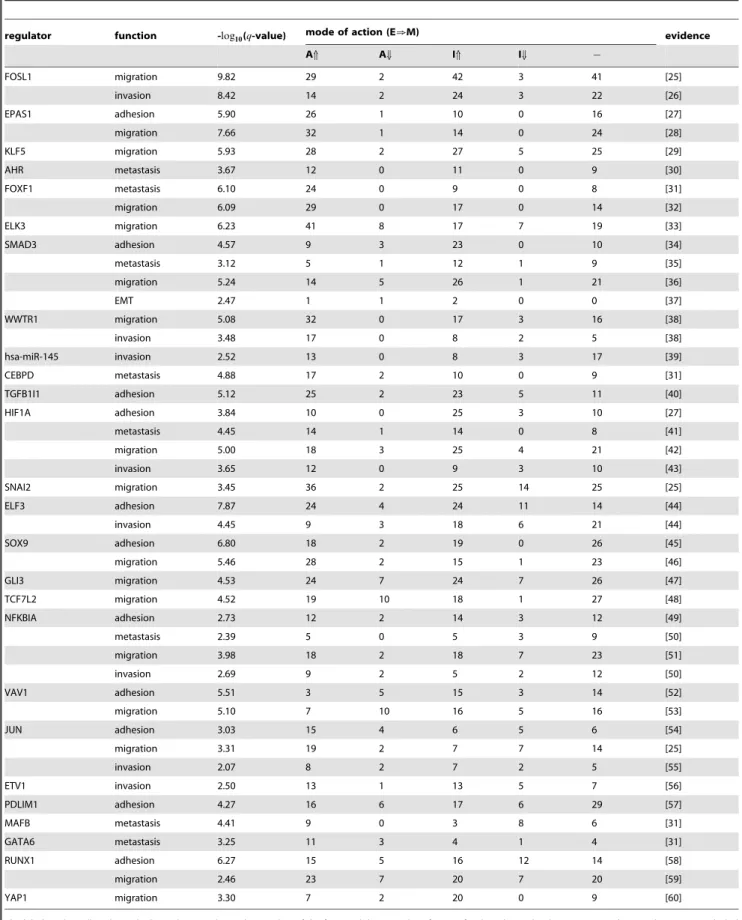
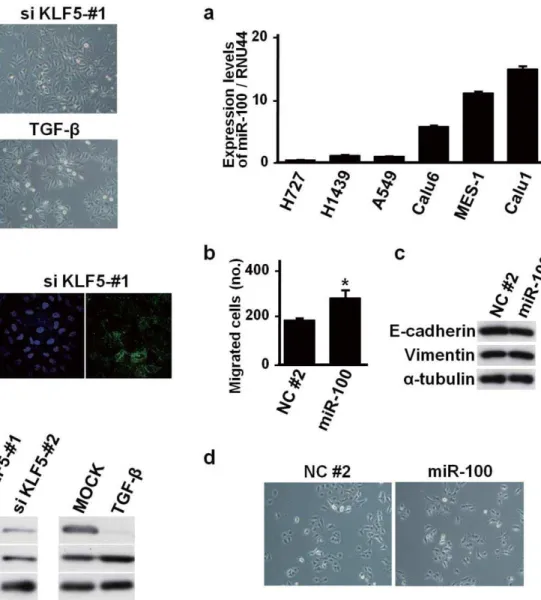
Documentos relacionados
The aim of this study was to correlate the immunohistochemical expression of molecules E-cadherin, N-cadherin and Bmi-1, important markers of the EMT process, and consequently
The concomitant loss of E-cadherin and gain of N-cadherin expression (cadherin switch) has been recently reported in malignant epithelial tumors including head and neck 30
To investigate whether KLF4 regulates EMT in ovarian cancer cells, the epithelial cell marker gene E-cadherin and mesenchymal marker genes snail2 and vimentin were examined in
Firstly, in this study, we showed that miR-124-1 is not methylated in normal blood cells but is hypermethylated in lymphoma and myeloma cell lines, which can be re-expressed
The serum levels of miR-3200, miR-21, and miR-205 at any point during chemotherapy treatment were not predictive for DFS; however, high levels of miR-451 at the time of diagnosis
Upon overexpression of miR-124 in Daoy cells, Nur77 mRNA and protein levels and the mRNA levels of Nur77 target genes decreased, showing that miR-124 affects not only Nur77
indeed promotes EMT in these ovarian cancer cells, we examined the expression of E-cadherin and N-cadherin, two well-established epithelial to mesenchymal makers, in
In order to begin to investigate whether an EMT like process is occurring in SSc sections of forearm skin were stained against the mesenchymal markers FSP-1 and vimentin, epithelial
