FLÁVIO SIQUEIRA D´AVILA
EFEITO DO FÓSFORO, NITROGÊNIO E POTÁSSIO NA PRODUÇÃO DE MUDAS CLONAIS DE EUCALIPTO
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Ciência Florestal, para obtenção do título de Magister Scientiae.
VIÇOSA
Ficha catalográfica preparada pela Seção de Catalogação e Classificação da Biblioteca Central da UFV
T
D’Avila, Flávio Siqueira, 1982-
D259e Efeito do fósforo, nitrogênio e potássio na produção 2008 de mudas clonais de eucalipto / Flávio Siqueira D’Avila.
– Viçosa, MG, 2008. viii, 53f.: il. ; 29cm.
Orientador: Haroldo Nogueira de Paiva.
Dissertação (mestrado) - Universidade Federal de Viçosa.
Inclui bibliografia.
1. Eucalipto - Nutrição. 2. Eucalipto - Propagação por miniestaquia. 3. Clonagem. 4. Eucalipto - Efeito do fósforo. 5. Eucalipto - Efeito do nitrogênio. 6. Eucalipto - Efeito do Potássio. I. Universidade Federal de Viçosa. II.Título.
FLÁVIO SIQUEIRA D´AVILA
EFEITO DO FÓSFORO, NITROGÊNIO E POTÁSSIO NA PRODUÇÃO DE MUDAS CLONAIS DE EUCALIPTO
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Ciência Florestal, para obtenção do título de Magister Scientiae.
APROVADA: 29 de abril de 2008.
_____________________________ _____________________________ Prof. Helio Garcia Leite Prof. Nairam Félix de Barros (Co-orientador) (Co-orientador)
______________________________ _____________________________ Prof. Aloísio Xavier Pesq. Fernando Palha Leite
___________________________________________ Prof. Haroldo Nogueira de Paiva
DEDICO
Ao meu pai, José Evonio D´Avila e à minha mãe, Adna Siqueira D´Avila
pela pessoa que sou hoje.
“Compartilhar os teus conhecimentos é a forma de conseguires a imortalidade”
AGRADECIMENTOS
- Aos meus pais, José Evonio D’Avila e Adna Siqueira D’Avila; - À Universidade Federal de Viçosa, por todos os ensinamentos; - À FAPEMIG – pela cessão da bolsa de estudos.
- À CENIBRA, em especial ao Dr. Fernando Palha Leite e Dr. Gualter Silva pela permissão e total apoio logístico na execução do trabalho;
- Ao meu orientador Prof. Haroldo Nogueira de Paiva, pela confiança depositada em mim, orientação e amizade.
- Aos Profs. Hélio Garcia Leite e Nairam Félix de Barros, por todo apoio durante a execução do trabalho;
- Aos membros da banca examinadora, pelas críticas e sugestões;
- Aos meus amigos de classe, Celso, Eric, Henrique, Isaque, Rodrigo e Sidney, pela amizade e companheirismo durante a graduação.
- Aos meus amigos de república, Carlos e Daniel, à minha irmã Diana, e a meu primo Helder Nilba pela amizade e momentos de descontração;
BIOGRAFIA
FLÁVIO SIQUEIRA D´AVILA, filho de José Evonio D´Avila e Adna Siqueira D´Avila, nasceu em 4 de maio de 1982, na cidade de Vassouras, Estado do Rio de Janeiro.
No ano de 1997, concluiu o 1o grau na Escola Estadual Liddy Mignone, em Paty do Alferes – RJ.
No ano de 2000, concluiu o 2o grau e o curso Técnico em Agropecuária na Escola Agrotécnica Federal de Barbacena - MG.
No ano de 2006, diplomou-se Engenheiro Florestal pela Universidade Federal de Viçosa, em Viçosa – MG.
SUMÁRIO
RESUMO ...vii
ABSTRACT...viii
1. INTRODUÇÃO GERAL...1
2. REVISÃO DE LITERATURA ... 2
2.1. Qualidade da muda... 2
2.2. Nutrição mineral ... 4
2.2.1 Nitrogênio ... 5
2.2.2. Fósforo ... 7
2.2.3. Potássio ... 8
2.3. Fases da produção de mudas de eucalipto... 9
2.3.1. Enraizamento ... 10
2.3.2. Aclimatação à sombra ... 10
2.3.3. Fase de crescimento ... 10
2.3.4. Fase de rustificação ... 10
3. REFERÊNCIAS BIBLIOGRÁFICAS ... 13
CAPÍTULO 1 – EFEITO DO FÓSFORO E NITROGÊNIO NA FASE DE CRESCIMENTO DE MUDAS CLONAIS DE EUCALIPTO... 17
RESUMO ... 17
1. INTRODUÇÃO ... 18
2. MATERIAIS E MÉTODOS ... 19
2.1. Recipiente e substrato ... 19
2.2. Estrutura física ... 19
2.3. Adubações ... 20
2.4. Tratamentos...21
2.5. Características avaliadas ... 22
2.6. Delineamento estatístico e análise dos dados... 22
3. RESULTADOS E DISCUSSÕES ...24
3.1. Características morfológicas ...24
3.1.1. Fertilização nitrogenada ... 25
3.1.2. Fertilização fosfatada ... 26
3.2. Características nutricionais ... 31
4. CONCLUSÕES ...34
5. REFERÊNCIAS BIBLIOGRÁFICAS... 35
CAPÍTULO 2 – EFEITO DO POTÁSSIO NA FASE DE RUSTIFICAÇÃO DE MUDAS CLONAIS DE EUCALIPTO ... 39
RESUMO ... 39
1. INTRODUÇÃO ... 40
2. MATERIAIS E MÉTODOS ... 42
2.1. Recipiente e substrato ... 42
2.2. Estrutura física ... 42
2.3. Adubações ... 43
2.4. Tratamentos...44
2.5. Características avaliadas ... 45
2.6. Delineamento estatístico e análise de dados ...45
3. RESULTADOS E DISCUSSÕES ...46
4. CONCLUSÕES ...51
RESUMO
D´AVILA, Flávio Siqueira, M. Sc., Universidade Federal de Viçosa, abril de 2008. Efeito do fósforo, nitrogênio e potássio na produção de mudas clonais de eucalipto. Orientador: Haroldo Nogueira de Paiva. Co-orientadores: Helio Garcia Leite e Nairam Félix de Barros.
ABSTRACT
D´AVILA, Flávio Siqueira, M. Sc., Universidade Federal de Viçosa, April of 2008. Effect of phosphorus, nitrogen and potassium on the production of clonal seedlings of eucalypt. Adviser: Haroldo Nogueira de Paiva. Co-Advisers: Helio Garcia Leite and Nairam Félix de Barros.
In forest nurseries the mineral fertilization for the production of eucalypt seedlings is a important forestry practice, because greater growth and better quality of seedlings depend very much on appropriate nutrition. The objective of this work was to evaluate the effect of phosphorus, nitrogen and potassium on the quality of seedlings of three clones of Eucalyptus spp. Two experiments were carried out in the seedling nursery of the
1. INTRODUÇÃO GERAL
A produção de mudas é uma das fases mais importantes para o estabelecimento dos povoamentos florestais. A nutrição adequada das mesmas e o uso de substrato de cultivo apropriado são fatores essenciais para assegurar boa adaptação e crescimento após o plantio (Del Quiqui et al., 2004). O emprego de fertilizantes nesta fase é prática usual nos viveiros florestais e funciona também como valiosa ferramenta para controlar o tamanho e o vigor das mudas, a fim de que se possa ajustar um cronograma de plantio às condições climáticas da região (Paiva e Gomes, 1993).
O efeito dos nutrientes, principalmente N, P e K, no crescimento de mudas, tem despertado o interesse dos pesquisadores florestais. Vários trabalhos têm sido feitos para se conhecer a melhor dosagem de adubos, bem como a melhor forma de aplicação na produção de mudas (Paiva e Gomes 1993).
Dentre os nutrientes, o nitrogênio (N) e fósforo o (P) são os que mais comumente limitam o crescimento de mudas na fase inicial de produção (Graciano et al., 2006). No viveiro, na primeira fase de produção de mudas, o crescimento das mudas é lento, pois a plântula direciona a maior parte de sua energia para a expansão da área foliar e a formação de raízes, quando o nitrogênio (N) e o fósforo (P) são os nutrientes mais importantes (Del Quiqui et al., 2004). Para atender a essa demanda, a suplementação dos nutrientes pode ser conseguida com base no estudo das curvas de acúmulo de biomassa e de nutrientes, tendo-se com isso a quantificação da necessidade nutricional numa determinada fase de crescimento, permitindo melhor definição da fertilização a ser empregada (Barros et al., 2000).
Ainda na fase de produção de mudas de eucalipto, aspecto ainda carente de informações vem a ser o efeito que a fertilização potássica pode proporcionar na resistência das mudas aos vários ciclos de umedecimento e secagem a elas impostos usualmente no viveiro para estimular o “endurecimento” e, consequentemente, maior sobrevivência e resistência quando do transplantio para o campo, especialmente em períodos de seca (Barros et al., 1990).
2. REVISÃO DE LITERATURA
2.1. Qualidade da muda
Em virtude da demanda cada vez maior de mudas de espécies florestais e da busca constante de melhor produtividade dos povoamentos, a qualidade das mudas tem sido discutida em vários trabalhos de pesquisa, os quais, na sua totalidade, procuram definir os melhores recipientes, substrato e adubações, adequando-os à produção de mudas de qualidade desejável (Paiva e Gomes, 1993).
A qualidade das mudas é um dos fatores básicos para a obtenção de povoamentos de alta produtividade. E esta qualidade se expressa tanto por características morfológicas como fisiológicas e nutricionais, e é resultante, além de fatores genéticos, dos procedimentos de manejo do viveiro (Silva, 1998).
Segundo Carneiro (1995) os critérios para a classificação da qualidade de mudas baseiam-se, fundamentalmente, em duas premissas de elevada importância: aumento do percentual de sobrevivência das mudas após o plantio; diminuição da freqüência dos tratos culturais de manutenção do povoamento recém implantado.
Stape et al. (2001) concluiram em seu trabalho que a sobrevivência inicial de mudas de Eucalyptus está diretamente relacionada à qualidade (morfológica e fisiológica) e às condições ambientais de campo.
Para Carneiro (1995), atenção tem sido dirigida para os parâmetros morfológicos das mudas, como altura, diâmetro de colo, maturação da parte aérea e desenvolvimento do sistema radicial. Estas características são de fácil avaliação e podem dar boas prescrições de qualidade.
A altura da parte aérea é uma característica de fácil medição e, devido a isso, sempre foi utilizada para estimar o padrão de qualidade de mudas de espécies florestais nos viveiros. É considerada um das mais importantes características para estimar o seu crescimento no campo, além do que sua medição não é destrutiva. Ela fornece excelente estimativa da predição do crescimento no campo (Gomes e Paiva, 2004).
Barros et al. (1978) constataram que a taxa de crescimento em altura das mudas de Eucalyptus grandis, após o plantio no campo, foi inversamente proporcional à altura das mudas no viveiro. Esta depressão no crescimento foi consideravelmente maior nas mudas mais altas com pequenos diâmetros de colo do que com as de maiores diâmetros.
de plantadas. Conclui, também, que as mudas têm que apresentar um diâmetro de colo mínimo, de acordo com a espécie, e que seja compatível com a altura, para que seu desempenho no campo corresponda às expectativas.
O diâmetro do colo é uma variável facilmente mensurável e, por ser obtido sem a destruição da planta, é considerado, por muitos pesquisadores, como um dos mais importantes parâmetros para estimar a sobrevivência, logo após o plantio de mudas de diferentes espécies florestais (Gomes e Paiva, 2004).
O crescimento semelhante entre povoamentos florestais, plantados com mudas de qualidade diferente, poderá ocorrer, porém a mortalidade nos primeiros anos pode apresentar uma estreita relação com o método de produção (Freitas e Klein, 1993) e, conseqüentemente, com a sua qualidade.
O maior crescimento inicial de mudas diminui a freqüência dos tratos culturais, minimizando os custos de implantação do povoamento (Carneiro, 1995), assim como o replantio, por ser uma operação bastante onerosa, sendo dispensável quando a sobrevivência for elevada (Novaes, 1998).
Guerreiro e Colli Junior (1984) apresentaram os seguintes critérios para a seleção de mudas de Eucalyptus urophylla e E. saligna produzidas por sementes: a) altura oscilando entre 15 e 35 cm;
b) diâmetro de colo, no mínimo de 2 mm; c) alto grau de rustificação;
d) sem problemas sanitários aparentes; e) possuir raiz pivotante com forma normal; f) parte aérea sem bifurcações;
g) parte aérea com, no mínimo, três pares de folhas; h) haste sem tortuosidade acentuada.
Segundo Alfenas et al. (2004) as características de uma muda de boa qualidade devem ser obtidas a partir de resultados experimentais para cada material genético, todavia na prática têm-se considerado os seguintes critérios de seleção para mudas de eucalipto produzidas por estacas:
a) Altura: 20-35 cm.
b) Diâmetro do colo: superior ou igual a 4 mm.
c) Número de folhas: quatro ou mais pares de folhas completamente expandidas.
f) Raiz: sistema radicular contendo pelo menos quatro raízes, bem distribuídas na periferia da estaca enraizada e com alta atividade fisiológica, evidenciada pela coloração branca das radicelas.
g) Rusticidade: grau de maturidade parcial dos tecidos foliares e caulinares.
A par das características morfológicas, o estado nutricional desempenha importante papel no vigor das plantas. A manipulação dos níveis de fertilização contribui também para a melhoria dos valores dos parâmetros morfológicos (Carneiro, 1995).
2.2. Nutrição mineral
Os nutrientes minerais possuem funções essenciais e específicas no metabolismo vegetal: podem agir como constituintes da estrutura orgânica, ativadores de reações enzimáticas, carreadores de cargas e osmorreguladores (Marschner, 1995).
A nutrição adequada e o uso de substrato de cultivo apropriado são fatores essenciais na produção de mudas, proporcionando boa adaptação e crescimento após o plantio (Del Quiqui et al., 2004). Em espécies florestais, afeta o crescimento e a qualidade de mudas, ao promover uma boa formação do sistema radicular e permitir uma melhor adaptação e conseqüente sobrevivência após o plantio no campo (Carneiro, 1995).
Os nutrientes necessários para o desenvolvimento das mudas em fase de crescimento são os mesmos para todas as espécies vegetais, sendo que as quantidades extraídas diferenciam-se entre e dentro de cada espécie (Higashi et al. 2002). Em plantas de eucalipto, a nutrição mineral assume grande importância, principalmente quando se visa a utilização mais adequada dos fertilizantes em função das exigências nutricionais dos clones ou classes de clones (Sgarbi et al., 1997).
Segundo Balloni (1984) citado por Camargo (1997), a integração entre a experimentação, análise do solo ou substrato e análise foliar, podem fornecer subsídios para uma orientação segura das necessidades nutricionais das espécies florestais.
Os tecidos de planta possuem alta concentração de nutrientes em boas condições de crescimento. A partir do momento que há falta de um nutriente, as reservas armazenadas no vacúolo são utilizadas e decrescem gradualmente com pouco efeito no crescimento da planta, mas com a redução no teor de nutrientes no tecido (Lambers et al., 1998). Entretanto, uma maior taxa na absorção do nutriente depois da fertilização
nem sempre resultará em maiores crescimentos.
Para Dighton et al. (1993), interações entre nutrientes são importantes para determinar a ótima nutrição em Eucalyptus grandis. O aumento do crescimento é ativado por um ótimo balanço nutricional, expresso principalmente pelas relações N:P, N:K, N:S, P:K e Ca:Mg (Herbert, 1996).
Em mudas de eucalipto, a nutrição por meio da fertirrigação inicia-se 30 a 40 dias após a semeadura ou plantio das estacas, e visa principalmente o fornecimento de N, K e B. Sugere-se que as fertirrigações sejam realizadas uma vez ao dia, na última irrigação (Silveira et al., 2001). Durante o inverno, as doses fertirrigadas devem ser 15 a 20% maiores que as usadas no verão. Nessa estação, essa suplementação nutricional é necessária em virtude da continuidade do processo de lixiviação de nutrientes pela água de chuva e/ou irrigação, e diante da menor atividade metabólica da planta (Silveira et al., 2001).
2.2.1 Nitrogênio
A fertilização nitrogenada tem trazido respostas significativas em eucalipto nos seus estágios iniciais de crescimento – formação de mudas – (Brandi, 1976). Segundo Neves et al. (1990), o uso de nitrogênio geralmente promove ganhos no crescimento de mudas, controlando ainda, o ritmo de crescimento, o tamanho e o vigor das mudas.
Os viveiros a pleno sol nas épocas de intensa precipitação, necessitam de maiores doses de nitrogênio e potássio para compensarem a intensa lixiviação nesse período (Silveira e Higashi, 2003).
A fertilização nitrogenada promove, de maneira geral, aumentos significativos no crescimento em altura nas mudas de espécies florestais (Gomes e Couto, 1984), principalmente nas de eucaliptos, pois isso contribui para uma menor lixiviação e certo controle sobre a velocidade de crescimento e endurecimento das mudas (Neves et al., 1990).
Drew e Saker (1975) concluíram que o efeito do nitrogênio sobre o crescimento radicular ocorria pela sua presença externa à raiz e não interna, uma vez que sua grande mobilidade na planta mantém uma concentração razoavelmente uniforme em todo sistema radicular, independente da parte em contato com o nitrogênio. Aparentemente o nitrogênio externo é sinal para que o fotoassimilado seja transportado para aquelas raízes em contato com o nutriente, o que proporcionaria seu maior crescimento.
A arquitetura do sistema radicular pode ser modulada por manchas de altas concentrações de nitrato, aumentando a ramificação de raízes e o alongamento de raízes laterais em muitas plantas (Assis et al., 2004). O nitrato tem sido apontado como sinalizador, controlando os caminhos de ramificação de raízes, independentemente de seus efeitos como nutriente no metabolismo de nitrogênio (Zhang et al., 1999).
Em experimento realizado por Brandi et al. (1982) avaliando o efeito do N, tanto nítrico quanto amoniacal e suas várias combinações no crescimento de mudas de eucalipto, evidenciou que, o melhor crescimento, considerando peso de matéria seca da parte aérea e de raízes, foi o do tratamento utilizando apenas o N amoniacal. Este mesmo tratamento apresentou melhor aspecto do sistema radicular, denotado por coloração muito mais clara das raízes e maior emissão de raízes finas. As combinações entre as duas formas de N utilizadas não diferiram entre si, sendo, no entanto ligeiramente inferiores ao do tratamento com nitrogênio amoniacal e cerca de 30% superiores ao do tratamento com nitrogênio nítrico.
Higashi et al. (2002) avaliando os efeitos das doses de nitrogênio nas concentrações dos nutrientes em minitouças de clones de Eucalyptus produzidas no sistema de minijardim clonal em canaletão verificaram que o aumento da dose de N na solução provocou reduções nas concentrações foliares de B, Cu, Fe e Mn, em dois clones, e de Zn somente em um clone.
macronutrientes, observando a influência da omissão e presença dos nutrientes sobre o crescimento e composição química das folhas, assim como do caule e raízes. Verificaram que a omissão de nutrientes afetou sensivelmente o crescimento, sendo que, o tratamento com omissão de nitrogênio foi o que mostrou maior efeito depressivo no crescimento. As plantas apresentaram sintomas de carência quando os níveis dos nutrientes nas folhas acusaram os seguintes valores em g Kg-1: N (6,9); P (0,4); K (1,4); Ca (0,4); Mg (0,5) e S (<0,5).
Em estudo com quatro clones de eucalipto, Camargo (1997) observou que as concentrações de nitrogênio diminuíram com a idade das mudas para todos os órgãos analisados, com exceção das raízes de dois clones. A mesma tendência de decréscimo do teor de nitrogênio com a idade foi verificado por Silveira et al. (1995).
Concentrações de N e P, especialmente em folhas, ramos e caules aumentam imediatamente após a fertilização, porém antes de algum efeito no crescimento ser detectado (Graciano et al., 2006).
2.2.2. Fósforo
O teor de fósforo nas plantas é bem menor do que o de nitrogênio e de potássio e, um adequado suprimento do nutriente é importante, no início do crescimento da planta, para a formação dos primórdios vegetativos (Camargo, 1970, citado por Gomes, 2001). A literatura é vasta no tocante a diferenças entre genótipos quanto a absorção, translocação e utilização de fósforo e outros nutrientes. Com relação à cultura de eucalipto, estudos indicam variações nos índices de eficiência de espécies. Desse modo, é provável que as variações na eficiência nutricional possam explicar porque certas espécies apresentam melhor crescimento que outras sob uma mesma condição de fertilidade (Furtini Neto et al., 1996).
Em função da idade, o teor de fósforo nas diversas partes da planta de Eucalyptus tende a decrescer durante a fase de produção de mudas (Silveira et al., 1995).
Brandi et al. (1977), avaliando os efeitos de N-P-K no crescimento de mudas de Eucalyptus citriodora, observaram que o fósforo foi o nutriente que mais contribuiu
para o crescimento das mudas, indicando que bons resultados poderiam ser obtidos somente com a aplicação do mesmo.
muito superiores aos que têm sido obtidos para culturas anuais como: soja, arroz, etc. Isso se deve ao fato de o eucalipto ter, possivelmente, uma baixa eficiência de absorção de P na fase inicial de crescimento.
Brandi et al. (1982), avaliando o efeito de diferentes relações aniônicas de N/P/S sobre o crescimento de plantas de E. grandis em solução nutritiva, constataram que, o tratamento em que apenas a concentração de P (4,38 mg.dm-3) foi aumentada permanecendo N e S nos seus níveis basais (35 mg.dm-3 e 8 mg.dm-3 respectivamente), foi o que apresentou sistema radicular mais vigoroso com raízes claras e parte aérea com bom aspecto nutricional e vegetativo.
Em trabalho realizado por Brandi et al. (1982), avaliando eficiência de absorção de P entre espécies de Eucalyptus, foram obtidos dados que permitiram estabelecer a seguinte ordem decrescente de eficiência: E. cloeziana, E. grandis, E. camaldulensis e E. paniculata. Em experimento complementar, os mesmos autores observaram que,
independente da espécie, o P foi o nutriente mais eficientemente utilizado seguido de Mg, Ca e K, respectivamente. Quando se considera a eficiência de utilização para cada nutriente entre as espécies, observa-se diferenciação entre as mesmas. Assim, para P e K, o E. urophylla foi o que apresentou maior eficiência, seguido do E. paniculata, E. grandis e E. cloeziana, respectivamente.
O acúmulo de P nas raízes e a regulação de seu transporte para a parte aérea parecem desempenhar papel fundamental na resposta das espécies de eucalipto ao nutriente (Furtini Neto et al., 1996).
2.2.3. Potássio
O potássio não faz parte de nenhum composto orgânico, não desempenhando função estrutural na planta. Este macronutriente atua na ativação de aproximadamente 50 enzimas, destacando-se as sintetases, oxiredutases, desidrogenases, transferases, quinases e aldolases (Marschner, 1995).
essa deficiência resulta em baixas taxas de fotossíntese e menor eficiência no uso da água (Bourne et al., 1988).
O potássio está ainda envolvido na síntese de proteínas: plantas com baixos teores de potássio apresentam baixo teor protéico, com acúmulo de compostos de baixo peso molecular como aminoácidos, amidas, aminas e nitratos (Silveira e Malavolta, 2000).
Brandi et al. (1982) verificaram que a deficiência de K propiciou escurecimento de raízes para E. grandis e escurecimento e morte de raízes para E. cloeziana. Para E. paniculata, além dos sintomas acima citados, a deficiência de K refletiu na parte aérea
das plantas, caracterizado pelo murchamento e posterior necrose das folhas. O papel do potássio no balanço hídrico nas plantas provavelmente foi a causa de tais sintomas.
Prezotti (1985) observou que o nível crítico de K no solo, para mudas de eucalipto, varia com o volume de solo utilizado. Quando se utilizam pequenos volumes de solo, as mudas crescem normalmente até um determinado estádio, quando então o potássio torna-se fator limitante, em virtude do pequeno volume de solo explorado pelas raízes, ocorrendo resposta da muda a qualquer adição de potássio que se faça nestas condições.
Respostas positivas de plantas de eucalipto à aplicação de potássio podem ser obtidas quando as mudas são cultivadas em recipientes que contenham pequenos volumes de substrato, mesmo quando os teores deste nutriente forem superiores ao nível crítico. A explicação para tal fato relaciona-se à quantidade, e não à concentração de potássio trocável ou disponível existente no substrato (Neves et al., 1990).
A presença do potássio em níveis crescentes, em experimento realizado por Carneiro (1995), proporcionou um maior endurecimento das mudas de espécies florestais e conseqüente aumento de suas resistências às condições adversas do meio.
2.3. Fases da produção de mudas de eucalipto
2.3.1. Enraizamento
O enraizamento é uma fase de formação de mudas que oscila em média de 15 a 30 dias, dependendo do clone de eucalipto (Ferreira et al., 2004).
O enraizamento adventício de estacas é influenciado por fatores intrínsecos e extrínsecos do material vegetal. Fatores estes como: espécie, presença de indutores e inibidores de enraizamento, juvenilidade dos brotos, presença de gemas e/ou folhas, período de coleta das estacas, período de dormência, estado nutricional, variações nas condições climáticas e sanidade (Cunha, 2006).
O controle da irrigação, temperatura e luminosidade é fundamental para o sucesso do enraizamento, assim como um substrato que proporcione boas características físico-químicas (Alfenas et al., 2004). Algumas empresas adotam também nesta fase a fertilização via água de irrigação, conforme apresentado pelos mesmos autores.
2.3.2. Aclimatação à sombra
Após o enraizamento, as mudas são submetidas à aclimatação sob condição de sombra, preparando-as para a fase de crescimento e rustificação, quando recebem um manejo diferenciado em cada fase, antes da expedição para plantio definitivo no campo (Alfenas et al., 2004).
Em geral, as adubações de cobertura por meio de fertirrigações iniciam-se na fase de aclimatação à sombra, ou seja, em torno de 20 dias após o estaqueamento.
2.3.3. Fase de crescimento
Esta é a fase de maior crescimento das mudas até atingir a altura padrão pré-estabelecida, cuja duração oscila, em média, de 20-30 dias (Alfenas et al., 2004).
2.3.4. Fase de rustificação
De acordo Gomes e Couto (1984), para que a muda seja considerada apta para ser levada para o campo, deve ser sadia e ter grau de resistência que lhe permita sobreviver às condições adversas do meio.
denominado rustificação, cuja duração varia, em média, de 15-20 dias. Em dias muito quentes e ensolarados podem ser efetuadas até três fertirrigações diárias, priorizando formulações com uma reduzida relação N/K. Esta fase é extremamente crítica e reflete diretamente na qualidade final das mudas (Alfenas et al., 2004).
Nos últimos anos a rustificação tem sofrido mudanças importantes em seus princípios e conceitos. Antes, mudas rustificadas, aptas para plantio, continham pouquíssimas folhas, de aspecto coriáceo. Muitas vezes, após serem mantidas por longos períodos no viveiro, apresentavam altura exagerada e sistema radicular agregado, porém enegrecido em virtude da morte das radicelas, e, em alguns casos, com princípio de enovelamento. Atualmente, algumas características ainda prevalecem, como rustificação parcial dos tecidos do caule e das folhas. No entanto, sabe-se, hoje, que uma muda de boa qualidade deve ser bem-enfolhada, possuir um sistema radicular agregado e adequada relação raiz/parte aérea e conter radicelas brancas, que são responsáveis pela maior absorção de água e nutrientes. Ademais, a permanência de mudas por períodos prolongados no viveiro para alcançar a rustificação passou a ser questionada. Em contrapartida, alterações na irrigação e nutrição passaram a ser adotadas, visando atingir a rustificação no menor período de tempo possível e, consequentemente, sem redução da atividade fisiológica. Nesta fase, as mudas tendem a ser mais resistentes a doenças em relação às fases anteriores, em virtude da menor proporção de tecido vegetal tenro e da menor umidade (Alfenas et al., 2004).
Para Silveira et al. (2001), da mesma forma como já foi mencionado, a rustificação tem como objetivo aumentar a resistência e ainda acrescenta que as mudas devem receber fertirrigações com baixa relação N/K na faixa de 1/3 a 1/5, no final do ciclo de produção (75-95 dias) ou quando as mudas atingirem uma altura média de 20-25 cm. Nesta fase, ocorre redução ou corte das adubações juntamente com o aumento do intervalo de irrigação. Isto provoca o engrossamento do caule e o aumento da resistência da muda ao estresse do plantio.
A rustificação de mudas pode ser avaliada pela relação altura/diâmetro de coleto. O valor resultante da divisão da altura da parte aérea de uma muda pelo seu respectivo diâmetro de coleto exprime um equilíbrio de crescimento também denominado de quociente de robustez (Gomes e Paiva, 2004). Para Carneiro (1983), quanto menor for o valor dessa relação maior será a capacidade de as mudas sobreviverem e se estabelecerem no campo.
3. REFERÊNCIAS BIBLIOGRÁFICAS
ALFENAS, A. C.; ZAUZA, E. A. V.; MAFIA, R. G.; ASSIS, T. F. de Clonagem e doenças do eucalipto.Viçosa, MG: Editora UFV, 2004. 442p.
ASSIS, T. F.; FETT-NETO, A. G.; ALFENAS, A. C. Current techniques and prospects for the clonal propagation of hardwood with emphasis on Eucalyptus. In: WALTER. C. e CARSON, M. Plantation forest biotechnology for the 21th century. New Delhi, Índia: Research Sign Post, 2004. p.303-333.
BARROS, N. F.; BRANDI, R. M.; COUTO, L. Efeitos de recipientes na sobrevivência e no crescimento de mudas de Eucalyptus grandis no viveiro e no campo. Revista Árvore, Viçosa, MG, v.2, n.2, p.141-151, 1978.
BARROS, N.F.; NOVAIS, R.F.; NEVES, J.C.L. Fertilização e correção do solo para o plantio de eucalipto. In: BARROS, N.F.; NOVAIS, R.F. Relação solo-eucalipto. Viçosa,MG: Editora Folha de Viçosa, 1990. p.127-186.
BARROS, N.F.; NEVES, J. C.; NOVAIS, R. F. Recomendações de fertilizantes minerais em plantios de eucalipto. In: GONÇALVES, J.L. de M; BENEDETTI, V. Nutrição e fertilização florestal. Piracicaba: IPEF, 2000. p.269-286.
BOURNE, G. H.; JEON, K. W.; FRIEDLONGER, M. Potassium estimation uptake and its role in the physiology and metabolismo flowering plants. International Review of Cytology, v.110, p.205-254, 1988.
BRANDI, R. M. Efeito de adubação NPK no desenvolvimento inicial e na
resistência à seca de mudas de Eucalyptus citriodora Hook. 1976. 69 f.. Dissertação (Mestrado em Fitotecnia) – Universidade Federal de Viçosa, Viçosa, MG.
BRANDI, R.M.; COUTO, L; NETO, F.P. Influência do espaçamento na sobrevivência, freqüência por classe de diâmetro e relação hipsométrica altura-diâmetro do Eucalyptus urophylla. Revista Ceres, v. 24, n.136, p.617-627, 1977.
BRANDI, R. M., BARROS, N. F.; NOVAIS, R. F.; NEVES, J.C.L.; GOMES, J. M. Nutrição e dinâmica de elementos minerais em plantios de eucalipto no estado de Minas Gerais. Relatório anual apresentado pela coordenação do contrato
IBDF/UFV/SIF ao instituto Brasileiro de Desenvolvimento Florestal. Viçosa, MG: SIF/UFV, 1982.
CAMARGO, M.A.F. Matéria seca, concentração e conteúdo de macronutrientes em mudas de clones de eucalipto, em função da idade. 1997. 94p. Dissertação (Mestrado em Solos e Nutrição de Plantas) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, SP.
CUNHA, A. C. M. C. M. Relações do estado nutricional de minicepas e condições meteorológicas com o número e o enraizamento de miniestacas de eucalipto. 2006. 99f.. Dissertação (Mestrado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa, MG.
DEL QUIQUI, E. M.; MARTINS, S. S.; PINTO, J. C.; PARAZZI, P. J. A.; MUNIZ, A. S.; Crescimento e composição mineral de mudas de eucalipto cultivadas sob condições de diferentes fontes de fertilizantes. Acta Scientiarum. Agronomy. Maringá, v.26, n.3, p.293-299, 2004.
DIGHTON, J.; JONES H.E.; POSKITT, J.M. The use of nutrient bioassays to assess the response of Eucalyptus grandis to fertilizer application. 1: Interaction between nitrogen, phosphorus and potassium in seedling nutrition, Canadian Journal of Forest Research, n.23, p.1–6, 1993.
DREW, M. C. e SAKER, L. R. Nutrient supply and the growth of the seminal root system in barley. Journal of Experimental Botany, Cary, USA, v.26, p.79-90, 1975. FERREIRA, E. M.; ALFENAS, A. C.; MÁFIA, R. G.; LEITE, H. G.; SARTORIO, R. C.; PENCHEL FILHO, R. M. Determinação do tempo ótimo do enraizamento de miniestacas de clones de Eucalyptus spp. Revista Árvore, Viçosa, MG, v.28, n.2, p.183-187, 2004.
FREITAS, A. J. P.; KLEIN, J. E. M. Aspectos técnicos e econômicos na mortalidade de mudas no campo. In: CONGRESSO FLORESTAL PANAMERICANO, 11.;
CONGRESSO FLORESTAL BRASILEIRO, 7., 1993, Curitiba. Anais... São Paulo: Sociedade Brasileira de Silvicultura, 1993. p.736.
FURTINI NETO, A. E.; BARROS, N. F.; GODOY, M. F.; NOVAIS, R. F. Eficiência nutricional de mudas de Eucalyptus em relação a fósforo. Revista Árvore, Viçosa, MG, v.20, n.1, p.17-28, 1996.
GOMES, J. M.; COUTO, L.; Produção de mudas de Eucalyptus spp. no estado de Minas Gerais – Evoluções e tendências. In: SIMPÓSIO INTERNACIONAL: Métodos de produção e controle de qualidade de sementes e mudas florestais, 1984, Curitiba: UFPR/FUPEF, 1984 p.440-458, 1984.
GOMES, J. M. Parâmetros morfológicos na avaliação da qualidade de mudas de
Eucalyptus grandis, produzidas em diferentes tamanhos de tubetes e de dosagens
de N-P-K. 2001. 166f.. Tese (Doutorado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa, MG.
GOMES, J. M.; PAIVA, H. P. Viveiros Florestais (propagação sexuada). Viçosa: UFV, 2004, 116p. (Caderno didático 72).
GRACIANO, C.; GOYA, J. F.; FRANGI, J. L.; GUIAMET, J. J. Fertilization with phosphorus increases soil nitrogen absorption in young plants of Eucalyptus grandis. Forest Ecology and Management, v.236, p.202-210. 2006.
INTERNACIONAL: Métodos de Produção e controle de qualidade de sementes e mudas florestais,1984, Curitiba: UFPR/FUPEF, 1984, p.127-133.
HERBERT, M.A. Fertilizer and eucalyptus plantations in South Africa. In: P.M. ATTIWILL and M. ADAMS. Nutrition of Eucalypts, Australian: CSIRO Publishing,1996, p.303–325.
HIGASHI E. N.; SILVEIRA, R. L. V. A.; GONÇALVES, A. N. Nutrição e adubação em minijardim clonal hidropônico de Eucalyptus. IPEF, 2002. (Circular técnica 194). LAMBERS, H.; CHAPIN, F. S.; PONS, Y. L. Plant Physiological Ecology. New York: Springer-Verlag,1998, 540p.
MARSCHNER, H. Mineral nutrition of higher plants. London: Academic Press. 1995. 889p.
MARTINEZ, H.E.P.; BARBOSA, J.G. O uso de substratos em cultivos hidropônicos. Viçosa: UFV, 1999. 49p. (Cadernos didáticos, 42).
NEVES, J. C. L.; GOMES, J. M.; NOVAIS, R. F.; Fertilização mineral de mudas de eucalipto. In: BARROS, N. F.; NOVAIS, R. F. Relação solo-eucalipto. Viçosa, MG, 1990, p.100-124.
NOVAES, A. B. Avaliação morfológica da qualidade de mudas de Pinus taeda L. produzidas em raiz nua e em diferentes tipos de recipientes. 1998. 116 p. Tese (Doutorado). Universidade Federal do Paraná, Curitiba.
PAIVA, H. N. de; GOMES, J. M. Viveiros florestais. Viçosa, MG: UFV, 1993. 56p. PREZOTTI, L.C. Nível crítico de potássio no solo para a produção de mudas de eucalipto. 1985. 58p. Tese (Mestrado em Solos e Nutrição de Plantas) – Universidade Federal de Viçosa, Viçosa, MG.
SGARBI, F.; SILVEIRA, R.L.V.A.; TAKAHASHI, E.; CAMARGO, M.A.F. Crescimento e produção de biomassa de clone de Eucalyptus grandis x Eucalyptus urophylla em solução nutritiva com omissões de macronutrientes, boro e zinco. In: IUFRO CONFERENCE ON SILVICULTURE AND IMPROVEMENT OF
EUCALYPTUS, Salvador, 1997. Anais da Conferência IUFRO sobre Silvicultura e Melhoramento de Eucalipto, v.3: Silvicultura, produtividade e utilização de eucaliptos. Salvador, 1997. p.92-97.
SILVA, M.R. Caracterização morfológica, fisiológica e nutricional de mudas de
Eucalyptus grandis (Hill ex. Maiden) submetidas a diferentes níveis de estresse
hídrico durante a fase de rustificação. 1998. 105p. Dissertação (Mestrado em Engenharia Florestal) - Setor de Ciências Agrárias, Universidade Federal do Paraná, Curitiba, PR.
SILVEIRA, R. L. V. A.; MALAVOLTA, E. Nutrição e adubação potássica em Eucalyptus. Informações Agronômicas, n.91, p.1-12, setembro 2000. (Potafós - encarte técnico).
SILVEIRA, R. L. V. A.; HIGASHI, E. N.; SGARBI, F.; MUNIZ, M. R. A. Seja o doutor do seu eucalipto. Informações Agronômicas, n.93, p.1-32, março 2001 (Potafós – encarte técnico).
SILVEIRA, R. L. V. A.; HIGASHI, E. N. Fertirrigação na produção de mudas de Pinus. Addubare, n.6, 2003. 15p. (Circular Técnica).
STAPE, J. L.; GONÇALVES, J. L. M.; GONÇALVES, A. N. Relationships between nursery practices and field performance for Eucalyptus plantations in Brazil. New Forests, v.22, p.19–41, 2001.
CAPÍTULO 1 – EFEITO DO FÓSFORO E NITROGÊNIO NA FASE DE CRESCIMENTO DE MUDAS CLONAIS DE EUCALIPTO
RESUMO
1. INTRODUÇÃO
A correta nutrição de mudas de eucalipto constitui-se em fator essencial para o estabelecimento de florestas de alta produtividade (Silva et al., 2004), sendo que, a nutrição adequada e o uso de substrato de cultivo apropriado influenciam diretamente na adaptação e crescimento das mesmas após o plantio em campo (Del Quiqui et al. 2004).
Dentre os nutrientes, o nitrogênio é o que tem proporcionado maior crescimento de plantas. De forma geral, a fertilização nitrogenada promove uma aceleração no crescimento, e conseqüentemente uma redução no tempo de formação das mudas quando aplicada em dosagem adequada, considerando as exigências para cada espécie e/ou material genético.
O nitrogênio é o quarto elemento mais abundante na planta, depois do carbono, hidrogênio e oxigênio, sendo um constituinte de aminoácidos, nucleotídeos e coenzimas. Normalmente, estreita relação entre o teor de nitrogênio e o crescimento das plantas é observada, visto que, sob condição de deficiência, ocorre a inibição da síntese de clorofila e consequentemente a diminuição da fotossíntese (Epstein, 1975).
O fósforo é outro nutriente de elevada importância na fase inicial de crescimento de mudas de eucalipto. Grande número de estudos, em muitas espécies tem mostrado que o suprimento de P na fase inicial da vida da planta é fundamental para o ótimo crescimento (Grant et al., 2001).
Em mudas de eucalipto, respostas positivas à adubação fosfatada têm sido generalizadas, sendo que no estado muito jovem são pouco efetivas quanto à absorção de P, necessitando de um teor de P no solo muito mais elevado que aquele requerido pela maioria das culturas (Barros et al., 1982). Na maioria das vezes, a adubação fosfatada resulta em maior crescimento das mudas (Paiva e Gomes, 1993) e, quando o fósforo não é aplicado ao substrato, as mudas não se desenvolvem (Neves et al., 1990).
2. MATERIAIS E MÉTODOS
Os ensaios foram conduzidos no viveiro de mudas da Celulose Nipo-Brasileira S. A. – CENIBRA em Belo Oriente - MG. O município localiza-se na região do Vale do Rio Doce, com clima do tipo Cwa (subtropical, chuvoso e mesotérmico), segundo a classificação de Köeppen. Apresenta precipitação média anual de 1.233 mm. Temperatura média anual de 21ºC, com máxima média de 27ºC e mínima média de 14ºC. O viveiro de mudas localiza-se na latitude 19º17'49" sul, longitude 42º23'26" oeste e, altitude de 233 m (Mafia et al., 2006).
Todos os procedimentos de análise química e pesagem de material vegetal foram realizados no laboratório de solos e análise de plantas da mesma empresa.
Foram utilizadas miniestacas oriundas de três clones de Eucalyptus grandis x Eucalyptus urophylla cultivados em minijardim clonal em leito de areia. Os clones
utilizados foram o 386; o 1213 e o 3837.
2.1. Recipiente e substrato
O estaqueamento foi realizado em substrato acondicionado em tubetes plásticos com dimensões de 12,5 cm de comprimento, 2,7 cm de diâmetro da abertura superior, 0,9 cm de diâmetro de abertura inferior e volume de 50 cm3, com seis estrias internas. Como suporte para os tubetes foram utilizadas bandejas com dimensões de 58,9 cm x 38,6 cm com capacidade para 176 mudas. O substrato utilizado foi vermiculita e casca de arroz carbonizada na proporção de 1:1, em volume.
2.2. Estrutura física
As mudas iniciaram enraizamento em casa de vegetação, permanecendo por 18 dias. Após este período foram transferidas para casa de sombra permanecendo nesta estrutura por mais 6 dias e, por fim, as mudas foram levadas para o estaleiro por mais 18 dias. Nesta última estrutura realizaram-se os tratamentos com as variações nas doses de nitrogênio via fertirrigação.
2.3. Adubações
A concentração de nutrientes e fontes utilizadas na adubação de substrato para todos os tratamentos encontra-se no Quadro 1.
A concentração de nutrientes e fontes utilizadas na fertirrigação de mudas em casa de sombra e estaleiro para todos os tratamentos encontra-se no Quadro 2.
Quadro 1 - Concentração de nutrientes e fontes utilizadas na adubação de substrato de enraizamento.
Nutrientes Substrato de enraizamento
mg de nutriente por litro
P *
(super fosfato simples)
N 146,0
(sulfato de amônio)
K 99,0
(cloreto de potássio)
S 176,0
B 4,9
(ácido bórico)
Cu 5,6
(sulfato de cobre)
Mn 5,1
(sulfato de manganês)
Zn 6,0
(sulfato de zinco)
Quadro 2 - Concentração de nutrientes e fontes utilizadas na fertirrigação de mudas em casa de sombra e estaleiro.
Nutrientes Casa de sombra Estaleiro
mg de nutriente por litro
N 56 *
(nitrato de cálcio)
P 42 108
(ácido fosfórico)
K 87 225
(cloreto de potássio)
Ca 49 127
Mg 61 159
(sulfato de magnésio)
S 20 53
B 0,80 2,1
(ácido bórico)
Cu 0,19 0,5
(sulfato de cobre)
Fe 3,50 9,0
(sulfato ferroso)
Mn 0,80 2,0
(sulfato de manganês)
Mo 0,04 0,11
(molibdato de sódio)
Zn 0,11 0,28
(sulfato de zinco)
* As doses de nitrogênio foram aplicadas de acordo com os tratamentos descritos no tópico a seguir.
2.4. Tratamentos
Os tratamentos, em esquema fatorial, foram compostos por quatro doses de fósforo em substrato de enraizamento e cinco doses de nitrogênio aplicadas via água a partir da fase de crescimento (vigésimo quarto dia). Foram três repetições por tratamento, sendo que cada repetição continha 12 mudas.
A fonte de P utilizada foi o superfosfato simples (18% P2O5). As quantidades de
P aplicadas em substrato foram: P1 - 0 (zero) mg.dm-3 (testemunha); P2 - 349 mg.dm-3;
Para o N, as fontes utilizadas foram o cálcio nítrico (10% de N) e a uréia (44% de N). As doses de N utilizadas foram:
N1 - 0 (zero) mg.L-1 (testemunha); N2 - 73 mg.L-1;
N3 - 146 mg.L-1; N4 - 219 mg.L-1; N5 - 292 mg.L-1.
Ao final do dia, realizava-se a fertilização com auxílio de um regador sendo aplicado 12 litros da solução nutritiva para cada tratamento.
2.5. Características avaliadas
Foram avaliadas características nutricionais (concentrações foliares) e características morfológicas: altura da parte aérea, diâmetro de colo, peso de matéria seca total (PMST) e a relação peso de matéria seca da parte aérea/peso de matéria seca de raiz. Os instrumentos utilizados nas determinações foram: régua, paquímetro (precisão de 0,05 mm) e balança digital. Considerou-se como altura da parte aérea o comprimento entre a base do caule em contato com a superfície do substrato e a base do par de folhas mais jovem.
As medições de altura e de diâmetro de colo das mudas foram realizadas no 40º e no 41º dia após o estaqueamento. No 42º dia se realizou a coleta do material vegetal para a pesagem e análise química das folhas.
Para a obtenção do peso de matéria seca, parte aérea e raízes foram colhidas separadamente, lavadas em água destilada e secas em estufa com circulação forçada de ar até atingirem peso constante.
Para a análise química foram colhidos os penúltimos e os antepenúltimos pares de folhas recém maduras.
2.6. Delineamento estatístico e análise dos dados
O delineamento estatístico adotado foi o inteiramente casualizado. O experimento foi composto por 20 tratamentos (4 doses de P e 5 doses de N) com três repetições.
matéria seca total (PMST), sendo os dados dessas variáveis submetidos à análise de variância e de regressão.
3. RESULTADOS E DISCUSSÕES
3.1. Características morfológicas
O resumo das análises de variância para as características morfológicas é apresentado na Quadro 3.
Quadro 3 - Resumo das análises de variância para altura, diâmetro de colo e peso de matéria seca total (PMST) de mudas avaliadas aos 42 dias após estaqueamento.
Quadrado médio FV GL
Altura Diâmetro de colo PMST
Clone 2 15,30* 0,01029* 0,5566*
Dose P 3 52,06* 0,00428* 0,3735*
Dose N 4 41,99* 0,00528* 0,4573*
ClonexP 6 1,80 ns 0,00007 ns 0,0169 ns
ClonexN 8 0,77 ns 0,00015 ns 0,0052 ns
NxP 12 0,73 ns 0,00010 ns 0,0103 ns
ClonexPxN 24 0,95 ns 0,00013 ns 0,0121 ns
Resíduo 120 1,81 0,00014 0,0120
Total 179 * P < 0,05.
Os resultados da análise de variância apresentados na Quadro 3 mostraram que o fator dose de P e dose de N avaliados isoladamente foram significativos para todas as características avaliadas.
Não existiu significância nas interações Clone x P, Clone x N e Clone x N x P, ou seja, os três clones responderam de forma semelhante às aplicações de N e P. Desta forma, o ajuste de cada equação foi efetuado a partir dos dados dos três clones de forma conjunta.
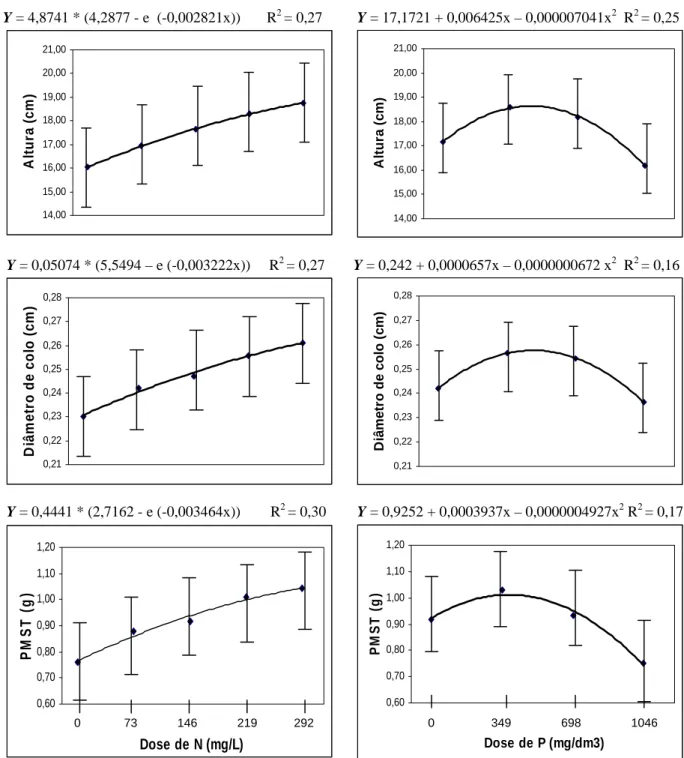
3.1.1. Fertilização nitrogenada
Na Figura 1 constam as características morfológicas altura, diâmetro de colo e peso de matéria seca total (PMST) em função das doses de N e os modelos de crescimento ajustados para cada característica. Ao variar as doses de nitrogênio, foi observado que as maiores doses de N proporcionaram os maiores valores em altura, indicando que o substrato não dispunha de quantidade suficiente do nutriente para atender a demanda das mudas. Tal fato ocorreu devido ao pequeno volume de tubete utilizado aliado às freqüentes irrigações, favorecendo perdas através da lixiviação. Segundo Neves et al. (1990), a perda de nutrientes por lixiviação é considerada alta quando se produzem mudas em tubetes, tanto pelo pequeno volume de substrato quanto pelas irrigações realizadas.
Efeito positivo da adubação nitrogenada em espécies florestais foi verificado
também por Marques et al. (2006). Os autores, avaliando o efeito de doses de N no
crescimento de mudas de sabiá (Mimosa caesalpiniaefolia), constataram que a altura foi
significativamente afetada por doses de N, aumentando linearmente com o acréscimo
das doses, independente do tipo de solo e da fonte aplicada. Segundo Paiva e Gomes
(1993), a aplicação de nitrogênio promove, de maneira geral, na produção de mudas, aumentos significativos no crescimento em altura, entretanto, doses mais elevadas podem apresentar resultados inversos aos esperados.
O diâmetro de colo apresentou comportamento semelhante ao observado para altura em função de doses de N, onde os maiores valores foram encontrados nas maiores doses. Neste caso, as maiores doses de N seriam as mais recomendadas, visto que, altos valores de diâmetro de colo são desejáveis quando se considera a qualidade das mudas. Segundo Gomes e Paiva (2004), as mudas devem apresentar altos valores de diâmetros de colo para melhor equilíbrio do crescimento da parte aérea. Essa variável é reconhecida como um dos melhores, senão o melhor indicador do padrão de qualidade de mudas (Moreira e Moreira, 1996) e, normalmente, é a mais observada para indicar a capacidade de sobrevivência da muda no campo (Daniel et al. 1997).
de N provocam um desequilíbrio na proporção peso da parte aérea/peso de raiz, favorecendo o crescimento da parte aérea. Wilkinson e Langdale (1974) reafirmam que a adubação nitrogenada é responsável pelo maior crescimento relativo da parte aérea que do sistema radicular. A explicação para este fato é que, quando o nitrogênio é o fator limitante do crescimento, ocorre na planta um acúmulo de carboidratos, tanto na parte aérea como nas raízes. Como estes carboidratos não podem ser utilizados, nem na formação de novos tecidos nem no crescimento, o nitrogênio absorvido tenderia a reagir com os carboidratos do sistema radicular, beneficiando as raízes em detrimento da parte aérea. A partir do momento em que se aumenta a disponibilidade de nitrogênio, maior proporção desse nutriente é translocado para parte aérea, possibilitando o uso de carboidratos na síntese de proteínas e no crescimento. Contudo, uma menor proporção de carboidratos é translocado para o sistema radicular, de forma que se justifica o menor crescimento das raízes que o da parte aérea (Brower, 1962)
Quanto maior o valor do PMST, melhor a qualidade da muda avaliada, considerando que o acúmulo de biomassa é uma característica importante, relacionada ao crescimento da planta. Leles et al. (2000) atribuíram o maior crescimento inicial das mudas em campo ao maior crescimento da parte aérea e raiz verificados ainda em viveiro. Segundo Koslowski et al. (1991), o crescimento inicial das plantas no campo depende de fotossintatos armazenados pela muda.
Considerando os resultados apresentados, dentre as maiores doses de N aplicadas, a dose 219 mg . L -1 seria a mais indicada, visto que, ao realizar um teste de contraste entre as médias, as duas maiores doses foram estatisticamente iguais, optando-se, portanto, pela menor (Quadro 5).
3.1.2. Fertilização fosfatada
Na Figura 1 consta as características morfológicas altura, diâmetro de colo e peso de matéria seca total (PMST) em função das doses de P e os modelos ajustados para cada característica.
al., 1990). Outros trabalhos evidenciaram efeito positivo no crescimento de mudas de
espécies florestais quando da aplicação de fósforo em substrato, por exemplo, Novais et al. (1979) em Eucalyptus grandis e Dias et al. (1990) em Acacia mangium.
O crescimento em altura de mudas de eucalipto pode estar relacionado a outros fatores não nutricionais. Segundo Barros et al. (1978), tubetes pequenos (50 cm3 de capacidade) podem causar menor crescimento no viveiro, mas o crescimento em altura de Eucalyptus grandis é recuperado no campo. De qualquer forma, mesmo utilizando tubetes pequenos (50cm3) e sendo as mudas avaliadas apenas aos 42 dias de idade, as alturas encontradas nas doses intermediárias de P apresentaram-se satisfatórias, alcançando valores próximos a 19 cm (Figura 1). Guerreiro e Colli Junior (1984), ao estabelecer critérios na seleção de mudas, relataram para Eucalyptus urophylla e Eucalyptus saligna alturas oscilando entre 15 e 35 cm. Para Silveira et al. (2001),
mudas de eucalipto que atingem altura média de 20-25 cm já estão prontas para a fase de rustificação, fase que antecede o plantio das mesmas no campo.
O crescimento em diâmetro de colo foi semelhante ao encontrado para altura em função das doses de P, apresentando comportamento quadrático (Figura 1). Daniel et al. (1997) também encontraram maiores valores de diâmetro de colo com aplicação de maiores doses de P em substrato ao trabalhar com Acacia mangium em Latossolo Roxo Distrófico. Os autores sugeriram, para esta espécie, a aplicação de no máximo 175 mg.dm-3 de P se a intenção for produzir plantas com bom diâmetro de colo, não havendo a necessidade de se aplicar mais.
Ao contrário do observado no presente estudo, Gomes et al. (2003) verificaram em trabalho com E. grandis que a disponibilidade de fósforo não afetou nenhuma característica morfológica da muda. No entanto, os autores justificaram que o nível de fósforo no substrato (845 mg.dm-3), antes das adubações, já estava bem alto e, possivelmente, muito além dos críticos exigidos para o crescimento.
Loneragan et al. (1966), a queda na produção de matéria seca, no mais alto nível de fertilização, pode estar associada à toxicidade causada pelo fósforo. Este efeito foi observado por Furtini Neto et al. (1996) em mudas de várias espécies de Eucalyptus.
Y = 4,8741 * (4,2877 - e (-0,002821x)) R2 = 0,27 Y = 17,1721 + 0,006425x – 0,000007041x2 R2 = 0,25
Y = 0,05074 * (5,5494 – e (-0,003222x)) R2 = 0,27 Y = 0,242 + 0,0000657x – 0,0000000672 x2 R2 = 0,16
Y = 0,4441 * (2,7162 - e (-0,003464x)) R2 = 0,30 Y = 0,9252 + 0,0003937x – 0,0000004927x2 R2 = 0,17
Figura 1 – Valores de altura, diâmetro de colo e peso de matéria seca total (PMST) em mudas de eucalipto aos 42 dias de idade após estaqueamento, em função de doses de N e P (dados dos 3 clones). As barras representam o intervalo de confiança e os pontos correspondem às médias.
14,00 15,00 16,00 17,00 18,00 19,00 20,00 21,00 A ltur a (c m) 0,21 0,22 0,23 0,24 0,25 0,26 0,27 0,28 D iâ m et ro d e co lo ( c m ) 14,00 15,00 16,00 17,00 18,00 19,00 20,00 21,00 Al tu ra ( c m ) 0,21 0,22 0,23 0,24 0,25 0,26 0,27 0,28 D iâm et ro d e co lo (c m ) 0,60 0,70 0,80 0,90 1,00 1,10 1,20
Dose de N (mg/L)
PM
ST
(
g
)
0 73 146 219 292
0,60 0,70 0,80 0,90 1,00 1,10 1,20
Dose de P (mg/dm3)
PM
ST
(
g
)
Quadro 4 – Distribuição do peso de matéria seca das mudas aos 42 dias em função das doses de N e P para os três clones avaliados
Distribuição do Peso da Matéria Seca (%) Tratamentos
Clones Compartimento da muda
N1 N2 N3 N4 N5
Parte Aérea 67 68 70 72 73
386
Raiz 33 33 33 33 33
Parte Aérea 69 70 74 76 77
1213
Raiz 31 30 26 24 23
Parte Aérea 69 71 72 73 75
3837
Raiz 31 29 28 27 25
P1 P2 P3 P4
Parte Aérea 70 71 71 70
386
Raiz 30 29 29 30
Parte Aérea 73 73 73 73
1213
Raiz 27 27 27 27
Parte Aérea 72 73 73 72
3837
Raiz 28 27 27 28
Obs.1: N1 (0 mg.L-1); N2 (73 mg.L-1); N3 (146 mg.L-1); N4 (219 mg.L-1) e N5 (292 mg.L-1). Obs.2: P1 (0 mg.dm-3); P2 (349 mg.dm-3); P3 (698 mg.dm-3) e P4 (1046 mg.dm-3).
Quadro 5 – Grupo de médias de altura, diâmetro de colo e peso de matéria seca total (PMST), em função das doses de N, definidos pelo teste Tukey (α= 0,05).
Altura Diâmetro de colo PMST
Grupo Grupo Grupo
Dose de N
(mg . L-1) Média
(cm) 1 2 3 4
Média
(cm) 1 2 3 4
Média
(g) 1 2 3 4
0 16,0 a 0,230 a 0,76 a
73 17,0 a b 0,245 b 0,88 b
146 17,6 b c 0,250 b c 0,92 b c
219 18,3 c d 0,255 c d 1,01 c d
3.2. Características nutricionais
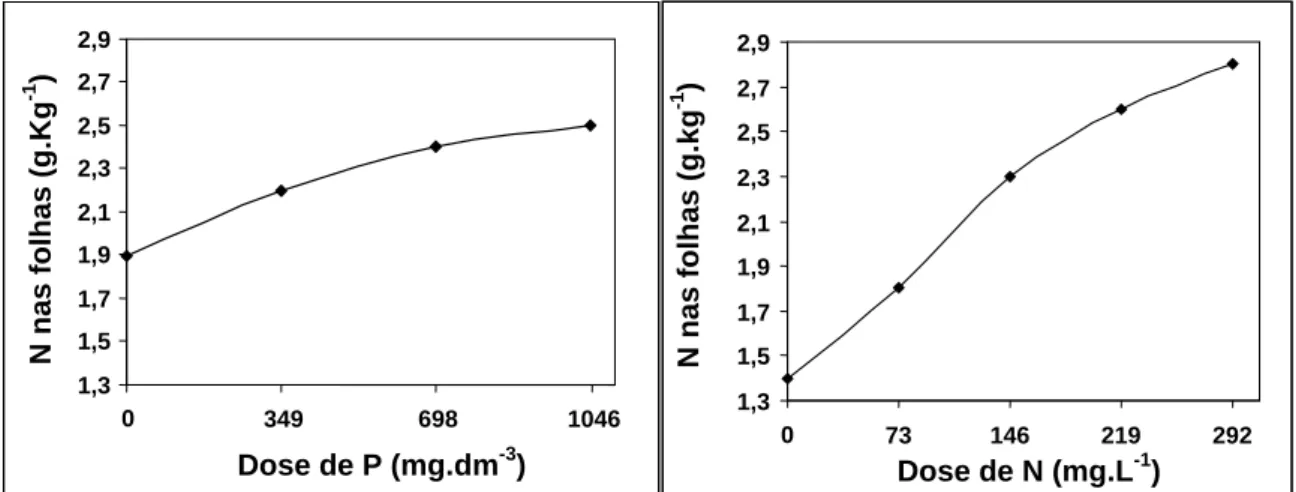
3.2.1. Concentração de N e P foliar
Maiores concentrações de N foliar foram encontradas nas maiores doses aplicadas, tanto de N quanto de P (Figura 2). As baixas concentrações de N foliar refletiram rapidamente na visualização dos sintomas de deficiência. No sétimo dia de permanência das mudas no estaleiro recebendo os tratamentos, já se observava nitidamente o “avermelhamento” das folhas no tratamento onde não se aplicou N. O mesmo foi relatado por Silveira et al. (2002) trabalhando com clones de Eucalyptus grandis x Eucalyptus urophylla aos 120 dias de idade, onde após 30 dias de ausência do
nutriente resultou no avermelhamento total das folhas mais velhas. Sintomas semelhantes foram encontrados por Kaul et al. (1970), Rocha Filho et al. (1978) e Dell et al. (1995).
A tendência de crescimento para o N foliar em função das aplicações de P foi a mesma observada por Neves et al. (1990). Os autores relataram que a resposta das mudas ao nitrogênio são dependentes, em grau variável, de um adequado suprimento de fósforo. Da mesma forma, Guimarães et al. (1959) observaram resposta positiva ao nitrogênio apenas quando o fósforo foi aplicado ao substrato de crescimento em mudas de E. saligna e E. camaldulensis.
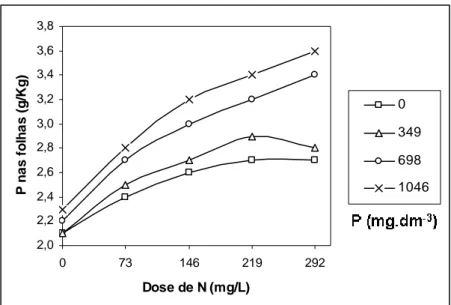
De acordo com a Figura 3, observa-se que a concentração de P nas folhas aumenta com o aumento da dose de N, porém esta tendência é limitada quando o P não foi aplicado em substrato (0 mg.dm-3) ou quando se aplicou 349 mg.dm-3. Estes resultados evidenciam a necessidade de uma quantidade ótima de fósforo a ser aplicada para que a fertilização nitrogenada possa atuar de forma plena na absorção do próprio P. Normalmente, verifica-se maiores valores de P foliar quando realizada a fertilização nitrogenada. Em plantas forrageiras, Noller e Rhykerd (1974) verificaram ao aplicar nitrogênio, maiores concentrações de fósforo na massa seca das plantas, sendo que, quando aplicado na forma de amônio (NH4+), os valores de P foliar foram maiores
do que quando aplicados na forma de nitrato (NO3-). O comportamento diferenciado
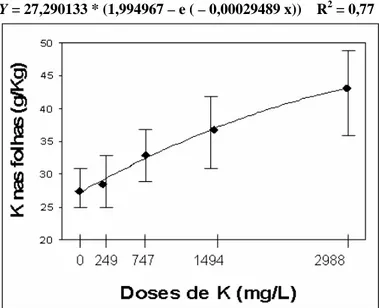
As duas maiores doses de P proporcionaram as maiores concentrações de P foliar e, ao mesmo tempo, um efeito depressivo nos valores de altura, diâmetro de colo e PMST. Segundo Furtini Neto et al. (1996), algumas espécies parecem não apresentar mecanismo efetivo para evitar “consumo de luxo” de P, levando à toxicidade. De acordo com Loneragan et al. (1966), se a concentração de P na matéria seca da parte aérea excede a 8 g.Kg-1, podem ser observados sintomas de toxicidade em certas plantas, enquanto outros trabalhos delimitaram esta concentração em 10 g.Kg-1 (Asher e Loneragan, 1967). No presente estudo, a concentração foliar de P alcançou 3,6 g.Kg-1 na maior dose de P aplicada, bem abaixo das concentrações consideradas como tóxicas. Para Herbert (1990), pode ser esperado nas fertilizações com fósforo, nenhum efeito ou efeito negativo nas plantas quando o status de nitrogênio em solo/substrato é baixo, devido ser a absorção de fósforo altamente influenciada pelo suprimento de nitrogênio.
Figura 2 – Concentração média de N foliar em função de doses de fósforo (P) aplicadas em substrato e de nitrogênio aplicadas na ferti-irrigação (dados dos três clones).
1,3 1,5 1,7 1,9 2,1 2,3 2,5 2,7 2,9
0 349 698 1046
Dose de P (mg.dm-3)
N nas folhas
(g.Kg
-1 )
1,3 1,5 1,7 1,9 2,1 2,3 2,5 2,7 2,9
0 73 146 219 292
Dose de N (mg.L-1)
N nas folhas
(g.kg
2,0 2,2 2,4 2,6 2,8 3,0 3,2 3,4 3,6 3,8
0 73 146 219 292
Dose de N (mg/L)
P
n
a
s f
o
lh
as
(
g
/K
g
)
0 349 698 1046
4. CONCLUSÕES
Com base nos resultados, concluiu-se para o presente ensaio:
a) O nitrogênio tem efeito positivo no crescimento e qualidade de mudas de eucalipto, sendo a dose 219 mg.L-1 de nitrogênio a recomendada durante a fase de crescimento para os três clones avaliados.
5. REFERÊNCIAS BIBLIOGRÁFICAS
ASHER, C. J.; LONERAGAN, J. F. Response of plants to phosphate concentration in solution culture: Growth and phosphorus content. Plant and soil, v.103, p.225-233, 1967.
BARROS, N. F.; BRANDI, R. M.; COUTO, L. Efeitos de recipientes na sobrevivência e no crescimento de mudas de Eucalyptus grandis no viveiro e no campo. Revista Árvore, Viçosa, MG, v. 2, n. 2, p. 141-151, 1978.
BARROS, N. F.; NOVAIS, R. F.; NEVES, J. C. L.; GOMES, J. M. Interpretação de análises químicas de solo para o crescimento de Eucalyptus spp. Revista Árvore, Viçosa, v. 6, n. 1, p. 38-44, 1982.
BROWER, R. Nutritive influences on the distribuition of dry matter in the plant. Netherlands in the plants. Netherlands Journal of Agricultural Science, v.10, n.5, p.399-408, 1962.
CURVE EXPERT. A curve fitting system for Windows. Disponível em <http://curveexpert.webhop.biz/>. Acesso em: 01 dez. 2007.
DANIEL, O.; VITORINO, A.C.T.; ALOVISI, A.A.; MAZZOCHIN, L.; TOKURA, A.M.; PINHEIRO, E.R. e SOUZA, E.F. Aplicação de fósforo em mudas de Acacia mangium WILLD. Revista Árvore, Viçosa, MG, v.21, n.2, p.163-168, 1997.
DEL QUIQUI, E. M.; MARTINS, S. S.; PINTO, J. C.; PARAZZI, P. J. A.; MUNIZ, A. S.; Crescimento e composição mineral de mudas de eucalipto cultivadas sob condições de diferentes fontes de fertilizantes. Acta Scientiarum. Agronomy Maringá, v.26, n.3, p.293-299, 2004.
DELL, B.; MALAJCZUK, N.; GROVE, T.S. Nutrients disorders in plantation
eucalypts.
Camberra: Australian Center for International Agriculture Research, 1995. 104 p.
DIAS, L. E.; ALVAREZ, V. H.; BRIENZA Jr., S. Formação de mudas de Acacia mangium: I. Resposta a calcário e fósforo. In: CONGRESSO FLORESTAL
BRASILEIRO, 6, 1990. Campos do Jordão. Anais… Campos do Jordão: SBS/SBEF, 1990.
EPSTEIN, E. Nutrição Mineral de Plantas: princípios e perspectivas. São Paulo: EDUSP, 1975. 341p.
GOMES, J. M. Parâmetros morfológicos na avaliação da qualidade de mudas de
Eucalyptus grandis, produzidas em diferentes tamanhos de tubetes e de dosagens
de N-P-K. 2001. 166f.. Tese (Doutorado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa, MG.
GOMES, J. M.; COUTO, L.; LEITE, H. G.; XAVIER, A.; Garcia, S. L. R. Crescimento de mudas de Eucalyptus grandis em diferentes tamanhos de tubetes e fertilização N-P-K. Revista Árvore, v.27, n.2, p.113-127, 2003.
GOMES, J. M.; PAIVA, H. P. Viveiros Florestais (propagação sexuada). Viçosa: UFV, 2004, 116p. (Caderno didático 72).
GRANT, C. A.; FLATEN, D. N.; TOMASIEWICZ, D. J.; SHEPPARD, S. C. A importância do fósforo no desenvolvimento inicial da planta. Informações agronômicas, Piracicaba, SP, n. 95, 16 p. , 2001.
GUERREIRO, C. A.; COLLI JUNIOR, G. Controle de qualidade de mudas de Eucaliptus spp. na Champion Papel e Celulose S. A. In: SIMPÓSIO
INTERNACIONAL: Métodos de Produção e controle de qualidade de sementes e mudas florestais,1984, Curitiba: UFPR/FUPEF, 1984, p.127-133.
GUIMARÃES, R. F.; GOMES, F. P.; MALAVOLTA, E. Adubação em “torrão paulista” de Eucalyptus saligna Sm. Companhia Paulista de Estradas de Ferro. 1959, 12 p. (Boletim 12).
HERBERT, M.A. Fertilizer site interactions on the growth and foliar nutrient levels of Eucalyptus grandis. Forest Ecology Manage, v.30, p.247–257, 1990.
HIGASHI E. N.; SILVEIRA, R. L. V. A.; GONÇALVES, A. N. Nutrição e adubação em minijardim clonal hidropônico de Eucalyptus. IPEF, 2002. (Circular técnica 194). KAUL, O. N.; SRIVASTAVA, P. B. L.; TANDON, V. N. Nutrition studies on Eucalyptus:
IV. diagnoses of mineral deficiencies in Eucalyptus globulus seedlings. Indian Forester, New Delhi, v. 96, n. 6, p. 453-456, June 1970.
KOZLOWSKI, T. T., KRAMER, P. J., PALLARDY, S. G. The physiological ecology of woody plants. New York: Academic Press, 1991. 657p.
LELES, P.S.S., CARNEIRO, J. G. A., BARROSO, D. G., MORGADO, I. F. Qualidade de mudas de Eucalyptus spp. produzidas em blocos prensados e em tubetes. Revista Árvore. v.24, n.1, p.13-20, 2000.
LONERAGAN, J. F.; CARROL, M. D.; SNOWBALL, K. Phosphorus toxicity in cereal crops. Journal of Australian Institute of Agricultural Science, v.32, p.221-223, 1966.
MARQUES, V. B.; PAIVA, H. N.; GOMES, J. M.; NEVES, J. C. L.; Efeitos de fontes e doses de nitrogênio no crescimento de mudas de sabiá (Mimosa caesalpiniaefolia Benth.). Scientia Forestalis, n. 7, p.77-85, agosto, 2006.
MOREIRA, F.M.S.; MOREIRA, F.W. Característica de germinação de 64 espécies de leguminosas florestais nativas da Amazônia, em condições de viveiro. Acta Amazônica, Manaus, v.26, n.1/2, p.3-16, 1996.
NEVES, J. C. L.; GOMES, J. M.; NOVAIS, R. F.; Fertilização mineral de mudas de eucalipto. In: BARROS, N. F.; NOVAIS, R. F. Relação solo-eucalipto. Viçosa, MG, 1990, p.100-124.
NOLLER, C. H.; RHYKERD, C. L. Relationship of nitrogen fertilization and chemical composition of forage to animal health and performance. In: MAYS, D. A. Forage fertilization. (Ed.). Madison: American Society of Agronomy, 1974. cap.17, p.363-387.
NOVAIS, R.F.; GOMES. J.M.; ROCHA, D. Calagem e adubação mineral na produção de mudas de eucalipto (Eucalyptus grandis W. HiII ex Maiden). Revista Árvore, Viçosa, v.3, p.121.134, 1979.
PAIVA, H. N. de; GOMES, J. M. Viveiros florestais. Viçosa, MG: UFV, 1993. 56p.
ROCHA, D.; BRAGA, J. M. Adubação fosfatada em eucaliptos no viveiro. 1. Interação entre espécies de eucaliptos e fontes de fósforo. In: CONGRESSO FLORESTAL BRASILEIRO, 4., 1982, Belo Horizonte. Anais... Belo Horizonte: 1982. p.455-459.
ROCHA FILHO, J. V.; HAAG, H. P.; OLIVEIRA. G. D. Deficiência de macronutrientes,
boro e ferro em Eucalyptus urophylla. Anais da ESALQ, Piracicaba, v. 35, n. 1, p. 19-34,
1978.
SILVA, M. R.; KLAR, A. E.; PASSOS, J. R. Efeitos do manejo hídrico e da aplicação de potássio nas características morfofisiológicas de mudas de Eucalyptus grandis W. (Hill ex. Maiden). Irriga, Botucatu, SP, v.9, n.1, p.31-40, jan./abril 2004.
SILVEIRA, R. L. V. A.; HIGASHI, E. N.; SGARBI, F.; MUNIZ, M. R. A. Seja o doutor do seu eucalipto. Informações Agronômicas, n.93, p.1-32, março 2001 (Potafós – encarte técnico).
SILVEIRA, R. L V. A.; MOREIRA, A.; TAKASHI, E. N.; SGARBI, F; BRANCO, E. F. Sintomas de deficiência de macronutrientes e de boro em clones híbridos de
eucalyptus grandis com eucalyptus urophylla. Cerne, Piracicaba, SP, v. 8, n.2, p.107-116, 2002.
STATSOFT INC. Statistica data analysis system version 8.0. Tulsa: Statsoft Inc.,
CAPÍTULO 2 – EFEITO DO POTÁSSIO NA FASE DE RUSTIFICAÇÃO DE MUDAS CLONAIS DE EUCALIPTO
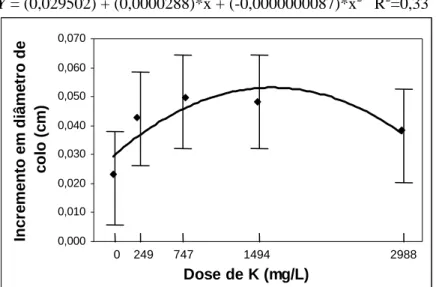
RESUMO
Dentro da questão de qualidade e nutrição de mudas de eucalipto, a aplicação de K está muitas vezes relacionada ao processo de rustificação, que tem por objetivo adaptar a muda às condições de plantio no campo. Neste sentido, este experimento foi conduzido visando avaliar o efeito do potássio na qualidade de mudas de três clones de Eucalyptus spp. O experimento foi instalado no viveiro de mudas da empresa