FIDELIDADE À ÁREA E PADRÃO DE ASSOCIAÇÃO EM SOTALIA GUIANENSIS, BASEADO NA TÉCNICA DE FOTO-IDENTIFICAÇÃO
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre do Programa de Pós-Graduação em Psicobiologia.
FIDELIDADE À ÁREA E PADRÃO DE ASSOCIAÇÃO EM SOTALIA GUIANENSIS, BASEADO NA TÉCNICA DE FOTO-IDENTIFICAÇÃO
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre do Programa de Pós-Graduação em Psicobiologia.
Orientadora: Prof.ª Maria Emília Yamamoto (emiliayamamoto@gmail.com).
Ananias, Sandra Mara de Araújo.
Fidelidade à área e padrão de associação em Sotalia guianensis,
baseado na técnica de foto-identificação / Sandra Mara de Araújo Ananias. – Natal, RN, 2006.
37 f.
Orientadora: Maria Emília Yamamoto.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-graduação em Psicobiologia.
1. Sotalia guianensis – Dissertação. 2. Foto-identificação –
Dissertação. 3. Padrão de associação – Dissertação. 4. Pipa (RN) – Dissertação. I. Yamamoto, Maria Emília. II. Universidade Federal do Rio Grande do Norte. III. Título.
“No fundo do meu poço, tem molas.”
AGRADECIMENTOS
RESUMO
ABSTRACT
The behavioral patterns follow to environmental changes, including area fidelity and individuals association patterns. Several techniques are used to record these behavioral patterns and the photo-identification has been suggested as a proper tool because of its various advantages. Based on this technique, this research verified, between August of 2005 and January of 2006, area fidelity and association patterns of Sotalia guianensis, at Distrito de Pipa’s bays, Rio Grande do Norte State south coast. Besides, we measured the association patterns by using the Jaccard index or “Half-Weight Index” (HWI). According the observation, 22 individuals were not resighted, 11 were resighted, and 36 new individuals were recorded. Nowadays, 69 individuals are cataloged. The residency rate indicated heterogeneity on studied area permanence and the association patterns between photo-identified seem to be context-specific. In addiction, the comparison of associations between two different age classes showed some individuals more frequently interacting with immature individuals. We also observed fluidity on association patterns among our individuals. We suggest that S.
SUMÁRIO
EPÍGRAFE...iv
AGRADECIMENTOS...v
RESUMO...vi
ABSTRACT...vii
LISTA DE ILUSTRAÇÕES...ix
1 1.. INTRODUÇÃO...1
2 2.. OBJETIVOS...7
3 3.. MATERIAIS E MÉTODO...8
4 4.. RESULTADOS E DISCUSSÃO...18
5 5.. CONCLUSÕES...29
6 6.. REFERÊNCIAS...30
LISTA DE ILUSTRAÇÕES
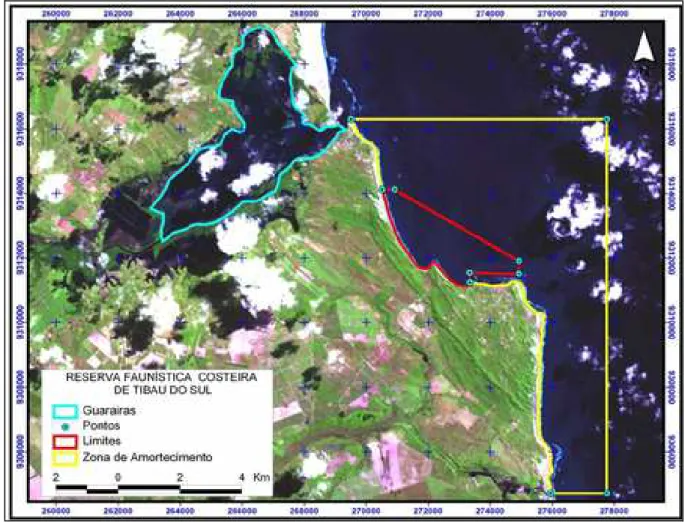
Figura 1 - Foto de satélite da área de estudo com respectivos limites da REFAUTS. Em vermelho, Zona de Uso Restrito. ...8
Figura 2 - Foto aérea da ligação direta da Laguna de Guaraíras com o mar (Foto: R. Matos)...9
Figura 3 - Espécime de boto cinza, Sotalia guianensis (Foto: P. I. A. P. Medeiros). ...10
Figura 4 - Mapa geral da distribuição para Sotalia guianensis. Adaptado de Nascimento (2002). ...11
Figura 5 - Barcos de turismo utilizados para obtenção dos dados utilizados neste estudo (Fonte: S. M. A. Ananias). ...12
Figura 6 - Desenho esquemático do percurso realizado pelas embarcações de turismo, traçado sobre uma foto aérea da região de estudo (Foto: R. Matos)...13
Figura 7 - Exemplo de espécimes de S. guianensis formando grupo coeso (Fonte: S. M. A. Ananias)...14
Figura 8 - Exemplo de subgrupos próximos de S. guianensis (Fonte: S. M. A. Ananias)...14
Figura 9 - Exemplos de fotografias obtidas e suas respectivas categorias: (A) excelente, (B) boa, (C) baixa e (D) comportamento (Fonte: S. M. A. Ananias). ...15
Figura 10 - Contorno da nadadeira dorsal do animal foto-identificado S60, obtido através do programa computacional CorelDRAW Graphics Suite 12 (Fonte: S. M. A. Ananias). ...16
Figura 11 - Freqüência absoluta de avistagem dos indivíduos foto-identificados considerados residentes ao longo do período de estudo. ...23
Figura 12 - Permanência temporal dos indivíduos foto-identificados considerados residentes durante o período de estudo. ...24
Tabela II - Total de indivíduos por número de avistagem ao longo do período de observação deste estudo. Os indivíduos que aparecem em negrito foram identificados em estudos anteriores e os restantes neste (ver nota). ...20
Tabela III - Índice de associação (Jaccard), freqüências observada e esperada para as possíveis díades durante o período de observação deste estudo. Os indivíduos adultos são representados por “A” e os imaturos por “I”..23
Tabela IV - Índice de associação (Jaccard) encontrado para os indivíduos foto-identificados (ID). ...27
Tabela V - Índice de associação (Jaccard) calculado, individualmente, entre L3 e S20 e entre estes e as diferentes faixas etárias. Os indivíduos adultos são representados por “A”, imaturos por “I” e a díade adulto/imaturo por “AI”. ...28
ANEXO A
1. INTRODUÇÃO
Sabemos que os organismos não se adaptam passivamente. Eles modificam e regulam ativamente o ambiente físico, dentro de limites impostos pelas leis naturais que determinam a transformação de energia e a ciclagem de materiais. A interação entre os sistemas genéticos e o ambiente determina o curso da seleção natural e, portanto, não apenas a forma como os organismos otimizam a sua sobrevivência, mas também a maneira pela qual os ecossistemas se modificaram e estão se modificando ao longo do tempo evolutivo (Odum, 1988).
Mesmo quando a comunidade e o ecossistema parecem não mudar, a densidade, a natalidade, a mortalidade, o potencial biótico (ou potencial reprodutivo), a dispersão, a sobrevivência, a estrutura etária, a taxa de crescimento e outros atributos das populações componentes geralmente flutuam à medida que as espécies se ajustam às forças físicas e umas às outras (Odum, 1988). Dessa forma, pode-se afirmar que as populações apresentam um comportamento dinâmico, mudando continuamente no tempo e no espaço (Ricklefs, 2003), e que essas mudanças podem fornecer um retrato das interações comunidade-ecossistema.
Dessa maneira, observar na íntegra a riqueza do repertório comportamental de uma espécie e entender as condições às quais cada atividade está adaptada, pode: (a) revelar aspectos do seu comportamento que não se conheceria de outra maneira e (b) proporcionar a compreensão de como um determinado comportamento do animal está adaptado ao ambiente em que vive (Martin Bateson, 1993).
No entanto, a estimativa de parâmetros populacionais a partir do estudo do comportamento pode sofrer variações entre as espécies em função, sobretudo, da complexidade de seu repertório comportamental. Em mamíferos, por exemplo, grandes problemas para a estimativa de parâmetros populacionais têm sido apontados, atribuídos especialmente à complexidade comportamental desta classe (Fernandez, 1995). Outra limitação para a investigação da estimativa populacional é o estudo do comportamento em ambiente natural.
dependendo este balanço, da espécie a ser estudada e das condições na qual esta vive (Martin Bateson, 1993). Estudos com cetáceos caracterizam-se como exemplos dessa dificuldade, pois segundo Lodi (2002), na maioria dos representantes dessa ordem não há dimorfismo sexual e durante a respiração, esses mamíferos vêm à tona por curtos intervalos de tempo, expondo apenas cerca de um terço da superfície corporal, passando a maior parte do tempo submerso.
Devido às dificuldades acima citadas, a obtenção dos dados ecológicos para esses animais torna-se um processo lento. No entanto, deve-se destacar a identificação dos indivíduos como um aspecto chave para o estudo de estimativas populacionais. Esse processo garante que cada animal seja contado uma única vez, permitindo que se evite a recontagem de um animal, o que pode ocasionar uma superestimação da população a ser estudada.
Embora nas décadas de 50 e 60, marcas artificiais tenham sido consideradas como pré-requisito para pesquisas enfocando o comportamento, o aumento no número de estudos de longo prazo de animais de vida livre, tem mostrado que vertebrados podem usualmente ser identificados por marcas naturais, fazendo da identificação individual a principal ferramenta das pesquisas de campo (Würsig & Jefferson, 1990).
Diversas técnicas alternativas de amostragem têm sido utilizadas. Para grandes felinos, por exemplo, é comum a tentativa de identificação de indivíduos através de particularidades das pegadas deixadas em determinada área (e.g. Miller, 2001). Outra abordagem é a obtenção de estimativas baseando-se em contagens de pilhas de fezes, entretanto essa técnica é pouco consistente, uma vez que o grau de incerteza é bastante elevado (Tomas & Miranda, 2004). Métodos cada vez mais sofisticados e valiosos têm sido desenvolvidos, resolvendo vários problemas quanto à amostragem para a estimativa, entre eles destacam-se: (a) a radiotelemetria, (b) os marcadores moleculares e (c) o armadilhamento fotográfico (Fernandez, 1995; Tomas & Miranda, 2004).
ferramenta que pode trazer várias informações sobre diversas populações a serem estudadas (Acuña, 2002).
1.1. FOTO-IDENTIFICAÇÃO
Essa técnica se baseia na identificação e reconhecimento individuais, de baleias e golfinhos, usando fotografias de marcas naturais (Whitehead, Christal & Tyack, 2000), sendo recomendáveis somente marcas naturais que se mantêm reconhecíveis ao longo do período de estudo e que são individualmente distintas (Würsig & Jefferson, 1990).
A técnica de foto-identificação foi desenvolvida para a utilização em estudos com cetáceos na década de 70, quando as primeiras pesquisas descreveram a ocorrência de entalhes (obtidos naturalmente) na nadadeira dorsal de golfinhos nariz-de-garrafa (Tursiops truncatus), auxiliando na identificação individual desses animais (Würsig & Würsig, 1977). Segundo Santos (1999), desde o final da década de 80 a foto-identificação passou a ser aplicada para o melhor conhecimento dos cetáceos no Brasil, sendo alvo de diversos estudos ecológicos que se utilizam dessa técnica. Somente a partir dos anos 90, começaram a se estabelecer bancos de dados que se utilizavam dessa ferramenta, possibilitando a realização de estudos de longo prazo (Whiteheadet al., 2000).
Nos últimos anos, o uso da foto-identificação por cetólogos tem apresentado inúmeras vantagens em relação à utilização de marcas artificiais, pois é uma técnica que pode ser usada como captura-recaptura que vem auxiliando os estudos de cetáceos em seu ambiente natural, sem interferir de forma invasiva na vida desses animais (Würsig & Jefferson, 1990).
A aplicabilidade da foto-identificação para estudos com Sotalia guianensis foi descrita por Flores (1999), e posteriormente aplicada por vários pesquisadores, apesar das dificuldades inerentes ao comportamento, pequeno porte e dimorfismo sexual não evidente dessa espécie.
biológico. No entanto, essas características são de interesse e relevância consideráveis, principalmente porque variações biologicamente significativas ocorrem muito freqüentemente e são essenciais na identificação individual.
Dessa maneira, o reconhecimento de indivíduos pode ser utilizado como subsídio para uma grande variedade de informações da história natural da espécie. Talvez o uso mais comum das técnicas de identificação para cetáceos tenha sido para composição de grupo e fidelidade de certos animais ao grupo. Porém, técnicas de captura-recaptura podem ser aplicadas para se obter uma estimativa do tamanho populacional (Würsig & Jefferson, 1990).
Embora a espécie apresente uma ampla distribuição, o conhecimento sobre diversos aspectos relacionados à dinâmica populacional de S. guianensis ainda é escasso (Santos, 1999).
1.2. ESTRUTURASOCIAL
O comportamento social inclui todos os tipos de interações entre co-específicos, da cooperação ao antagonismo. Cada interação exige que o indivíduo perceba o comportamento dos outros e dê as respostas apropriadas. A seleção imposta pelas interações com membros familiares e com indivíduos não aparentados numa população proporciona a base para a modificação evolutiva do comportamento social (Ricklefs, 2003).
Ainda assim, muitos estudos têm mostrado que animais que vivem em grupo obtêm benefícios através (1) do efeito diluição, com a diminuição do risco de ser capturado; (2) da detecção precoce do perigo; (3) da repulsão do inimigo de forma mais eficiente; (4) da manutenção da temperatura corporal, quando os animais são pequenos, e ainda (5) da facilitação social (Alcock, 2001).
Alterações intra-específicas de padrões sociais, tais como associações, estabilidade e composição de grupo e estratégias reprodutivas têm sido interpretadas como uma representação de respostas adaptativas a variações de parâmetros ambientais (Quintana-Rizzo & Wells, 2001). Claramente, o ambiente social e familiar de um indivíduo, junto com suas relações com membros do sexo oposto, impõe uma forte seleção sobre o comportamento e, indiretamente, sobre a história de vida e as relações ecológicas (Ricklefs, 2003).
Assim como os elefantes e a maioria dos primatas, muitos odontocetos (cetáceos com dentes) levam uma vida social complexa (Connor, Mann, Tyack & Whitehead, 1998). Desde a década de 70, pesquisas com o objetivo de entender a organização social em cetáceos (e.g. Würsig, 1978) vem sendo realizadas. Estudos com golfinhos nariz-de-garrafa, Tursiops truncatus, constataram que as associações são complexas em virtude dos padrões de movimento encontrados, como migração e residência (Quintana-Rizzo & Wells, 2001). Já para golfinhos de Hector,
Cephalorhynchus hectori, os padrões de associação foram caracterizados como
fluidos, com duração de não mais do que poucos dias (Bräger, 1999; Slooten, Dawson & Whitehead, 1993).
Dessa forma, a reavistagem de indivíduos a fim de avaliar os padrões de associação compreende uma ferramenta importante para o entendimento de mudanças na organização social de uma população (Quintana-Rizzo & Wells, 2001).
Índices de associação foram primeiramente desenvolvidos para descrever espécies que co-habitavam um mesmo ambiente ou nicho ecológico, mas recentemente e de modo crescente, têm sido utilizados para medir o grau de associação entre indivíduos, especialmente entre vertebrados sociais (Bejder, Fletchert & Bräger, 1998). Esses índices também vêm sendo largamente utilizados em pesquisas referentes à estrutura social de cetáceos (e.g. Bejder, Fletchert & Bräger, 1998; Sartorio, 2005; Whitehead, 1997).
2. OBJETIVOS
2.1. OBJETIVOGERAL
Verificar a fidelidade à área e o padrão de associação apresentados por indivíduos da espécie Sotalia guianensis, nas enseadas pertencentes ao Distrito de Pipa, litoral sul do Estado do Rio Grande do Norte.
2.2. OBJETIVOSESPECÍFICOS
2.2.1. Avaliar a fidelidade dos animais reconhecidos, que utilizam a área de estudo, através da técnica de captura-recaptura por foto-identificação.
2.2.2. Especificar o padrão de associação utilizado pelos animais individualmente distintos durante o período de estudo.
3. MATERIAL E MÉTODO
3.1. ÁREA DE ESTUDO
A área de estudo compreende as Enseadas do Curral e do Madeiro e adjacências, no litoral sul do Estado do Rio Grande do Norte, distrito de Pipa, pertencente ao município de Tibau do Sul, distando 85km da capital Natal. Encontra-se inEncontra-serida na Zona de Uso Restrito da ReEncontra-serva de Fauna Costeira de Tibau do Sul (REFAUTS) (Fig.1).
Apresenta uma topografia subaquática com declive suave e extenso, constituída por fundo arenoso com a presença de poucas rochas (Link, 2000). Além disso, é um ambiente bastante influenciado pelo complexo lagunar Guaraíras situado em Tibau do Sul, medindo 7km de comprimento e largura variando entre 1,4 e 2km; caracterizado pela alternância entre lagoa e laguna, com esta última, recebendo grande influência marinha, pois tem ampla comunicação com o mar (Sartorio, 2005) (Fig. 2).
Figura 2 – Foto aérea da ligação direta da Laguna de Guaraíras com o mar (Foto: R. Matos).
3.2. OBJETO DE ESTUDO
Sotalia guianensis (Cunha et al., 2005), popularmente denominado boto-cinza
Figura 3 – Espécime de boto-cinza, Sotalia guianensis(Foto: P. I. A. P. Medeiros).
Em geral, os odontocetos não realizam migrações extensas e regulares, como observada na maioria dos misticetos (baleias verdadeiras). Seus deslocamentos estão relacionados a movimentos de aproximação e distanciamento da linha da costa, podendo variar sazonalmente, em função da temperatura da água, dispersão e densidade espacial e temporal de recursos disponíveis (di Beneditto, Ramos Lima, 2001). Portanto, a espécie em questão, que pode ser encontrada regularmente freqüentando certas praias e baías ao longo de todo o litoral brasileiro, tem sido estudada em diferentes ecossistemas (Hetzel & Lodi, 1993).
A Enseada de Barra de Tabatinga no município de Nísia Floresta, e as Enseadas do Madeiro e do Curral no município de Tibau do Sul são as principais áreas de concentração de S. guianensis no litoral sul do Estado do Rio Grande do Norte, onde indivíduos dessa espécie podem ser observados durante todo o ano (Link, 2000). Corroborando com a referida autora, Sartorio (2005) verificou, ainda, a utilização do complexo lagunar Guaraíras pela espécie objeto deste estudo.
em estudos anteriores (e.g. Geise, 1989) e ao contrário do que proposto por Hetzel & Lodi (1993).
3.3. COLETA DOS DADOS
3.3.1 EQUIPAMENTOS
Para a realização da foto-identificação, foram utilizados filmes SUPERIA 400 ASA em uma câmera fotográfica reflex com lente zoom de 80 a 300mm, variando a velocidade do obturador entre 1/1000s e 1/4000s. Para o registro do número de grupos, número de indivíduos por grupo, faixas etárias e tipos de associação que ocorriam entre os indivíduos, foi utilizado um mini-gravador portátil.
3.3.2. PROCEDIMENTOS
As observações e fotos foram obtidas a partir de barcos de turismo, que transitam entre as enseadas do Curral e do Madeiro e áreas adjacentes, sempre próximos da costa (Figs. 5 e 6), entre os meses de Agosto/2005 e Janeiro/2006.
Figura 5– Barcos de turismo utilizados para obtenção dos dados utilizados neste estudo (Foto: S. M. A. Ananias).
Figura 6– Desenho esquemático do percurso realizado pelas embarcações de turismo, traçado sobre uma foto aérea da região de estudo (Foto: R. Matos, Adaptação: M. Gondim). Indica o início e o término do percurso.
As seguintes categorias foram definidas e adotadas:
ƒ Grupo de animais: todos os animais que estivessem utilizando ao mesmo tempo uma mesma área (dentro ou fora das enseadas).
ƒ Faixas etárias: adultos
imaturos: animais com no máximo 2/3 do tamanho do adulto. ƒ Critérios de associação:
- coesos: indivíduos que permaneciam a menos de um metro de distância uns dos outros, participando da mesma atividade comportamental e, por vezes, emergindo sincronicamente (Fig. 7).
Figura 7 – Exemplo de espécimes de S. guianensis formando grupo coeso (Foto: S. M. A. Ananias).
Figura 8 – Exemplo de subgrupos próximos de S. guianensis(Foto: S. M. A. Ananias).
3.3.2.1. ANÁLISE DAS FOTOGRAFIAS
adequadas para evidenciar a presença de qualquer marca permanente; (c) baixa qualidade – fotos que não permitiram a identificação de quaisquer tipos de marca, nem o contorno da nadadeira dorsal; e (d) comportamentos – fotos que registraram comportamentos diversos, mas que não possibilitavam a identificação de quaisquer tipos de marca, nem o contorno da nadadeira dorsal (Fig. 9).
(B) (A)
(C) (D)
Figura 9– Exemplos de fotografias obtidas e suas respectivas categorias: (A) excelente, (B) boa, (C) baixa e (D) comportamento (Fotos: S. M. A. Ananias).
Figura 10– Contorno da nadadeira dorsal do animal foto-identificado S60, obtido através do programa computacional CorelDRAW Graphics Suite 12 (Foto: S. M
.3.2.2. TAXA DE RESIDÊNCIA
Seguindo procedimento utilizado em pesquisas atuais que abordam padrões de residên
axa de residência = número de meses em que o animal foi foto-identificado x 100
.3.2.3. PADRÕES DE ASSOCIAÇÃO
Utilizou-se o índice de similaridade de Jaccard ou “Half-Weight Index” (HWI) para cal
diferentes faixas etárias anteriormente definidas.
. A. Ananias).
3
cia (e.g. Rossi-Santos, Wedekin e Monteiro-Filho, 2006; Hardt, Cremer, Simões-Lopes e Tonello-Júnior, 2006), no presente estudo as taxas de residência foram analisadas considerando-se a presença e ausência temporal dos indivíduos identificados na área de estudo; e calculada a partir da equação abaixo:
T
número total de meses
3
O índice de similaridade de Jaccard é calculado pela equação:
HWIij: aij
aij + bix + cjy
onde: aij é o número de perío os quais i e j são avistados
ntos; bix é o número de episódios nos quais i é avistado associado com quaisquer
e 0, menor a associação entre os anim
.3.2.3. ANÁLISEESTATÍSTICA
não paramétrico qui-quadrado (Ȥ2) para investigar
ossíveis diferenças nas associações predominantes dos indivíduos foto-identificados dos observados durante
ju
outros indivíduos (x), menos j; e, cjy é o número de observações nas quais j é avistado
associado com quaisquer outros indivíduos (y), menos i.
Este método determina valores que variam de 0 e 1, dependendo da proximidade entre os indivíduos: quanto mais próximo d
ais; e quanto mais próximo de 1, maior essa associação.
3
Aplicou-se o teste estatístico p
4. RESULTADOS E DISCUSSÃO
.1. FOTO-IDENTIFICAÇÃO
abalho inédito na região de Pipa e utilizando-se da técnica e foto-identificação, iniciou a composição de um catálogo dos botos avistados, no qual fo
dividualmente distint
5 dias de observação, totalizando 88 saíd
boa qualidade), perfazendo um total de 31,89% de aprove
nte a cada mês do período de estudo, com respectivas
Período/2005 Qualidade das Fotografias obtidas Total de Fotografias
4
Link (2000), em um tr d
i possível o registro de 18 indivíduos, baseando-se principalmente em marcas nas nadadeiras dorsais dos mesmos. Continuamente à conclusão desse trabalho, a autora ainda pôde reconhecer mais 9 animais, na mesma região, até o ano de 2002, totalizando 27 animais (L. O. Link, comunicação pessoal, 15/03/2002).
Dando prosseguimento a essa pesquisa, Ananias (2004) fez uma releitura do catálogo iniciado pela autora acima citada, totalizando 18 animais in
os; acrescentando-se ainda 16 novos registros.
Durante o período deste estudo, que compreendeu 6 meses de esforço, entre Agosto de 2005 e Janeiro de 2006, foram realizados 5
as de barco, em média 1,6 saídas ao dia; sendo possível contabilizar um total de 509 avistagens, entre animais identificados e não-identificados, com uma média de 5,78 indivíduos por saída.
Além disso, pôde-se obter um total de 279 fotografias. Dessas, 89 foram analisadas (excelente e
itamento da amostra (Tabela I).
Tabela I – Aproveitamento fotográfico refere qualidades das fotografias obtidas.
excelente boa baixa comportamentos
Agosto 18 41 14 73
Setembro 3
utubro
13 50 17
ro
83 3
O 3 13 15 3 4
Novemb 10 13 1 24
Dezembro 7 20 21 13 61
Janeiro
1
2 2 4
Total 3 76 142 48 279
Comparando-se o núm obtida cada mês de observação, odemos verificar que nos dois primeiros meses, houve a aquisição de uma maior quantidade de fotos de baixa qualidade (ver dados em negrito na Tabela I). Isso pode ser explicado devido ao aumento na incidência de ventos na região, que vai de meados de Julho até meados de Outubro, fazendo com que o mar fique com ondas maiores, comprometendo a estabilidade da embarcação e do pesquisador, dificultando a avistagem dos animais e a obtenção de melhores registros fotográficos.
Entretanto, esse elevado número de fotos de baixa qualidade não implicou, de uma forma geral, em um baixo índice de reconhecimento dos animais já inseridos no catálogo de foto-identificação, ou a identificação de novos espécimes.
Do mesmo modo que foi realizado por Ananias (2004), para que fossem acrescentados novos indivíduos ao catálogo, as imagens obtidas durante a coleta de dados do presente estudo foram comparadas tanto com os dados de Link (2000), como com os de Ananias (2004); verificando-se, assim, a presença de um falso positivo inserido no catálogo. Além disso, com a inserção dos novos registros, optou-se por modificar a nomenclatura dos indivíduos reconhecidos pela última pesquisadora – de “I” para “S” – para: (a) não confundir o código do animal com qualquer faixa etária e, (b) identificar somente através da inicial do espécime foto-identificado, qual pesquisador teria feito esse registro.
Dessa maneira, a partir da aplicação da técnica de foto-identificação, foram diferenciados 69 indivíduos, acrescentando ao catálogo 36 novos registros (Anexo A).
Do total dos indivíduos foto-identificados, 22 animais não foram avistados durante o período de estudo; 35 foram vistos uma única vez; 6 indivíduos em duas vezes; 2 por três vezes; 1 em quatro vezes e 1 em cinco diferentes vezes. Freqüências de avistagem maiores que cinco foram observadas apenas para 2 indivíduos, com 29 ocasiões para L3 e 11 para S20 (Tabela II).
Apesar de diversos estudos utilizarem o formato da nadadeira dorsal, o padrão de coloração e cicatrizes ao longo do dorso do animal (e.g. Flores, 1999; Pizzorno, 1999; Santos, 1999), no presente trabalho, o principal tipo de marca usado para diferenciar os indivíduos foi o contorno da nadadeira dorsal, que na maioria dos casos apresentava cortes. Além disso, como era de fácil percepção a diferença entre as
marcas
Tabela II – Total de indivíduos por número de avistagem ao longo do período de observação deste encontradas nos animais, não foi necessária a aplicação da equação da razão da nadadeira dorsal sugerida por Defran, Schultz e Weller (1990).
estudo. Os indivíduos que aparecem em negrito foram identificados em estudos anteriores e os restantes neste (ver nota).
Nº. de
Avistagens IndivíduosTotal de Indivíduos Identificados 0 22 L4, L5, L6, L7, L8, L9, L10, L14, L15, L16, L17
S23, S26, S27, S28, S29, S30, S31, S32 , S19, S21, S22,
1
2
35 L1, L11, L12, S24, S25, S35, S37, S38, S39, S40, S41, S42, S44, S45, S46, S47, S50, S51, S52, S53, S54, S55, S56, S57, S58, S59, S60, S61, S62, S63, S64, S65, S66, S67, S69
6 L13, L18, S33, S36, S43, S68
3 2 S48, S49
4 1 L2
5 3 L3, S20, S34
Nota: Os indivíduos em negrito foram foto-identificados e catalogados em estudos anteriores, por Link (2000; identificados por um L antes do número) e Ananias (2004; identificados por um S antes do número).
Constatou-se neste trabalho a presença de espécimes de S. guianensis durante todos os dias de observação. Durante o período de pesquisa, 50,72% dos indivíduos foto-identificados foram avistados uma única
identificados de
mesma área de estudo, durante nove meses.
ero de animais catalogados, ainda assim a quantidade de indiví
obtida por Link (2000). É interessante pontuar que do grupo dos animais reavistados
lagunar de Cananéia/SP; e, Flores (1999) que verificou 86% de reavistagem na Baía vez. Já para a reavistagem, esse valor alcançou 17,39% (n=12) do total. Link (2000) obteve para 18 espécimes
foto-S. guianensis, 74% (n=13) de reavistagem realizando observações na
A diferença entre os percentuais de reavistagem alcançado no presente trabalho e o obtido por Link (2000), ocorreu em função do aumento na obtenção de novos registros. Considerando-se que há um maior núm
duos reavistados observada neste estudo é similar àquela
nessa pesquisa, sete deles foram registrados em ambos os estudos, correspondendo a um indicativo de fidelidade à área.
Norte/SC; os altos índices de reavistagem de botos cinza encontrados podem ter sido obtidos em virtude do ambiente ser mais protegido nessas diferentes regiões.
Ainda assim, verificou-se que sete dos animais foto-identificados nesse estudo
animais que não foram avistados, nem
f s, e capturados, não estavam utilizando a área.
nias (20 basea indivíduos foto-ide ficado nos quais os animais foram
entre 1 e 2 meses; ocasionais, de 3 a 5 meses; freqüentes, de 6 a 7 meses; e comuns,
reavistados por 8 ou mais me ação, adaptou os padrões de
resid ia proposto por Z 002), a saber: (1) residentes, aqueles indivíduos que estavam inseridos nas categorias acima citadas de comuns e freqüentes; (2) residentes sazonais, aqueles classificados como ocasionais; e, (3) transientes, para os
tanto, para que fosse possível a utilização desses parâmetros temporais, seria necessário um período mínimo de oito meses de acompanhamento dos animais individualm
Dessa maneira, o tempo de permanência calculado para os animais que de observação foi: 33% para L18 e S68; 50% para S49; 67% para L2 e S20; 83% para S34; e 100% para L3.
identificados apresentaram valores baixos (16,67%) para esse índice. Ao contrário do itonga, litoral norte de Santa utilizaram a área por pelo menos dois meses do período de observação (Figs. 11 e 12). Contudo, isso não significa que aqueles
otografado ntre os meses que foram
Ana 04) ndo-se em Quintana-Rizzo & Wells (2001), classificou os nti s de acordo com o número de meses do período de estudo reavistados, sendo considerados: raros, aqueles reavistados
ses. A partir dessa categoriz
ênc s olman (2
que foram incluídos na categoria raros ou não foram novamente avistados. Entre
ente distintos. Em virtude do tempo de coleta de dados, optou-se pelo método utilizado por Rossi-Santos, Wedekin e Monteiro-Filho (2006) e Hardt et al. (2006), no qual os autores analisaram as taxas de residência através da divisão do número de reavistagens pelo número de meses de esforço, multiplicando-se por cem.
utilizaram a área de estudo com base no intervalo total
Como observado por Rossi-Santos, Wedekin e Monteiro-Filho (2006), na região do estuário do Rio Caravelas, no Estado da Bahia, as taxas de residência alcançadas para os animais amostrados na região de Pipa também demonstraram heterogeneidade de permanência, visto que cerca de 50,72% dos animais
Catarina, utilizando o mesmo método de análise, cuja parcela da amostra considerada residente foi de 37,2%.
Apesar de terem sido encontrados baixos valores referentes à permanência dos animais foto-identificados na área de estudo, o Projeto Pequenos Cetáceos da Universidade Federal do Rio Grande do Norte, a partir da foto-identificação do indivíduo L3 em Agosto de 1999 por Link (2000), vem obtendo dados relevantes que podem melhores resulta o durant ntre o barco
marinha concentra-se perto do litoral, onde as condições são mais favoráveis devido ao contribuir para a construção da história natural desse espécime, em virtude do mesmo ser continuamente acompanhado nas enseadas da região de Pipa, município de Tibau do Sul (Ananias, 2004; Jesus, 2004; Nascimento, 2002; Spinelli, 2004), bem como na enseada formada na praia de Barra de Tabatinga, município de Nísia Floresta, distantes 20 km (Gondim, 2004).
Os baixos índices de reavistagens dos animais suscitam questões acerca das limitações metodológicas durante a obtenção dos dados, bem como a respeito do ambiente utilizado pelos animais objeto desse estudo.
Muitos obstáculos devem ser superados para que se obtenham
dos com a utilização da técnica de foto-identificação a partir de embarcações turísticas. No presente trabalho, verificou-se a interferência de banhistas advindos da praia, bem como, os que estavam presentes nas embarcações turísticas e que, na parada para o banho, freqüentemente nadavam em direção aos animais, afugentando-os. Além disso, os turistas que permaneciam em cima da embarcação, na tentativa de obter registros fotográficos, não permitiam ao pesquisador posicionamento adequado, tant
e o percurso quanto durante a parada para banho. Observou-se ainda que barcos pesqueiros funcionaram como barreiras físicas em função da ancoragem desses no interior das enseadas, posicionando-se, dependendo do ângulo de observação, e
de turismo e o animal, dificultando a obtenção de fotografias. Ademais, as condições de ventos fortes e de mar com ondas elevadas, influenciaram na estabilidade do barco e, conseqüentemente, do pesquisador.
carreamento de nutrientes em direção ao mar (Odum, 1988). Por conseguinte, a permanência de 7 animais na região indica que esta pode fornecer suprimentos alimen
xemplo de movimentação semelhante acontece em Canan
tares, principalmente pela influência do complexo estuarino de Tibau do Sul, formado pela laguna de Guaraíras. Além disso, verifica-se uma maior proteção aos animais (especialmente aos grupos com filhotes), devido à topografia das enseadas que propicia locais abrigados próximos à costa (Link, 2000).
O baixo índice de reavistagem na área de estudo pode sugerir que haja grupos maiores próximos das enseadas e que ocorra uma grande movimentação entre os poucos indivíduos que apresentam fidelidade à área e àqueles que são avistados em algumas ocasiões utilizando a região, o que torna elevada a flutuabilidade do número de indivíduos na população. Um e
éia (E. L. A. Monteiro-Filho, comunicação pessoal, 30/10/06).
AGO SET OUT
15
0 3 6 9 12
L2 L3 L18 S20 S34 S49 S68
Animais Identificados
Nº de Av
ista
ge
ns
NOV DEZ JAN
AGO SET OUT NOV DEZ Período de Observação
JAN L2
L3
L18
S20
S34
S49
S68
igura 12 – Permanência temporal dos indivíduos foto-identificados considerados residentes durante o período de estudo.
4.2. PADRÕES DE ASSOCIAÇÃO
Diversos autores apontam dificuldades no estudo do S. guianensis, tais como a identificação dos animais e seus comportamentos (Yamamoto, 2002). Spinelli (2004) assinala a dificuldade em distinguir as faixas etárias infante e juvenil, e por isso os agrupou em uma única categoria: imaturos. Neste trabalho, apesar das observações terem sido realizadas a bordo de uma embarcação, conseqüentemente mais próximas dos animais, adotou-se a mesma categorização de Spinelli (2004), possibilitando melhor análise dos dados referentes aos padrões de associação.
De acordo com as observações realizadas durante o período de estudo
compostas pelas diferentes categorias etárias anteriormente definidas; (b) entre os animais foto-identificados; e, (c) predominantes para os animais foto-identificados L3 e S20, individualmente calculado entre esses e as diferentes faixas etárias anteriormente definidas, estas últimas formando díades ou não.
F
Os valores concernentes ao índice de associação (Jaccard) para as possíveis díades compostas pelas diferentes categorias etárias, foram alcançados levando-se em conta o número de indivíduos em cada uma delas e comparando-as. Analisando esses valores, averiguamos predominância entre as associações compostas por um adulto e um imaturo (1A + 1I) (HWI=0,477) e um adulto e dois imaturos (1A + 2I) (HWI=0,294). Esses dados representam as composições com maior estabilidade dentre todas as combinações possíveis, conforme demonstrado pela comparação das freqüências observadas para cada tipo de associação (Ȥ2=162,31; gl=10; p0,05)
(Tabela III), condizendo com o que foi encontrado por Sartorio (2005) para a espécie em questão na mesma área de estudo.
Tabela III – Índice de associação (Jaccard), freqüências observada e esperada para as possíveis díades durante o período de observação deste estudo. Os indivíduos adultos são representados por “A” e os
Associação Índice de Jaccard(HWI) FreqüênciaObservada FreqüênciaEsperada Percentual
imaturos por “I”.
Tipos de
1A + 1A 0,048 7 9,9 6,42%
1A + 1I 0,477* 43 9,9 39,45%
23 9,9 21,10%
1A + 3I 0,013 1 9,9 0,92%
1A 2A
1A + 2I 0,294*
+ 4I 0,014 1 9,9 0,92%
+ 1I 0,080 6 9,9 5,50%
2A + 2I 0,217 10 9,9 9,17%
2A + 3I 0,240 6 9,9 5,50%
2A + 4I 0,041 2 9,9 1,83%
3A + 3I 0,125 1 9,9 0,92%
1I + 1I 0,084 9 9,9 8,26%
Nota: (*) maiores índices de Jaccard encontrados.
observar diversidade nos tipos de associação, indicando um padrão dinâmico e,
os comportamentais (Gero, Bejder, Whitehead, Mann e Connor, 2005).
Com base na faixa etária e no número de indivíduos por agrupamento, pode-se
A partir do índice de similaridade de Jaccard, pode-se também verificar associações entre os indivíduos foto-identificados (Tabela IV). De todas as associações registradas, cinco alcançaram valores máximos (HWI=1), permanecendo associadas ao longo de todo o período de observação: S39/S40, L18/S51, S52/S53, S54/S55 e S49/S59. Contudo, esse é um valor que não expressa a dinâmica existente para esses animais, já que os mesmos somente foram vistos associados uma única vez durante o todo o período deste estudo.
Já quando analisamos os valores obtidos para as outras associações que ocorreram entre os animais reconhecidos naturalmente, observamos que L3/S20, S20/S34, bem como S43/S48 apresentaram os maiores índices (HWI=0,333), podendo sse valor indicar associações esporádicas para realização de atividades específicas.
nseada do Curral: em princípio, divididos em duplas (L3/I e S20/I),
depois foi visto que os dois ltos ( se junt ram em e,
post te retorn indo ao en ntro dos dois imaturos qu iam
perm os enqua adultos pescavam; em seguida voltaram à form inici I e S20/I). oborando a i ia acima, podemos citar Monteiro-Filho (2000), que observou para indivíduos da mesma espécie, no complexo estuarino lagunar de Cananéia, litoral sul do Estado de São Paulo, a ocorrência de associações contexto-específico.
na mesma área de estudo, a existência de assoc entre os in os L3 e S20, stas representa cerca de um dos rrelacionou os dias que os animais foram vistos por eles desempenhadas. Dessa maneira, não é possível inferir, a p
e
Tal condição ocorreu, por exemplo, em uma das ocasiões em que L3/S20 estavam associados na e
adu L3 e S20) aram, saí forrageio
eriormen aram, co e hav
anecido junt nto os ação
al (L3/ Corr dé
Jesus (2004) também constatou,
iações divídu e ndo sexto
dias observados. Porém, o autor não co associados com as atividades
Tabela IV – Índice de associação (Jaccard) encontrado
Associações ID Índice de Jaccard (HWI) para os indivíduos foto-identificados (ID).
S20/S34 0,333 S20/S48 0,125 S20/S68 0,142 S39/S40 1 S43/S48 0,333 S48/S68 S4 0,250 9/S59 1 S52/S53 1 S54/S55 1 L18/S51 1 L3/S20 0,333
Os resultados referentes às análises dos tipos predominantes das associações dos indivíduos L3 e S20, foram obtidos comparando-os com cada uma das faixas etárias anteriormente definidas – adulto (A) e imaturo (I) – e, entre esses e a possível díade formada – adulto/imaturo (AI).
Avaliando os resultados acima citados, fundamentado no índice de Jaccard, verificamos uma maior associação do indivíduo L3 com animais imaturos (HWI=0,454), quando comparamos sua associação com a dupla adulto/imaturo (HWI=0,243). Assim como para L3, o índice de associação de S20 com imaturo (HWI=0,105) foi mais elevado quando comparado com a díade adulto/imaturo (HWI=
nimais imaturos do que com outros adultos. Além disso, Jesus (2004), baseado a freqüência das observações, verificou o indivíduo L3 se associou um maior número de vezes com imaturos do que S20.
0,035) (Tabela V). A análise das freqüências observadas das díades formadas pelos indivíduos L3 e S20 com imaturo (L3/I e S20/I: Ȥ2=6,00; gl=1; p0,05)
apresentaram diferenças significativas. Já comparando a associação desses indivíduos foto-identificados com a dupla adulto/imaturo (L3/AI e S20/AI: Ȥ2=3,81; gl=1;
p=0,0510), não houve significância.
Sartorio (2005) observou que L3 estava presente nas enseadas mais associada com a
Tabela V– Índice de e S20 e entre esses e
as diferentes faixas et por “A”, imaturos por “I” e a díade
adulto-imaturo por “AI”.
Associações
ID/Faixa etária Índice de J(HWI) FreqüênciaObservada peradaqüência Percentual
associação (Jaccard) calculado, individualmente, entre L3 árias. Indivíduos adultos são representados
accard Fre
Es
L3/A 0 0 -
-S20/A 0 0 -
-L3/AI 0,225 9 5 90%
S20/AI 0,035 1 5 10%
L3/I 0,454 20 12 80%
S20/I 0,10 4 12 20%
* 5 (*) maior índice verificado.
Os dados do presen alho não podem confirmar a organização social atrilinear sugerida por Sartorio (2005), o qual se baseou na formação de associações de (a)
êmea, em função de alguns aspectos como o investi
dices de encefalização e tempo de vida longo, é plausív
te trab m
uma única fêmea acompanhada de um imaturo (filhote, sendo este seu ou não) ou (b) uma fêmea acompanhada de um imaturo (filhote) e um outro animal imaturo (filhotes ou juvenis). Entretanto, a associação freqüente entre L3 e indivíduos imaturos pode sugerir que se trata de uma f
mento parental típico dos mamíferos, o qual envolve gestação interna e lactação, com possibilidade de deserção reduzida da fêmea (Clutton-Brock, 1991).
Na presente pesquisa, também foram observadas associações entre indivíduos adultos, embora menos freqüentes. De acordo com Vonhof, Whitehead & Fenton (2004), os aspectos do sistema social entre mamíferos são únicos e, em geral, apontam para grupos baseados no parentesco ou na cooperação. O sistema cooperativo tem sido observado em uma grande variedade de taxa animal e uma outra maneira para analisar as associações seria considerá-las formações cooperativas (Norris & Schilt, 1988). Considerando-se que, de uma forma geral, os cetáceos caracterizam-se pela existência de estrutura social complexa, altos ín
el que esses animais apresentem episódios de cooperação e competição, possivelmente observados nas associações entre os indivíduos desse estudo (Connor et
5. CONCLUSÕES
ise os, a esquis que:
plicação da t nica de foto-identificação continua favorável para estudos
S. guianen região de Pipa.
qualidade das imagens obtidas para estudos que fazem uso da técnica de foto-identificação.
uos Após anál dos dados obtid presente p a constatou
a a éc
com sisna
condições ambientais influenciam na
os espécimes de S. guianensis que utilizam a região de Pipa apresentam elevada plasticidade.
os tipos de associação entre os indivíduos que utilizam a região de Pipa mostrou-se dinâmico.
não é possível inferir a respeito das associações existentes entre os indivíd foto-identificados que utilizam a região de Pipa.
6. REFERÊNCIAS
Acuña, L. B. (2002). Estimativa do tamanho da população do boto-tucuxi-marinho, Sotalia fluviatilis (Cetacea, Delphinidae) na região do estuário de Ca
es idade de São Paulo. São Paulo. 73pp.
lcock, J. (2001). The Evolution of Social Behavior (pp 421-455). In J. Alcock.
An
Ananias o boto-cinza, Sotalia
fluviatilis, avaliada através de foto-identificação. Graduação. Monografia, Un
Bejder, L.; Fletcher, D. & Bräger, S. (1998). A method for testing association patterns of social animals. Animal Behaviour,56: 719-725.
Ce .Canadian Journal of Zoology, 77:13-18.
arr, T. & Bonde, R. K. (2000). Tucuxi (Sotalia fluviatilis) occurs in Nicaragua, 800 Km
45
hilvers, B. L. & Corkeron, P. J. (2002). Association patterns of bottlenose dolphins (T
Jo
Clutton-Brock, T. H. (1991). The evolution of parental care. Princeton, University Press.
Connor, R. C.; Mann, J.; Tyack, P. L. & Whitehead, H. (1998). Social evolution in toothed whales. Trends in Ecology and Evolution, 13: 228-232.
Cunha, H. A.; Da Silva, V. M. F.; Lailson-Brito Jr, J.; Santo, M. C. O.; Flores, P. A. C.; Martin, A. R.; Azevedo, A. F.; Fragoso, A. B. L.; Zanellato, R. C. & Solé-Cava, A. M. (2005). Riverine and marine ecotypes of Sotalia dolphins are different species. Marine Biology,148: 449-457.
da Silva, V. M. F. & Best, R. C. (1996).Sotalia fluviatilis. Mammalian Species,527: 1-7pp.
nanéia, São Paulo, por meio de catálogo de foto-identificação para a pécie. Mestrado. Dissertação, Univers
A
imal Behavior. Sinauer, Sunderland.
, S. M. A. (2004). Recorrência e fidelidade espacial d
iversidade Federal do Rio Grande do Norte. Natal. 32pp.
Bräger, S. (1999). Association patterns in three populations of Hector’s dolphin,
phalorhynchus hectori
C
north of its previously known range. Marine Mammal Science, 16: 447-2.
C
ursiops aduncus) off Point Lookout, Queensland, Australia. Canadian
Defran, R. H; Schultz, G. M. & Weller, D. W. (1990). A technique for the photographic identification and cataloging of dorsal fins of the bottlenose
di Be
.
rto Alegre.
e II: Tópicos em
Flore ification study of the marine
néia, SP e na Baía de Guanabara, RJ. Mestrado. Dissertação,
Gero
ottlenose dolphins, Tursiops spp. Canadian
Gondim, M. A. (2004). Ecologia comportamental do boto-cinza Sotalia fluviatilis
ederal do Rio
Hard
Workshop on Research and
dolphin (Tursiops truncatus). Report International Whaling Commission, 12: 53-56.
neditto, A. P. M; Ramos, R. M. A. & Lima, N. R. W. (2001). Quem são os golfinhos? (pp. 9-24). In A. P. M. Di Beneditto, R. M. A. Ramos & N. R. W Lima (orgs). Os Golfinhos: Origem, classificação, captura acidental, hábito alimentar.Cinco Continentes Editora: Po
Fernandez, F. A. S. (1995). Métodos para estimativas de parâmetros populacionais por captura, marcação e recaptura. Pp 01-26, em P. R. Peres-Neto, J. L. Valentin e F. A. S. Fernandez (eds). Oecologia Brasiliensis, Volum
Tratamento de Dados Biológicos.
s, P. A. C. (1999). Preliminary results of a photoident
tucuxi,Sotalia fluviatilis, in southern Brazil. Marine Mammal Science,15: 840-847.
Geise, L. (1989). Estrutura social, comportamental e populacional de Sotalia sp (Gray, 1886); (Cetacea, Delphinidae) na região estuarino-lagunar de Cana
Universidade de São Paulo. São Paulo. 199pp.
, S., Bejder, L., Whitehead, H., Mann, J., & Connor, R. C. (2005). Behaviourally specific preferred associations in b
Journal of Zoology,83: 1566-1573.
(Gervais, 1853) na enseada de Tabatinga, litoral sul do Rio Grande do Norte. Bacharelado. Monografia de Graduação. Universidade F
Grande do Norte, Natal. 63p.
t, F. A. S., Cremer, M. J., Simões-Lopes, P. C. Tonello Jr., A. J. (2006). Padrões de residência do golfinho Sotalia guianensis (Cetacea, Delphinidae) na Baía da Babitonga, litoral norte de Santa Catarina, Brasil.
Hetzel, B. & Lodi, L. (1993). Boto-cinza (pp. 156-162). In B. Hetzel & L. Lodi (orgs).
Baleias, Botos e Golfinhos: guia de identificação para o Brasil. Nova Fronteira: Rio de Janeiro.
Jesus, A. H. (2004). Perfil comportamental do boto-cinza, Sotalia fluviatilis
Link, -cinza, Sotalia fluviatilis
Lodi, referencias do boto-cinza, Sotalia fluviatilis
Martin, P. & Bateson, P. (1993). General issues (pp 7-24). In P. Martin & P. Bateson
Mille
coleta de pegadas. Wildlife
Mont
ern Brazil. Ciência e Cultura Journal of
Nasc
rte. Mestrado. Dissertação, Universidade Federal
Norri
9-179. Odum, E. P. (1988). Ecologia. Guanabara Koogan: Rio de Janeiro.
(Gervais, 1853), (DELPHINIDAE, CETACEA), no litoral sul do Rio Grande do Norte, Brasil. Mestrado. Dissertação, Universidade Federal do Rio Grande do Norte. Natal. 70pp.
L. O. (2000). Dinâmica populacional do boto
(GERVAIS, 1853) (MAMMALIA: CETACEA), no Litoral do Rio Grande do Norte. Mestrado. Dissertação, Universidade Federal do Rio Grande do Norte. Natal. 76p.
L. F. (2002). Uso do habitat e p
(Cetacea, Delphinidae), na Baía de Paraty, Rio de Janeiro. Mestrado. Dissertação, Universidade Federal Rural do Rio de Janeiro. Seropédica. 167pp.
(orgs). Measuring Behavior: an introductory guide. Cambridge University Press: Cambridge.
r, C. M. (2001). Medir pegadas de onça pintada: um método promissor para identificação de indivíduos. Protocolo pra
Conservation Society. Belize.
eiro-Filho, E. L. A. (2000). Group organization of the dolphin Sotalia fluviatilis
guianensis in an estuary of southeast
the Brazilian Association for the Advancement of Science,52: 97-101.
imento, L. F. (2002). Descrição comportamental do boto-cinza, (Sotalia Fluviatilis (Gervais, 1853), DELPHINIDAE, CETACEA) no litoral sul do Estado do Rio Grande do No
do Rio Grande do Norte. Natal. 73pp.
Pizzorno, J. L. A. (1999). Estimativa populacional do boto-cinza, Sotalia fluviatilis, na Baía de Guanabara, por meio de catálogo de foto-identificação.
Dissertação de Mestrado. Seropédica: Universidade Federal Rural do Rio de
Quin
nandian Journal of Zoology,79: 447-456.
Ross
ahia, Brasil. Workshop on Research and Conservation of the
Santo
ersidade Federal do Rio Grande do Norte. Natal.
Shan
ce, 22:
Slote
nde do Norte, Natal. Janeiro. 47p.
tana-Rizzo, E. & Wells, R. S. (2001). Resighting and association patterns of bottlenose dolphins (Tursiops truncatus) in the Cedar Keys, Florida: insights into social organization.Ca
Ricklefs, R. E. (2003). A Economia da Natureza. Guanabara Koogan: Rio de Janeiro. i-Santos, M., Wedekin, L., & Monteiro-Filho, E. L. A. (2006). Residência e
fidelidade de área por Sotalia guianensis na região do estuário do Rio Caravelas, estado da B
genusSotalia. Resumos. 48p.
s, M. C. O. (1999). Novas informações sobre cetáceos no litoral sul de São Paulo e norte do Paraná com base em estudos sobre encalhes e na aplicação da técnica de foto-identificação individual de Sotalia fluviatilis (Cetacea, Delphinidae). Mestrado. Dissertação, Universidade de São Paulo. São Paulo. 114pp.
Sartorio, R. (2005). Padrões de agrupamento, comportamento e uso da área de Sotalia guianensis (CETACEA, DELPHINIDAE) no litoral sul do RN, Brasil. Doutorado. Tese, Univ
80pp.
e, S. H. (2004). Residence patterns, group characteristics, and association patterns of bottlenose dolphins near Sanibel Island, Florida. Gulf of Mexico Scien
1-12.
en, E.; Dawson, S. M. & Whitehead, H. (1993). Associations among pgotographically identified Hector’s dolphins. Canadian Journal of Zoology,
71: 2311-2318.
Spinelli, L.H.P. (2004). Brincadeira e participação de animais imaturos no comportamento de forrageio em botos cinza, Sotalia fluviatilis. Mestrado.
Tomas, W. M. & Miranda, G. H. B. (2004). Uso de armadilhas fotográficas em levantamentos populacionais (pp 243-268). In L. Cullen Jr., R. Rudran & C. Valladares-Padua (orgs). Métodos de Estudos em Biologia da Conservação e Manejo da Vida Silvestre. Fundação O Boticário de Proteção a Natureza /
Vonh
novel social
Whit
techniques (pp 65-87). In J. Mann, R. C. Connor,
Whit
es in the Study of Behavior,28: 33-74.
Würs (1990). Methods of photoidentification for small
Würs
sition and stability of coastal porpoises (Tursiops truncatus). Science,198:
Yam
xto de simpósio). In: Sociedade Brasileira de Etologia (Ed.),
Zolm
al Science, 18(4): 879-892.
Universidade Federal do Paraná, Curitiba.
of, M. J., Whitehead, H., & Fenton, M. B. (2004). Analysis of Spix’s disc-winged bat association patterns and roosting home ranges reveal a
structure among bats. Animal Behaviour,68: 507-521.
Whitehead, H. (1997). Analysing animal social structure. Animal Behaviour, 53: 1053-1067.
ehead, H.; Christal, J. & Tyack, P. L. (2000). Studying cetacean social structure in space and time: Innovative
P. L. Tyack & H. Whitehead (orgs). Cetacean Societies. University of Chicago Press, Chicago.
ehead, H. & Dufault, S. (1999). Techniques for Analyzing Vertebrate Social Structure Using Identified Individuals: Review and Recommendations.
Advanc
Würsig, B. (1978). Occurrence and group organization of Atlantic bottlenose porpoises (Tursiops truncatus) in an Argentine bay. Biological Bulletin, 154: 348-359.
ig, B. & Jefferson, T. A.
cetaceans.Reporting International Whaling Commission, 12: 43-52.
ig, B. & Würsig, M. (1977). The photographic determination of group size, compo
755-756.
amoto, M. E. (2002). Questões metodológicas na observação do comportamento de cetáceos (Te
Anais do XX Encontro anual de etologia (pp. 33-38). Natal.
Tabela A1 – Freqüência absoluta da avistagem dos espécimes foto-identificados (ID), reconhecidos durante o período de estudo. Em destaque, os indivíduos com freqüência de avistagem 5.
Período de Observação
2005 2006 ID
Agosto Setembro Outubro Novembro Dezembro Janeiro
Total de avistagens
L1 0 0 0 0 1 0 1
L2 1 1 0 1 1 0 4
L3 2 6 2 14 3 2 29
L11 0 0 0 1 0 0 1
L12 0 1 0 0 0 0 1
L13 1 1 0 0 0 0 2
L18 1 0 1 0 0 0 2
S20 0 3 2 2 4 0 11
S24 1 0 0 0 0 0 1
S25 0 0 0 0 1 0 1
Animais Identificados em estudos anterior
es
S33 0 2 0 0 0 0 2
S34 1 0 1 1 1 1 5
S35 1 0 0 0 0 0 1
S36 1 1 0 0 0 0 2
S37 1 0 0 0 0 0 1
S38 1 0 0 0 0 0 1
S39 1 0 0 0 0 0 1
S40 1 0 0 0 0 0 1
S41 1 0 0 0 0 0 1
S42 0 1 0 0 1
S43 0 1 1 0 0 0 2
S44 0 1 0 0 0 0 1
S45 0 1 0 0 0 0 1
S46 0 1 0 0 0 0 1
S47 0 1 0 0 0 0 1
S48 0 2 1 0 0 0 3
S49 0 1 0 1 1 0 3
S50 0 1 0 0 0 0 1
S51 0 0 1 0 0 0 1
S52 0 0 1 0 0 0 1
S53 0 0 1 0 0 0 1
S54 0 0 1 0 0 0 1
S55 0 0 1 0 0 0 1
S56 0 0 1 0 0 0 1
S57 0 0 0 1 0 0 1
S58 0 0 0 0 1 0 1
S59 0 0 0 0 1 0 1
S60 0 0 0 0 1 0 1
S61 0 0 0 0 1 0 1
S62 0 0 0 0 1 0 1
S63 0 0 0 0 1 0 1
S64 0 0 0 0 1 0 1
S65 0 0 0 0 1 0 1
S66 0 0 0 0 1 0 1
S67 0 0 0 0 1 0 1
S68 0 1 0 0 1 0 2
Animais Identificados no
prese
nte estudo
S69 0 0 0 1 0 0 1
TOTAL 14 26 14 22 23 3 102