Intracellular reprogramming of expression, glycosylation, and function of a plant-derived antiviral therapeutic monoclonal antibody.
Texto
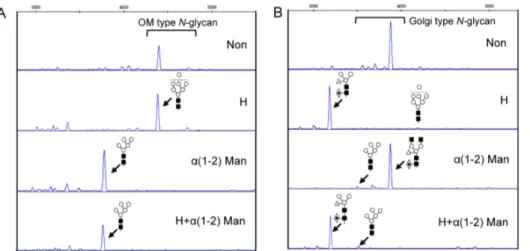
Imagem
![Figure 6. Stability profiles of mAb H , mAb P K and mAb P in mice. The % value was calculated with the formula [1006 (concentration of each day/concentration of the first day (24 hr) after injection)], followed up to day 10](https://thumb-eu.123doks.com/thumbv2/123dok_br/17235906.244764/6.918.89.574.93.369/figure-stability-profiles-calculated-concentration-concentration-injection-followed.webp)
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Ainda que as opiniões sejam dissonantes relativamente ao impacto que as conversas sobre sexualidade entre pais e filhos têm no comportamento sexual do
Este relatório relata as vivências experimentadas durante o estágio curricular, realizado na Farmácia S.Miguel, bem como todas as atividades/formações realizadas
Este modelo permite avaliar a empresa como um todo e não apenas o seu valor para o acionista, considerando todos os cash flows gerados pela empresa e procedendo ao seu desconto,
As doenças mais frequentes com localização na cabeça dos coelhos são a doença dentária adquirida, os abcessos dentários mandibulares ou maxilares, a otite interna e o empiema da
Buscando contribuir para uma solução dentro do contexto exposto acima, este trabalho analisa a substituição de parte da frota de veículos novos a gasolina por veículos
