OPEN ACCESS
International Journal of Zoological Research ISSN 1811-9778 DOI: 10.3923/ijzr.2016.1.10Research Article
Variation of Sediment Properties among the Radial Profiles of
Fiddler Crab Burrows in Mangrove Ecosystem
1
Mohammad Mokhtari,
1Mazlan Abd Ghaffar,
1Gires Usup and
1,2Zaidi Che Cob
1School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, National University of Malaysia, UKM Bangi 43600, Selangor, Malaysia
2
Marine Ecosystem Research Centre, Faculty of Science and Technology, National University of Malaysia, UKM Bangi 43600, Selangor, Malaysia
Abstract
Fiddler crabs burrow creates oxic-anoxic interfaces on the burrow walls. Accordingly burrow walls represent the transitions site between oxic and anoxic condition where the sediment properties varied significantly across it. In this study the burrows of three species of fiddler crabs including Uca rosea, Uca forcipata and Uca pardussumieri were sampled at three depth layers. Sediment properties of burrow walls including; temperature, redox potential, pH, density, porosity, water content, organic content, chlorophyll content and solid phase iron pools were measured to determine the magnitude of burrow effects on mangrove sediments. The results indicated that U. paradussumieri
effectively reduced the sediment porosity of surrounding sediments down to 45%. Oxidized layer was more extended around
U. paradussumieri burrows. Burrow walls of U. forcipata and U. paradussumieri contain higher water content than ambient sediment and burrows of U. rosea efficiently decreased the organic content of sediment. The PCA biplots indicated that the burrow walls of the all three species of fiddler crabs at 3 and 8 cm depth were correlated with oxidized iron. Ambient sediments of U. forcipata habitat were correlated with reduced iron and organic content. At 20 cm depth, burrow walls of U. paradussumieri were highly correlated with water content, while ambient sediment was correlated with reduced iron. The results of this study revealed that the thickness of oxidized layer varied according to sediment depth and burrow volume. Consequently the burrow effect varied significantly among different species of fiddler crabs as results of different habitat characteristics, sediment types and crab size.
Key words: Burrow walls, mangrove sediment, bioturbation, oxidation effect, principal component analysis
Received: October 01, 2015 Accepted: November 30, 2015 Published: February 15, 2016
Citation: Mohammad Mokhtari, Mazlan Abd Ghaffar, Gires Usup and Zaidi Che Cob, 2016. Variation of sediment properties among the radial profiles of fiddler crab burrows in mangrove ecosystem. Int. J. Zool. Res., 12: 1-10.
Corresponding Author: Zaidi Che Cob, School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, National University of Malaysia, UKM Bangi 43600, Selangor, Malaysia
Copyright: © 2016 Mohammad Mokhtari et al. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Competing Interest: The authors have declared that no competing interest exists.
INTRODUCTION
Fiddler crabs are the most abundant macrofuana on the mudflats of mangrove forest. They excavate and maintain a cylindrical j-shaped burrow with one entrance. They feed near their burrows and defends its territory against other fiddler crabs. Burrows used as refuge from predators and harsh environmental conditions (Crane, 1975; Macintosh, 1988). Fiddler crabs feed on bacteria, diatoms, ciliated protozoa and nematodes living within sediment grains by the use of their mouth appendages (Robertson and Newell, 1982; Hoffman et al., 1984; Meziane et al., 2002). These crabs are active during the low tide and before the onset of high tide they returned to their burrows and plugged the burrows entrance (Crane, 1975).
During the low tide oxygen penetrates into burrows and oxidizes the reduced subsurface sediments of burrow walls (Kristensen, 2008; Nielsen et al., 2003; Gribsholt et al., 2003; Furukawa, 2005). In spite of this burrow excavation and maintenance displace oxidized surface sediments with reduced subsurface layers (McCraith et al., 2003; Gutierrez et al., 2006). Fiddler crabs simply by increasing the oxic-anoxic interface area induce significant effects on biogeochemical properties of sediment (Kristensen, 2008).
Previous studies revealed that burrowing activity of fiddler crabs, increase turnover rate of sediments (Katz, 1980),
sediment metabolism and alter the partitioning of carbon oxidation pathways (Kostka et al., 2002; Nielsen et al., 2003; Gribsholt et al., 2003). Botto and Iribrane (2000) claimed that fiddler crabs alter sediment grain size, porosity and water content. Nielsen et al. (2003) indicated that the effect of fiddler crab burrow on carbon oxidation is limited to thin layers of burrow walls. They also stated that burrow walls of fiddler crabs are unique sites within anoxic sediment layer where iron reduction and re-oxidation of sulfide occur. Michaels and Zieman (2013) claimed that the oxidation effect of burrows is depended on sediment properties including organic content, porosity, drainage and sediment texture. Furukawa (2005) stated that biogeochemical properties of burrow wall is influenced by infaunal species and sampling site. Therefore, variations of sediment properties among the burrow walls must be examined at base of site and species. Consequently, in this study, burrow samples collected from three species of fiddler crab at different shore level will aid in better determination of burrow effects.
MATERIALS AND METHODS
Field sampling: The sampling was conducted in
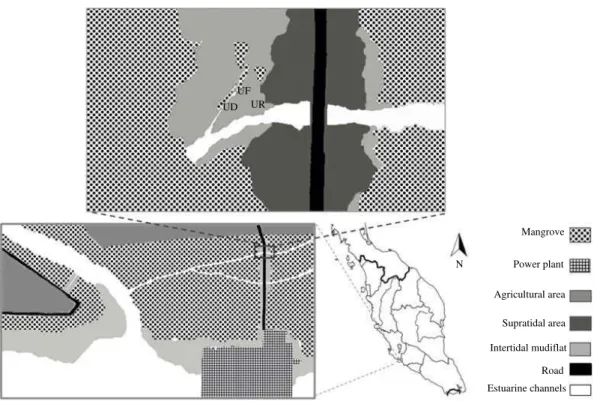
Sepang-Lukut mangrove forest on the west coast of peninsular Malaysia (Fig. 1). Six species of fiddler crab including;
U. paradussumieri, U. forcipata, U. rosea, U. perplexa,
Fig. 1: Location of sampling sites in the study area, UD: U. paradussumieri habitat, UF: U. forcipata habitat, UR: U. rosea habitat
N UD UR
UF
Mangrove
Power plant
Agricultural area
Supratidal area
Int. J. Zool. Res., 12 (1-2): 1-10, 2016
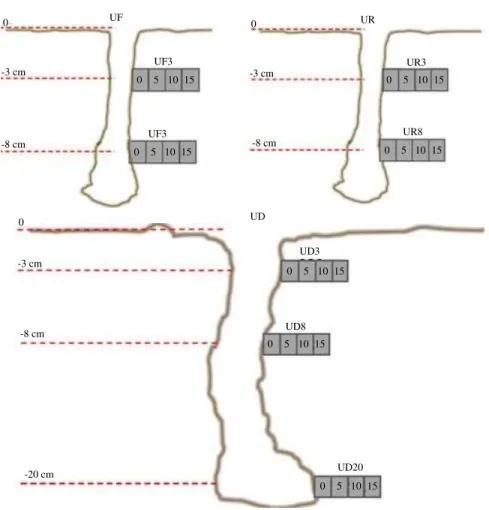
Fig. 2: Schematic figure of core sampling from the burrow walls of U. forcipata (UF), U. rosea (UR) and U. paradussumieri (UD). Each sediment core was expressed by the name of the crab species and the depth layer (e.g., UF3 sediment core represent the radial profile at 3 cm depth from U. forcipata burrow). The cells inside the core signify core slices and the numbers represent the distance of each slice from the burrow wall with scale of millimeter
U. annulipes and U. triangularis were found in this area. Among these, burrows of U. paradussumieri, U. forcipata and
U. rosea were chosen for this study, mainly because the core sampling from their burrow walls were more feasible due to their larger burrow size. In this area U. paradussumieri build the largest burrow with maximum length of 30 cm and 30 mm diameter. The size of the U. rosea and U. forcipata burrows are comparatively similar which usually extend down to 15 cm depth. Prior to sampling three burrows were selected for each species as replicate. Radial profiles of burrows were removed horizontally from the burrow walls by cut-end 5 mL syringe. In large burrows of U. paradussumieri sediment cores were collected from three depths of 3, 8 and 20 cm, while two sediment cores were collected at 3 and 8 cm depth from the burrows of U. rosea and U. forcipata (Fig. 2). Syringe cores were sealed by rubber stopper, labeled, packed in zipped plastic bags and stored in ice box immediately after collection. pH, temperature and redox potentials were measured in situ
from the burrow walls (at 3, 8 and 20 cm depth) with 0.5 cm intervals. The pH and temperature were measure by pH meter (Exstick Model pH 100) while redox potential was measured by ORP meter (Extech RE 300).
Laboratory analysis: In laboratory sediment cores were sliced
with intervals of 5mm. Sediment density was determined by weighting a known volume of sediment. Water content was measured by weight loss after drying the sediment samples at 70EC for 12 h. Accordingly Sediment porosity calculated by the following formula:
Sediment porocity = Density×water content/100
Organic content was estimated by the loss on ignition method at 520EC for 12 h. Chlorophyll content was measured by Lorenzen method (Lorenzen, 1967). Briefly, 0.5 g of sediment were incubated overnight in 90% acetone at 4EC
3
UF3
UF3 UR3
UR8
UD8 UD3
UD20 UD
-3 cm
-8 cm 0
0 5 10 15 0 5 10 15
0 5 10 15
0 5 10 15 0 5 10 15 0 5 10 15
0 5 10 15
-3 cm
-8 cm 0
-3 cm
-8 cm 0
-20 cm
followed by centrifugation at 3000 rpm for 5 min. The chlorophyll content was subsequently determined by spectrophotometer at 665 and 750 nm before and after acidification with 1 M HCl.
Solid phase iron pools of sediment (Fe II and Fe III) were determined by Kristensen and Alongi (2006) modified method of Lovley and Phillips (1988). About 300 mg of sediment was extracted in 5 mL of 0.5 M HCl for 30 min in the shaking platform. After centrifugation (3000 rpm for 5 min), 40 µL of aliquots was transferred to curvets containing ferrozine to determine Fe (II). One milliliter of supernatant was transferred to test the tube containing reducing agent hydroxylamine hydrochloride and was shaked for 15 min before mixing with ferrozine to determine total reducible iron pools. Therefore, Fe (III) amount was simply calculated by subtraction of total amount of reducible Fe with Fe (II).
Statistical analysis: Principal Component Analysis (PCA) with
aim of biplots were performed to compare sediment properties among the burrow walls, depth layer and species. In biplots the longest variables line represents the most influential variable. The distance between sample point and environmental variable line indicate the correlation between them. The lesser the distance observed between the point and line the more sample and variable are correlated. The data were mean centered and standardized prior running the PCA. All statistical analyses were performed by Minitab 16 (Minitab Inc., State College, PA, USA). Biplots were created by BiPlot add-on for Excel (Lipkovich and Smith, 2002).
RESULTS
The burrow density between the three fiddler crabs was in similar range. U. rosea had highest density with 28.3 individuals in one square meter while the density of U. forcipata and U. paradussumieri were 23 and 20 individual per square meter, respectively. The burrow of U. paradussumieri was significantly larger than two other species with mean of 17 mm burrow diameter comparing to 12 and 10 mm for U. forcipata and U. rosea.
Uca paradussumieri burrows were more abundant in low shore, U. forcipata burrows were commonly observed near mangrove trees while U. rosea was more frequent on mudflats with high sediment density. Sediment porosity, water and organic content was not different between the depth layers of 3, 8 and 20 cm. Sediment porosity at
U. paradussumieri habitat was higher than U. rosea and
U. forcipata habitat (Fig. 3g and 4g). Near the burrow wall of
U. paradussumieri, sediment porosity decreased to level of U. rosea and U. forcipata habitat. Sediment porosity did not change among the radial profile of U. rosea and
U. forcipata burrows (p>0.05). Near the burrow walls of
U. paradussumieri sediment porosity were 42-45% lower than ambient sediment.
Except in U. rosea habitat, higher water content was observed in the burrow walls (Fig. 3e and 4e). The highest water content was observed in burrow walls of
U. paradussumieri at 20 cm depth which contains 13% higher water content than ambient sediment (Fig. 4e). Organic content was higher in U. rosea habitat (Fig. 3f and 4f). Radial profiles from U. rosea burrows revealed that organic content was higher in ambient sediment especially at 8 cm. In
U. paradussumieri habitat, organic content was lower in burrow walls than ambient sediment in 3 cm depth, conversely at 20 cm organic content decreased towards the ambient sediment. Variation of organic content in U. forcipata
burrow profiles was less conspicuous although lower organic content at burrow walls was observed at 8 cm depth.
Redox potential was elevated at the burrow walls of UF3, UF8 and UD20 (Fig. 3b and 4b). In U. forcipata burrow walls, redox potential was 57 and 90% higher than ambient sediment at 3 and 8 cm depth, respectively. In 20 cm depth of
U. paradussumieri burrows redox potential was 30% higher than ambient sediment. pH was higher in U. forcipata
burrows comparing to U. rosea and U. paradussumieri habitat (Fig. 3a and 4a). Radial profile of burrows indicated that sediment pH is not affected by crab burrows. In U. forcipata
and U. rosea burrows temperature increased away from the burrow (Fig. 3c and 4c). In contrast at U. paradussumieri
burrows, temperatures were slightly higher than surrounding sediments. Only at 3 cm depth, chlorophyll content was significantly higher at the burrow walls (p<0.05) (Fig. 3d and 4d). In U. rosea burrow chlorophyll content was higher by 92% while in the burrows of U. paradussumieri and
U. forcipata, chlorophyll content were, respectively 78 and 63% higher than ambient sediment.
In all three crab burrows the amount of oxidized iron was higher in 3 cm depth profiles (Fig. 3h and 4h) particularly in
U. rosea burrow which located in high shore sediments. At 8cm depth of U. forcipata and U. rosea burrows oxidized iron pools were predominantly higher within 5 mm layer of burrow walls. In U. paradussumieri burrows oxidized iron were observed as far as 15 mm from the burrows. However in 20 cm depth the presence of oxidized iron was limited to the narrow 2 mm layer of burrow walls.
Int. J. Zool. Res., 12 (1-2): 1-10, 2016 7.5 7.0 6.5 6.0 5.5 5.0 pH
5 10 15 20 25
(a)
5 10 15 20 25
(b) 300 250 200 150 100 50 0 -50 -100 R edo x p o te n ti al ( m V ) UF3 UR3 UD3 36 34 32 30 28 26 T em p er at u re ( °C )
5 10 15 20 25
(c)
1.25 3.75 6.25 17.5
(d) 7 6 5 4 3 2 1 0 C h lo ro p h y ll (µ m o l g G 1) 25
0 10 20
(e) 60 55 50 45 W ater con ten t (% )
0 10 20
(f) 30 24 22 20 18 16 14 12 10 Or g an ic co n te n t ( % )
0 10 20 30
(g) 1.5 1.0 0.5 0.0 Po ro si ty mm
0 10 20
(h) 30 60 50 40 30 20 10 0 F e (l ll ) ( µ m o l g ) G 1 mm
Fig. 3(a-h): (a) Variation of pH, (b) Redox potential, (c) Temperature, (d) Chlorophyll, (e) Water, (f) Organic content, (g) Porosity and (h) Oxidized iron within the burrow walls of U. rosea (UR), U. forcipata (UF) and U. paradussumieri (UD) at 3 cm depth
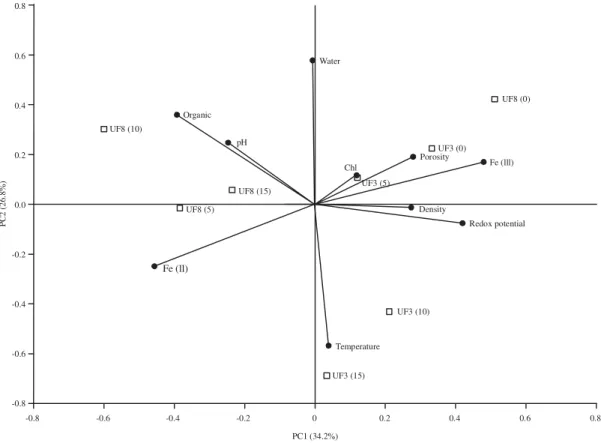
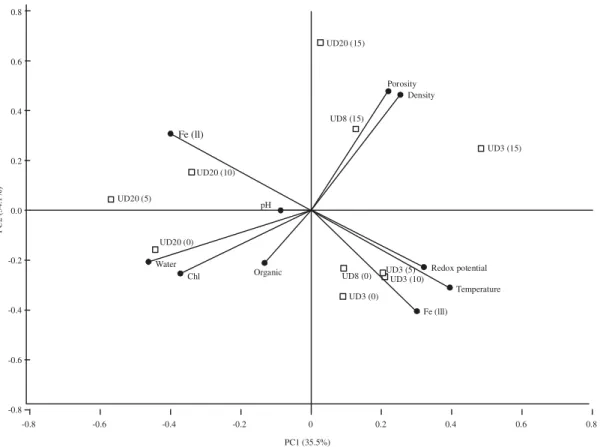
ambient sediments of 8 cm depth were more correlated with reduced iron (Fe II) and organic content (Fig. 5). On the other side UF3 (5) sample was correlated with chlorophyll content. Ambient sediment samples of 3 cm depth (UF3 (15) had lowest amount of water content and therefore showed a negative correlation with water content in PCA graph. Samples from U. rosea burrow wall (at 3 and 8 cm depth) were positively correlated with oxidized iron and chlorophyll (Fig. 6). Ambient sediment of 3 and 8 cm depth was more correlated with PH. Only UR8 (15) sample was particularly correlated with reduced iron. Burrow walls of U. paradussumieri at 3 and 8 cm depth were highly correlated with oxidized iron (Fe III) (Fig. 7). At 20 cm depth, burrow walls were correlated with water
content while ambient sediments were correlated with Fe (II). In all depth, ambient sediments of 15 mm radial profiles were correlated with porosity.
DISCUSSION
Powers and Cole (1976) found that burrows of U. panacea
are much cooler than surface sediments which could be used as heat refuge during the mid days. Gribsholt et al. (2003) recorded higher temperatures in ambient sediment than burrow wall which is related to higher water content measured at the burrow walls. In this study at the burrow walls of U. rosea and U. forcipata, lower temperature was measured.
5 10 15 20 25 (b) 300 250 200 150 100 50 0 -50 -100 7.5 7.0 6.5 6.0 5.5 5.0 pH
5 10 15 20 25
(a) UF8 UR8 UD8 UD20 36 34 32 30 28 26 T emp er atu re ( °C )
5 10 15 20 25
(c)
(f)
0 5 10 15 20 25 30
24 22 20 18 16 14 12 10 30
0 10 20
(e) W ater co nten t (% ) 63 61 59 57 55 53 51 49 47 45 15 25 5
0 10 20
(g) 30 1.9 1.4 0.9 0.4 P or os it y mm
5 15 25 0 10 20
(h) 30 F e (l ll ) (µ m o l g ) G 1 mm
5 15 25
60 50 40 30 20 10 0
1.25 3.75 6.25 17.5
(d) 7 6 5 4 3 2 1 0 R ed o x po te n tial ( m V ) C h lo ro p h y ll ( µ mo l g G 1 ) O rg an ic con ten t (% )
Fig. 4(a-h): (a) Variation of pH, (b) Redox potential, (c) Temperature, (d) Chlorophyll, (e) Water, (f) Organic content, (g) Porosity and (h) Oxidized iron within the burrow walls of U. rosea (UR), U. forcipata (UF) and U. paradussumieri (UD) at 8 and 20 cm depth
However, sediment temperature did not change significantly among the burrow profiles and exhibited no relationship with either depth or water content. It seems that the temperature did not vary significantly at the scale of a few cm in the mudflats.
At 3 cm depth pH was higher at the burrow walls of
U. paradusumieri and U. forcipata which is consistent with the results obtained from fiddler crabs burrow walls at georgia salt marsh (Gribsholt et al., 2003).
Both pH and redox potentials could affect the partitioning of carbon oxidation in sediment. Combination of low pH and
low redox potential enhance the iron reduction. In high pH values, redox potential must be lower to initiate iron reduction (Gotoh and Patrick, 1974). Connell and Patrick (1968) substantiated that sulfate reduction occur at redox potentials of less than -150 mV and pH between 6.5-8.5. In this study sediment of U. rosea habitat was more oxidized with lower pH value which is ascribed to the position of U. rosea habitat on the high shore. Uca rosea burrows commonly found on the high shore where the high sediment density facilitates the excavation and maintenance of burrow for U. rosea
Int. J. Zool. Res., 12 (1-2): 1-10, 2016
pH
Water
Chl Organic
0.8
0.6
0.4
0.2
0.0
-0.2
-0.4
-0.6
-0.8
Fe (lll)
Temperature
Redox potential UF8 (0)
Porosity
Density
Fe (ll)
-0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8
PC1 (34.2%)
P
C
2
(
2
6.
8%
)
UF3 (0)
UF3 (5) UF8 (15)
UF8 (10)
UF8 (5)
UF3 (10)
UF3 (15)
pH Water
Chl Organic
0.8
0.6
0.4
0.2
0.0
-0.2
-0.4
-0.6
-0.8
UR8 (5) Fe (ll)
Temperature
Redox potential UR8 (0)
Fe (lll)
Porosity
Density
-0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8
PC1 (47.5%)
P
C
2 (
16
.2
%
)
UR3 (15)
UR3 (10)
UD8 (10)
UR3 (5) UR3 (0)
UR8 (15)
Fig. 5: PCA biplot graph of the Burrow wall samples from U. forcipata burrow
Fig. 6: PCA bilpot of U. rosea Burrow wall samples
pH
Water
Chl Organic 0.8
0.6
0.4
0.2
0.0
-0.2
-0.4
-0.6
-0.8
UD20 (5)
UD20 (10)
UD20 (0)
UD3 (0) UD8 (0) UD3 (5)UD3 (10)
Fe (lll)
Temperature Redox potential UD8 (15)
UD3 (15) UD20 (15)
Porosity Density
Fe (ll)
-0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8
PC1 (35.5%)
P
C
2 (
3
4.1
%
)
Fig. 7: PCA biplot graph of the Burrow wall samples from U. paradussumieri burrow
exposure time during the low tide and therefore are more oxidized. Alternatively lower pH values due to higher accumulation rate of mangrove dribs is convincing in high shore sediments.
Fiddler crab burrows enhance below ground decomposition rate and marsh production through increasing soil drainage and redox potential (Bertness, 1985). Weissberger et al. (2009) claimed that the volume of oxidized sediment with burrowing animals is 100 m3 higher than
defuanated sediments in 1 h. In this area the effect of burrow on the redox potential was more apparent in 8 and 20 cm depth. It seems that in 3 cm depth, sediments were more oxidized through oxygen diffusion from the surface and therefore the effect burrow was not detectable in this layer. Similar oxidation pattern was reported from previous studies at 3 and 5 cm depth (Nielsen et al., 2003; Gribsholt et al., 2003).
In intertidal mudflats, the presence of chlorophyll " is an indication of benthic microalgae biomass. Benthic microalgae are the important producers in intertidal and subtidal habitats and constitute around 50% of primary production in estuarine ecosystem (De Jonge, 1994). Although, chlorophyll content was higher in the burrow, the burrows had no direct effect. Higher chlorophyll content is due to higher availability of light
in the burrow walls compared to the ambient sediment at the same depth. In this study, chlorophyll content of sediment decreases with increasing distance from surface sediments and burrow walls due to its dependency to sunlight. In contrast burrow walls in 20 cm depth had higher chlorophyll content than 8 cm depth. This is related to the fact that in the 20 cm depth burrows were filled with seawater and hence contain higher benthic microalgae than semi-dry sediments of burrow walls at 8 cm depth. The presence of microalgae indirectly influences the oxidation status and biogeochemical properties of sediments.
It was documented that the crab burrows increase sediment-water interface of sediment (Katz, 1980). Hence, in this study water content was higher near the burrow walls except in U. rosea burrows which commonly located at the high shore with minimum inundation time during a single tide. Gribsholt et al. (2003) found 2-9% higher water content in burrow walls. Botto and Iribrane (2000) also stated that burrowing activity increase water content. Nevertheless, the higher water content observed in ambient sediment of
Int. J. Zool. Res., 12 (1-2): 1-10, 2016
content. Oscillation in redox potential at subsurface bioturbated sediments enhance organic matter decomposition (Aller, 1994; Papaspyrou et al., 2010). On the other hand burrows could act as trap for deposition of organic debris (Botto and Iribrane, 2000) and consequently increase organic matter content of the sediments. Even though previous studies reported no significant change of organic content among the burrow profiles (Gribsholt et al., 2003), the sharp decrease of organic content in the burrow walls of
U. rosea might be result of longer exposure to atmospheric oxygen during the low tide. As in Fig. 3b and 4b redox potential was more elevated in U. rosea habitat. At 20 cm depth of U. paradussumieri burrows, organic content was higher in the burrow wall. This part of the burrow was commonly filled with seawater during the low tide which makes a favorable location for deposition of organic dribs and miofuanna organisms. Higher water content in the burrow walls of U. paradussumieri compared to ambient sediment might be the reason for the observed lower porosity. The results indicated that burrowing activity of U. paradussumieri
clearly reduced the sediment density of surrounding sediments. In contrast sediment density at the burrow walls of
U. forcipata and U. rosea were higher than ambient sediment. Some researchers claimed that some infaunal organisms pushed the surrounding sediment during burrow excavation which results in higher sediment density in the burrow walls (Aller and Yingst, 1980; Jones and Jago, 1993).
Uca rosea burrows which were located at high shore sediment, experienced longer exposure time during a low tide and thus are more oxidized comparing to other crab burrows. In the other hand burrow profiles of U. paradussumieri due to their larger size had higher oxidation effect (Fenchel, 1996).
Although, the oxidation effect of burrows decreased with depth, limited oxidation at 20 cm depth might also related to the higher water content. Iron pools were found in oxidized form at sediments surrounding the burrows. The thickness of oxidized layer varied according to sediment depth and burrow volume. In all three burrows the thickness of oxidized layer decreased with increasing depth. In spite of this, in large
U. paradussumieri burrows, oxidized layer was more extended. Alternatively, anoxic condition prevailed in ambient sediment of 8 and 20 cm depth where the iron pools were found in reduced form.
The results of PCA indicated that Sediment samples from the burrow walls in U. forcipata (in both 3 and 8 cm) were more oxidized and hence positively correlated with oxidized iron (Fe (III)) and redox potential. These layers are more exposed to sunlight and hence positioned close to Chlorophyll arrow in PCA graph. In contrast ambient sediments in 8 cm depth which were located in anoxic layer revealed a positive
correlation with reduced iron (Fe (II)) and organic content. It seems that ambient sediment of 3 cm depth was more influenced by evaporation effect of sunlight's. As the ambient sediment contain lower water content comparing to burrow walls and therefore higher temperature were recorded from these sediments. due to high oxidation status of high shore sediments, only a thin layer of U. rosea burrow walls were correlated with redox potential and oxidized iron. Large
U. praradussumieri burrows clearly had the higher oxidation capacity than other fiddler crab burrows. So the sediment samples from 10mm distance from the burrow walls were correlated with oxidized iron and only ambient sediments at 20 cm depth were correlated with reduced iron. There was a positive correlation between burrow wall of 20 cm depth and water content as the lower parts of U. paradussumieri
burrows were commonly filled with seawater.
CONCLUSION
In this study, crab burrows significantly affect sediment properties like redox potentials, solid phase iron pools, water content and density. In spite of this some sediment factors such as temperature, chlorophyll and organic content varied significantly among the burrow profiles. U. paradussumieri
burrows due to their larger volume had a higher oxidation effects. Extended oxidized layer around U. rosea burrows were related to location of burrows on the high shore mudflats. Our results revealed that the magnitude of burrow effect was controlled by burrow volume, location of crab burrow and shore level.
ACKNOWLEDGMENTS
The authors would like to thank the JIMAH Energy Ventures Sdn., Bhd, for their kind collaboration during the sampling occasions. This research was financially supported by National University of Malaysia (grant no: UKMGUP-2011-200).
REFERENCES
Aller, R.C. and J.Y. Yingst, 1980. Relationships between microbial distributions and the anaerobic decomposition of organic matter in surface sediments of Long Island Sound, USA. Mar. Biol., 56: 29-42.
Aller, R.C., 1994. Bioturbation and remineralization of sedimentary organic matter: Effects of redox oscillation. Chem. Geol., 114: 331-345.
Bertness, M.D., 1985. Fiddler crab regulation of Spartina alterniflora production on a new England salt marsh. Ecology, 66: 1042-1055.
Botto, F. and O. Iribarne, 2000. Contrasting effects of two burrowing crabs (Chasmagnathus granulata and
Uca uruguayensis) on sediment composition and transport in estuarine environments. Estuarine Coastal Shelf Sci., 51: 141-151.
Connell, W.E. and W.H. Patrick Jr., 1968. Sulfate reduction in soil: Effects of redox potential and pH. Science, 159: 86-87. Crane, J., 1975. Fiddler Crab of the World. Princeton University
Press, Princeton, New Jersey.
De Jonge, V.N., 1994. Wind-Driven Tidal and Annual Gross Transport of Mud and Microphytobenthos in the Ems Estuary and its Importance for the Ecosystem. In: Changes in Fluxes in Estuaries: Implications from Science to Management, Dyer, K.R. and R.J. Orth (Eds.). Olsen and Olsen, Fredensborg, Denmark, ISBN: 9788785215222, pp: 29-40.
Fenchel, T., 1996. Worm burrows and oxic microniches in marine sediments. 1. Spatial and temporal scales. Mar. Biol., 127: 289-295.
Furukawa, Y., 2005. Biogeochemical Consequences of Infaunal Activities. In: Interactions between Macro and Microorganisms in Marine Sediments, Kristensen, E., R.R. Haese and J. Kostka (Eds.). American Geophysical Union, Washington, DC., pp: 159-177.
Gotoh, S. and W.H. Patrick, 1974. Transformation of iron in a waterlogged soil as influenced by redox potential and pH. Soil Sci. Soc. Am. J., 38: 66-71.
Gribsholt, B., J.E. Kostka and E. Kristensen, 2003. Impact of fiddler crabs and plant roots on sediment biogeochemistry in a Georgia saltmarsh. Mar. Ecol. Progr. Ser., 259: 237-251. Gutierrez, J.L., C.G. Jones, P.M. Groffman, S.E.G. Findlay,
O.O. Iribarne, P.D. Ribeiro and C.M. Bruschetti, 2006. The contribution of crab burrow excavation to carbon availability in surficial salt-marsh sediments. Ecosystems, 9: 647-658. Hoffman, J.A., J. Katz and M.D. Bertness, 1984. Fiddler crab
deposit-feeding and meiofaunal abundance in salt marsh habitats. J. Exp. Mar. Biol. Ecol., 82: 161-174.
Jones, S.E. and C.F. Jago, 1993. In situ assessment of modification of sediment properties by burrowing invertebrates. Mar. Biol., 115: 133-142.
Katz, L.C., 1980. Effects of burrowing by the fiddler crab,
Uca pugnax (Smith). Estuarine Coastal Mar. Sci., 11: 233-237. Kostka, J.E., B. Gribsholt, E. Petrie, D. Dalton, H. Skelton and E. Kristensen, 2002. The rates and pathways of carbon oxidation in bioturbated saltmarsh sediments. Limnol. Oceanogr., 47: 230-240.
Kristensen, E. and D.M. Alongi, 2006. Control by fiddler crabs (Uca vocans) and plant roots (Avicennia marina) on carbon, iron and sulfur biogeochemistry in mangrove sediment. Limnol. Oceanogr., 51: 1557-1571.
Kristensen, E., 2008. Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. J. Sea Res., 59: 30-43. Lipkovich, I. and E.P. Smith, 2002. Biplot and singular value
decomposition macros for Excel. J. Stat. Software, 7: 1-15. Lorenzen, C.J., 1967. Determination of chlorophyll and
pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr., 12: 343-346.
Lovley, D.R. and E.J.P. Phillips, 1988. Manganese inhibition of microbial iron reduction in anaerobic sediments. Geomicrobiol. J., 6: 145-155.
Macintosh, D.J., 1988. The Ecology and Physiology of Decapods of Mangrove Swamps. In: Aspects of Decapod Crustacean Biology, Fincham, A. and P.S. Rainbow (Eds.). Oxford University Press, London, ISBN-13: 978-0198540083, pp: 315-341.
McCraith, B.J., L.R. Gardner, D.S. Wethey and W.S. Moore, 2003. The effect of fiddler crab burrowing on sediment mixing and radionuclide profiles along a topographic gradient in a Southeastern salt marsh. J. Mar. Res., 61: 359-390.
Meziane, T., M.C. Sanabe and M. Tsuchiya, 2002. Role of fiddler crabs of a subtropical intertidal flat on the fate of sedimentary fatty acids. J. Exp. Mar. Biol. Ecol., 270: 191-201.
Michaels, R.E. and J.C. Zieman, 2013. Fiddler crab (Uca spp.) burrows have little effect on surrounding sediment oxygen concentrations. J. Exp. Mar. Biol. Ecol., 448: 104-113. Mokhtari, M., M.A. Ghaffar, G. Usup and Z.C. Cob, 2015.
Determination of key environmental factors responsible for distribution patterns of fiddler crabs in a tropical mangrove ecosystem. PLoS ONE, Vol. 10. 10.1371/journal.pone.0117467 Nielsen, O.I., E. Kristensen and D.J. Macintosh, 2003. Impact of
fiddler crabs (Uca spp.) on rates and pathways of benthic mineralization in deposited mangrove shrimp pond waste. J. Exp. Mar. Biol. Ecol., 289: 59-81.
Papaspyrou, S., M. Thessalou-Legaki and E. Kristensen, 2010. The influence of infaunal (Nereis diversicolor) abundance on degradation of organic matter in sandy sediments. J. Exp. Mar. Biol. Ecol., 393: 148-157.
Powers, L.W. and J.F. Cole, 1976. Temperature variation in fiddler crab microhabitats. J. Exp. Mar. Biol. Ecol., 21: 141-157. Robertson, J.R. and S.Y. Newell, 1982. Experimental studies of
particle ingestion by the sand fiddler crab Uca pugilator
(Bosc). J. Exp. Mar. Biol. Ecol., 59: 1-21.