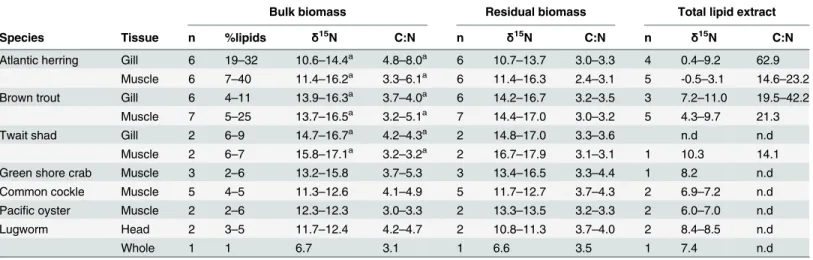
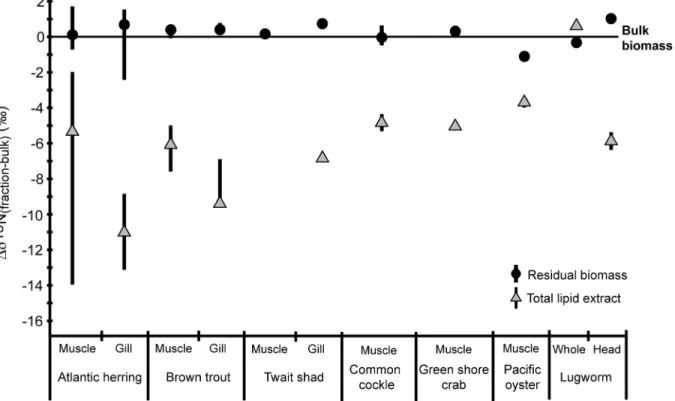
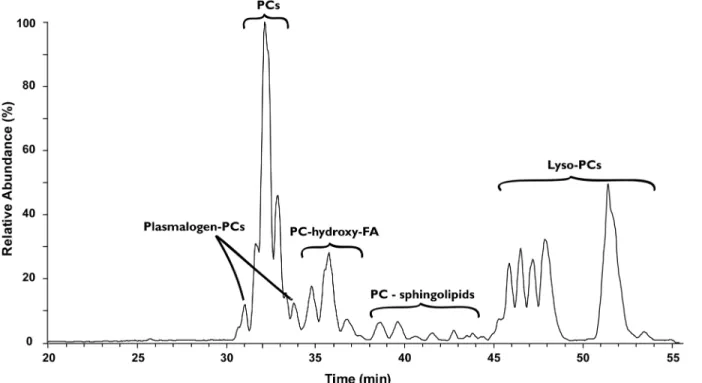
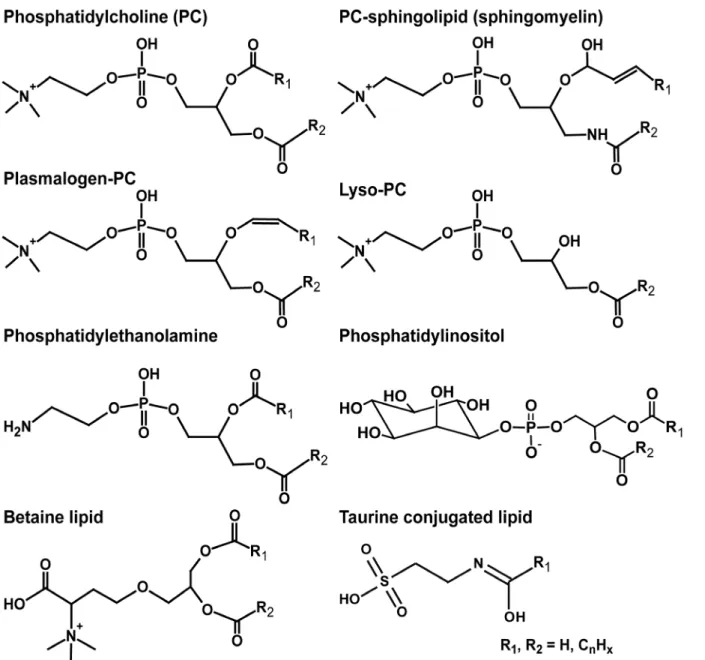
Factors Controlling the Stable Nitrogen Isotopic Composition (δ15N) of Lipids in Marine Animals.
Texto
Imagem
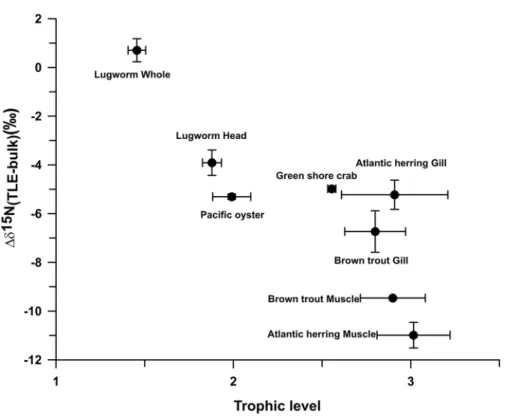
Documentos relacionados
Citation: Ma J-Y, Sun W, Liu X-N, Chen F-H (2012) Variation in the Stable Carbon and Nitrogen Isotope Composition of Plants and Soil along a Precipitation Gradient in Northern
Os valores do coeficiente de alfa de Cronbach obtidos por ano escolar e sexo são também satisfatórios, bem como os valores obtidos por áreas avaliadas (interesses por
Entre estas encontram-se a área da Quinta da Lousa, que compreende a urbanização com esse nome e dois outros aglomerados con guos; a área dos Lagueirões, que é talvez
Carbon and nitrogen stable isotope ratios of aquatic consumers in Polo Stream indicated that mangrove-derived carbon (likely in the form of detritus) is the most important carbon
The influence of nitrogen (N) limitation on the cell growth, chlorophyll content, intracellular lipid production and expression levels of two pathway genes in Chlorella
Assim, este estudo assume importância ao determinar o impacto da depressão na funcionalidade percebida e na performance das pessoas idosas no contexto de
A investigação realizada consistiu na implementação, em sala de aula, de uma pasta de tarefas de geometria plana, concebidas para a investigação, com o objectivo de gerar
custos da deficiência, e não patrimoniais, decorrentes da anulação do direito de procriar de forma livre e esclarecida e do sofrimento derivado da deficiência. 6) As
