Arch. BioI.Sci., Belgrade, 55 (1-2),5P-6p, 2003.
RELATIONSHIP BETWEEN ESTIMATES OF SPATIAL AND TEMPORAL, PLASTICITY, GENETIC
VARIABILITY AND BETWEEN-POPULATION DIFFERENTIATION IN
IRIS PUMILA.
A. Tarasjev,
Department of Evolutionary Biology, Institute for Biological Research, "S.Stankovic" University of Belgrade,
11060 Belgrade, Serbia and Montenegro
UDC 582.579.2: 581.54
The ability of a genotype to phenotypically respond to changes in environmental conditions(e.g.phenotypic plasticity sensuBra d s haw 1965) is of major interest in evolutionary biology because it is a possible mechanism of adaptation to heterogeneous environments (P i gIi u c c i 2001 and refe-rences therein). Many authors have expressed the expectation that phenotypic plasticity and within-population genetic vari-ability should be negatively correlated as "alternative ways of dealing with environmental heterogeneity" (references in SchiIi c h tin g and Lev i n 1984), but plasticity is also a factor that can, under certain circumstances, maintain genetic variability (Z h i v0 t0v sky and G a v r i let s 1992).Itis also often assumed that high phenotypic plasticity should 'act as a "buffer against selection" and therefore inhibit evolution-ary change (G ran t 1985), but We s t E be r h a r d t (1989) advocated that phenotypic plasticity could facilitate evolutionary change in various ways. Therefore, relationship of
phenotypic plasticity with genetic differentiation between and genetic variability within populationsshould'beextensively stu-died. In those studies, use of samples from natural populations in the most common natural environments for estimation of quantitative genetic parameters is highly recommended (Clark 1987).
In this work we examined the relationship between phe-notypic plasticity (both spatial and temporal) and genetic vari-ability (both within and between populations) in natural popu-lations of dwarf bearded iris -- Iris pumila L. (Iridaceae). Analysis was performed on morphometric data set from reci-procal-transplant experiment on two I. pumila populations (spatial aspect of phenotypic plasticity) which covered three flowering seasons (temporal aspect of phenotypic plasticity). Location and habitat characteristics of the analyzed popula-tions, experimental design, and 20 utilized floral and vegetative traits were given in detail in Tar a s j e v (1995,1997).
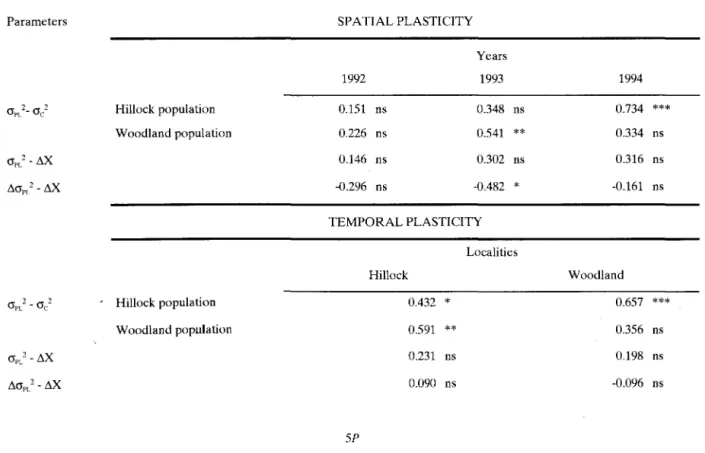
Table 1. Relationsbip between phenotypic plasticity (spatial and temporal) and genetic variability (within and between populations) measured by Spearman corelation coefficients among following parameters: plastic variances«JPL2),between-clone variances«Jc2),
trait divergences between populations(f,.X),and divergence of plastic variances between populations(f,.(JPL2).For estimation proce-dures see text.
Parameters
Hillock population
Woodland population
SPATIAL PLASTICITY
Years
1992 1993 1994
0.151 ns 0.348 ns 0.734
***
0.226 ns 0.541
**
0.334 ns0.146 ns 0.302 ns 0.316 ns
-0.296 ns -0.482
*
-0.161 nsTEMPORAL PLASTICITY
Localities
Hillock population
Woodland population
5P Hillock
0.432
*
0.591
**
0.231 ns
0.090 ns
Woodland
0.657
***
0.356 ns
0.198 ns
6P
All analyses were performed on replicate means trans-formed to natural logarhitms. Prior to that, the weights were transformed by cubic root in order to correct for dimensiona-lity (H0 uIe 1992). In order to examine relationships among plasticity, genetic variability and between population diver-gence of different traits, we calculated Spearman rank correla-tion coefficients (S0k aI and R0hi f 1981) among them. Rank correlations were also calculated among amounts of be-tween population divergence for plasticity and divergence of traits themselves. The estimation of correlation coefficients between those parameters over different traits is theoretically justified because they are specific for the trait in question (B r ads haw 1965), but significance levels can be inflated due to intertrait correlations and, therefore, should be ap-proached with caution. Compared parameters were estimated in the following manner. For every year/population combina-tion, two-way ANOVA was performed with Site, Clone and Site x clone as effects. Spatial plasticity was estimated from variance components determined by Restricted Maximum Likelihood (REML) analysis as the sum of Site and Site x clone components, a measure that is identical to "plastic variance"
(O"PL2) of S c h e i n e rand Goo d n i g h t (1984). Estimation of temporal plasticity was done in a similar fashion for every site/population combination by addition of Year to Year x clone component. Variance between clones(O"c2)was taken as a measure of within-population genetic variability. Since the effect of trait means on variances was removed by logarithmic transformation, genetic variances of different traits were in fact proportional to trait "evolvabilities" sensu H0uIe (1992). For comparison of plasticity with between-population trait divergence, average trait plasticity over both populations was used. Amounts of between-population divergence for traits themselves, as well as for phenotypic plasticity were estimated by the absolute value of Canberra metrics (S n eat hand So k aI 1973) on trait mean values and plastic variances re-spectively. For the analysis of spatial plasticity, trait diver-gences were averaged over sites, while for the analysis of tem-poral plasticity, trait divergences were averaged over years. Statistical analysis was performed by SAS statistical package (S A SIn s tit ute 1989) with PROC MEANS, PROC GLM and PROC CORR procedures.
Correlations between amounts of plasticity divergence and amounts of trait divergence were negative in sign (tempo-ral plasticity on Hillock being the exception), but statistically insignificant for both types of plasticity with the exception of spatial plasticity in 1993. Therefore, the amount of trait diver-gence betweenIris pumila populations was not positively cor-related with the amount of between-population divergence in trait plasticity, which indicates a possibility for independent evolution. Within-population genetic variability was positively correlated with both spatial (all correlations positive, two of them significant) and temporal plasticity (all correlations posi-tive, two out of three significant). Hence, two possible ways of dealing with environmental heterogeneity (genetic variability and phenotypic plasticity of a trait) were not mutually exclu-sive. Finally, correlations between plasticity and the amount of trait divergence between populations (between-population genetic variability) were positive in sign but statistically insig-nificant for both types of plasticity. Accordingly, contrary to predictions that plasticity acts as a "buffer against selection", more plastic traits in 1.pumilsare shown not to diverge less.
Acknowledgements- This work was supported by MSTD of Serbia. Research grant NO.I725.
References:Bradshaw, A. D. (1965).Adv. Genet.13, 115155. -Clark, A. G. (1987).Am .Nat. 129, 932-40. - Grant, V. (1985). The Evolutionary process: A Critical Rewiew of Evolutionary Theory. Columbia Univ. Press. - Houle, D. (1992). Genetics
130, 195-204. - Pigliucci, M. (2001) Phenotypic plasticity. J.
Hopkins Univ, - SAS Institute (1989).SAS/STAT User's Guide, Version6,4thed. - Scheiner, S. M. and J.C.Goodnight. (1984).