w ww.e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Cissampelos
sympodialis
has
anti-viral
effect
inhibiting
dengue
non-structural viral protein-1 and pro-inflammatory mediators
Fagner Carvalho Leite
a,b, Cíntia da Silva Mello
c, Luciana Gomes Fialho
c, Cintia Ferreira Marinho
c,
Ana Luisa de Araujo Lima
a, José Maria Barbosa Filho
d, Claire Fernandes Kubelka
c,
Marcia Regina Piuvezam
a,∗aLaboratório de Imunofarmacologia, Departmento de Fisiologia e Patologia, Universidade Federal da Paraíba, João Pessoa, PB, Brazil
bFaculdade São Francisco da Paraíba, Cajazeiras, PB, Brazil
cLaboratório de Imunologia Viral, Instituto Oswaldo Cruz, Fundac¸ão Oswaldo Cruz, Rio de Janeiro, RJ, Brazil
dLaboratório de Fitoquímica, Departmento de Ciências Farmacêuticas, Universidade Federal da Paraíba, João Pessoa, PB, Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received2November2015 Accepted6March2016 Availableonline19May2016
Keywords:
Cissampelossympodialis
Dengue Huh-7cells Cytokines NS1 Warifteine
a
b
s
t
r
a
c
t
Dengueisthemostimportantviralinfectiontransmittedamonghumansbyarthropod-borne.Thereare currentlynovaccinesorspecifictherapeuticaltreatment.Therefore,immunomodulatorycompounds fromplantshavebeenwidelyexaminedfortheirantiviraleffects.CissampelossympodialisEichler, Menis-permaceae,hasscientificallyproventopresentimmunomodulatoryactivities.Hereweassessedthe antiviralactivityofleafhydroalcoholicextract,warifteineormethylwarifteinefromC.sympodialisin aninvitrodenguevirusinfectionmodel.Theresultsdemonstratedthatleafhydroalcoholicextractor warifteine/methylwarifteinetreatmentdidnotreducedenguevirus-Ag+hepatocyte(Huh-7cell)rates inpresentexperimentalconditions.However,weassessedthepotentialantiviraleffectofleaf hydroalco-holicextractorwarifteine/methylwarifteineondenguevirus-infectionbytheproductionofinflammatory molecules,TNF-␣,MIF,IL-8andPGE2.DenguevirusinfectionenhancedTNF-␣,MIF,IL-8andPGE2 pro-ductionininfectedHuh-7cellsandleafhydroalcoholicextractbutnotwarifteine/methylwarifteine treatments,significantlyreducedthesemoleculesininfectedcells.Indenguevirus-infectedHuh-7cells, non-structuralprotein-1isproducedandleafhydroalcoholicextractsignificantlyinhibitedit indepen-dentlyofalkaloids.Ourfindingsimplythatleafhydroalcoholicextractmayattenuatedenguevirus infectioninHuh-7cellsbyinhibitingtheenhancedofpro-inflammatorymediatorsandnon-structural protein-1productioninducebydenguevirusindependentlyofwarifteine/methywarifteineitsmajor compound.
©2016SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Thisisanopen accessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
Dengue virus (DENV) is an important human infectious
pathogeninthetropics andsubtropics;itremainsanimportant
publichealth burden requiring continuing attention. The
clini-calmanifestations of dengue diseaserange fromasymptomatic
infection,undifferentiatedfeveranddenguefever,todengue
hem-orrhagicfeverwithplasmaleakageandpotentiallylife-threatening
dengueshocksyndrome(Amorimetal.,2014).
∗ Correspondingauthor.
E-mail:[email protected](M.R.Piuvezam).
Ethnobotanicalstudieshavebeentheprimarysourcefor
selec-tionofmoleculesinscientificinvestigationsandtheyrepresenta
richtrialforimmunomodulation products.Cissampelos
sympodi-alisEichler,Menispermaceae,popularlyknownasmilonaoccurs
inseveralBrazilianstatessuchasParaíba.Theaqueousinfusionof
theleaveshasbeenusedinfolkmedicinetotreatinflammatory
dis-eases(Piuvezametal.,2012).Theleafhydroalcoholicextract(AFL)
and its bisbenzylisoquinoline alkaloidspresented
immunomod-ulatory effectin severalexperimental modelsof inflammations
(Piuvezametal.,2012).
Inthepresentstudy,wehypothesizedthatAFL,warifteineor
methylwarifteinedisplayantiviraleffectviaits
immunomodula-toryproperties.ToproveitweusedtheinvitromodelofDENV
infectioninHuh-7cells(humanhepatocytecelllineage).
http://dx.doi.org/10.1016/j.bjp.2016.03.013
Materialsandmethods
Plantmaterial,obtainingandpreparationofleafhydroalcoholic extract(AFL)ofCissampelossympodialis
TheleavesofCissampelossympodialisEichler,Menispermaceae,
wereobtainedfromtheBotanicalGardenoftheFederalUniversity
ofParaiba(vouchespecimenAGRA1456)andtheextractofthe
leaves(AFL)aswellasitalkaloidswarifteine(WAR)and
methyl-warifteine(MWAR)weregentlyprovidedbyDrJoseMariaBarbosa
Filho(DeFreitasetal.,1996;Vieiraetal.,2013).Inbrief,3kgoffresh
leaveswascollected,driedandpulverized.After,threesuccessive
alcoholextractionswereperformedinapercolatoratroom
tem-perature(25–30◦C).TheAFLwasobtainedwithamixtureofwater
andethanol(30/70,v/v).Thensolventwasremovedandthedry
weightoftheextractwas79.9%basedonthepresentsolidwaste.
TheAFLwaspreparedinsterilesaline(Piuvezametal.,1999)to
posteriorstandardizationusingthealkaloidwarifteineasapattern
(Cerqueira-Limaetal.,2010).AFLwasdissolvedindimethyl
sulf-oxide(DMSO)andstocksolutions(1mg/ml)storedat−20◦C.AFL
wasdilutedtotheindicatedconcentrations(0.1–100g/ml)with
culturemediumbeforeuseinexperiments.DMSOconcentration
didnotexceed0.01%.
Bisbenzylisoquinolinealkaloidsextractionandpurification
Toobtainwarifteineandmethylwarifteine,theC.sympodialis
extractwasdissolvedin3%HClandextractedwithCHCl3.
Aque-ousfractionwasbasifiedwithNH4OHatpH9andagainextracted
withCHCl3.TheCHCl3 extractwaswashedwithH2Oanddried
withMgSO4togetthetertiaryalkaloidfraction.After,thetertiary
alkaloidfractionwassubjectedtochromatography columnover
alumina,elutingwithhexanecontainingCHCl3/MeOH.Fractions
elutedwithCHCl3–MeOH(49:1)werefurtherpurifiedbythinlayer
column(1mmlayer)forisolationofWARandMWAR(
Cerqueira-Limaetal.,2010;Meloetal.,2003;Vieiraetal.,2013).WARor
MWARpowder(purity90%)wasdissolvedin0.1NHCl.Foreach
experiment,thestocksolutionwasfurtherdilutedwith0.1NHCl
todesiredconcentrations(1,2.5,5,10M).
CulturemediumandpreparationofHuh-7cells
Huh-7cells(hepatocarcinomacellline),obtainedfrom
Ameri-canTypeCultureCollection(ATCC,cellline-615),weremaintained
inDulbecco’sModifiedEagleMedium(DMEM)(Sigma, St.Louis,
MO)supplementedwith10%fetalbovineserum(FBS)(Biological
Industries,KibbutzBeitHaemek,Israel),100U/mlpenicillin,and
100g/mlstreptomycin(InvitrogenLifeTechonologies).
MTTassayforcellviabilitycytotoxicityofAFL,warifteineand methylwarifteine
Huh-7cells wereincubatedin96-wellplates at1×105 cells
perwellcontaining100lofDMEMmediumanddifferent
con-centrationsofAFL(0at200g/ml),warifteine(WAR,0–200M)
or methylwarifteine (MWAR, 0–200M) for 72h. Cells were
washedoncebeforeadding50lFBS-freemediumcontainingMTT
(5mg/ml).After4hofincubationat37◦C,themediumwas
dis-cardedandtheformazanbluethatformedinthecellswasdissolved
inDMSO(SIGMA).Theopticaldensitywasmeasuredat540nm
(Mosmann,1983).
Cellcultures,virusstockpreparationandvirustitration
TheDENVserotype2strain16681hasprovidedbyDr.SB
Hal-stead(Naval Medical Research Center, USA) and propagated in
Aedes albopictusC6/36cellclonetoobtainedthevirusstock,as
describedbefore(Reisetal.,2008).Inbrief,A.albopictusC6/36cell
clonewasgrownasmonolayersat28◦ConLeibovitzmedium
(L-15)supplementedwith200mMglutamine,1%non-essentialamino
acid solution, 0.5% tryptose phosphate broth, 100 U/penicillin,
10g/streptomycinand5%fetalbovineserum(FBS)andinfected
withDENV-2for8days.After,thesupernatantcontainingthevirus
particleswasultracentrifuged(100,000×g)for1hat4◦C.The
pel-letwasstoredat−70◦CandVirustiterwascalculatedas50percent
tissuecultureinfectiousdose(TCID50).Thevirusstockusedwas
ataconcentrationof1.6×109TCID50/ml(Reisetal.,2008;
Lima-Junioretal.,2013).
Huh-7cellsinfectionandtreatmentwithAFL,warifteineor methylwarifteine
Huh-7 cells were resuspended in supplemented RPMI 1640
medium,plus10%FCSandseededat2×106cells/mlon96-or
24-wellplates.Afteranovernightincubation,infectionwaseffected
witha diluted inoculum (30 or300l) in cell culturemedium
containing1.6×109TCID50/ml.Aftera2h-incubationperiodfor
adsorption,thecellculturesupernatantwasreplacedwitha2%FBS
mediumandincubatedwithleafhydroalcoholicextract(AFL)ofC.
sympodialis(0.1,1or10g/ml),WAR(0.1,1or10M)orMWAR
(0.1,1 or 10M)and subsequently incubatedat 37◦C with5%
CO2.After24,48or72h,supernatantswerecollectedandstocked
at−20◦Cforcytokinemeasurementandcellsrecoveredforviral
antigendetermination,cellviabilitydeterminedinculturebyMTT
assay.Wellcontentwithcellcontrol,inactivatedandinfectious
DENVwasassayed.
ViralantigendeterminationinHuh-7cellsbyflowcytometry
Huh-7cellswererecoveredinacoldcellculturemedium,set
at1×106cells/microtube,thencentrifuged(350
×g,10min)and
washedoncewithphosphatebufferedsalinepH7.4containing1%
bovineserumalbuminand0.1%NaN3(PBS/BSA).Afterwards,cells
werefixedwithparaformaldehyde2%inPBS/BSAat4◦Cfor20min
andpermeabilizedwithsaponin0.15%inPBS/BSA.Permeabilized
cellswerethenblockedwith5%inactivatedplasmainPBS/BSAat
4◦Cfor30minandincubatedwithmouseanti-DengueComplex
monoclonalantibody(MAB8705,Millipore)at4◦Cfor60min.Cells
werethenwashedandincubatedfor30minat4◦Cwithanti-mouse
IgGAlexaFluor488(A20181,LifeTechnologies).Afterincubation,
cells werewashed withPBS/BSA,resuspendedin
paraformalde-hyde2%,andkeptat4◦C untilcellacquisition(5000 eventsfor
gated monocytes) by FACS® Calibur flow cytometer (Beckon &
Dickinson)andanalyzedwithFlowJoSoftware(TreeStarInc.).An
isotype-matchedantibodywasadoptedasastainingnegative
con-trol(Lima-Junioretal.,2013).
Cytokinequantification
ELISAcytokinekitswasusedtomeasureTNF-␣,IL-8,andMIF
in the cells supernatant, and the assaywas performed
accord-ing to themanufacturer’s instructions (R&D Systems, CA,USA)
as described previously (Assuncao-Miranda et al., 2010). Data
analyses of all assays were performed with Bio-Plex Manager
software(Bio-Rad). Prostaglandin(PG)E2 concentrationsin the
cellculturesupernatants fromHuh-7cells weredeterminedby
anenzymeimmunoassay(ELISA)kitaccordingtotheprocedures
suppliedbythemanufacturer(CaymanChemical,AnnArbor,MI,
50 60
a
b
c
1000**
*
800
600
400
200
0 40
20
0
Inf
ection,
%
TNF-α
pg/ml
MIF pg/ml
40
30
20
10
0
Control DENV-2 AFL 0.1 µg/ml AFL 1 µg/ml AFL 10 µg/ml
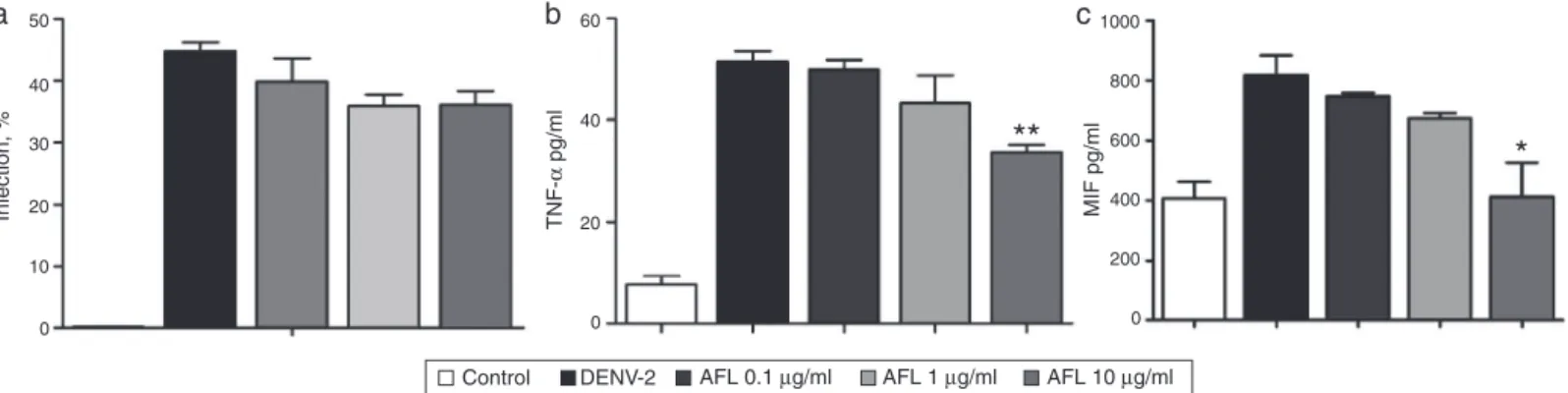
Fig.1. EffectofAFLonDenguevirustype2(DENV-2strain16681)Dinfection,TNF-␣andMIFproductionininfectedHuh-7cells.(A)DENVantigenpositivecellsdetected byflowcytometryanalysis.(B)TNF-␣and(C)MIF-␣levelsdetectedbyELISA.Resultswerepresentedasmean±SEMwhere*p<0.05wasconsideredsignificantbyone-way ANOVAandaDunnetttest,asapost-testwhenthetestsubstancewerecomparedwithDENV-2.Dataarerepresentativeofthreeindependentexperimentsperformedin triplicates.
NS1quantification
NS1wasmeasuredinculturemediumfromHuh-7cellsinfected
withdenguevirusandpre-treatedusingacommercialsandwich
ELISAkit(PlateliaNS1;BioRadLaboratories).Thetestswere
con-ductedwithaccordingtothemanufacturer’sprotocols.
Resultsanddiscussion
Immunomodulatory compounds of plants have been widely
examinedfortheirantiviraleffects(AbdKadiretal.,2013;Reis
etal.,2008).Inthisstudy,anti-DENVactivityofleafhydroalcoholic
extract(AFL)ofC.sympodialis,warifteine(WAR)anditsmethylated
formmethylwarifteine(MWAR)themajoralkaloidsisolatedfrom
theplantwereevaluated.TheconcentrationofAFLorWAR/MWAR
usedwastestedbyMTT assay(seesection Culturemediumand
preparationofHuh-7cells)andtheydonotshowtoxiceffectson
Huh-7cells(datanotshown).
To determine whether AFL has anti-DENV effect, we firstly
measuredDENV-antigenexpression,TNF-␣andMIFproductionin
Huh-7cellsinfectedbyDENV-2.Wepre-infectedHuh-7cellsfor
2handtreatedwithAFL(0.1,1and10g/ml)andcellswere
col-lectedat72hpost-infection.AsshownonFig.1A,theplantextract
(AFL)didnot interfere withtheexpressionof DENV. However,
AFL(10g/ml) reduced the production of TNF-␣ at 72h
post-infection(Fig.1B)andreducedMIFproductionbyHuh-7infected
cells(Fig.1C).
TNF-␣isthemaincytokinethatinducesvascularleakageand
itsserumlevelshavebeenimpliedinDENV-infectedpatientswith
seriousformsofdisease(Bragaetal.,2001;Sellahewa,2013).In
mice,vascularhemorrhagecanbeinducedbyinjectionofDENV
andislesssevereinTNF-␣deficientmiceorblockingby
anti-TNF-␣antibodies(Chenetal.,2007;Shrestaetal.,2006).Furthermore
(Perez et al., 2010)demonstrated an association between high
levelsofexpressionofTNF-␣alleleandsusceptibilityofdengue
hemorrhagicfever(DHF).
AnotherimportantcytokineinDENVinfectionismacrophage
migrationinhibitoryfactor(MIF).Assuncao-Mirandaetal.(2010)
showedenhancedofMIFonplasmaofpatientswithsevereforms
ofdengue.Therefore,endogenousMIFcontributestothe
pathogen-esisofdengueinfection.Inthepresentstudy,invitroinfectionof
Huh-7cellswithDENV-2increasedMIFlevelsthatwere
success-fullysuppressedbyAFLtreatment(Fig.1C)
ToconfirmtheimmunomodulatoryactivityofAFLwemeasured
onsupernatant,IL-8andPGE2levels.AFL(10g/ml)inhibitedboth
IL-8andPGE2(Fig.2AandB).Interleukin-8andPGE2induce
alter-ationsinendothelialfunctions(Purwatietal.,2011;Lima-Junior
etal.,2013).Elevatedlevelsofthesemoleculesweredetectedin
serumandpleuralfluidofpatientswithDHF(Raghupathyetal.,
1998;Purwatietal.,2011)suggestingthatthefluideffusionintothe
pleuraandotherserouscavitiesinDHFpatientsmaybeatleastin
partattributedtoIL-8and/orPGE2.TheproductionofIL-8appears
tobedependentonIL-1andTNF-␣(Raghupathyetal.,1998)andas
itwasrelatedaboveAFLdecreasedtheamountofTNF-␣on
DENV-2infectedHuh-7cellssuggestingacytokinedownregulatoryeffect
ofAFL.
In DENV infection, hepatocytes produce and activate
cyclooxygenase-2(COX-2)withproduction ofPGE2(Liou etal.,
2008).Theso-calledbreak-bonepainindenguediseaseis
prob-ably related to the overproduction of PGE2 and this molecule
800
**
*
2015
10
5
0
IL-8 pg/ml PGE2 pg/ml
600
400
200
0
Control DENV–2 AFL 10 µg/ml
a
b
Fig.2.EffectofAFLonDENV-2inducedIL-8,PGE2orNS1productionininfectedHuh-7cellstreatedwithAFL(10g/ml)for2hpre-infection.Thesupernatantofcellswere
collectedat48hpost-infectionwithDenguevirustype2(DENV-2strain16681).(A)IL-8levelinDENVinfectedHuh-7cellstreatedwithAFL(B)PGE2levelinDENVinfected
hasimportant effect on replicationand infectivity of the virus (SteerandCorbett,2003;Tsatsanisetal.,2006).ThereforeCOX-2
modulationbytheplant extractmaybe a promisingtarget for
controllingnotonlyinflammationandpainbutalsoviralinfection
indicatinganimportantroleoftheplantinthisdisease(Roccaand
FitzGerald,2002;SteerandCorbett,2003).
DENVis anenvelopedvirus,and itsgenomeiscomposedof
apositive-sense,single-strandedRNAcodingforthreestructural
proteinspresentinthevirionand infectedcellsandseven
non-structural(NS)proteins(NS1,NS2a,NS2b,NS3,NS4a,NS4b,and
NS5) not present in the virion (Amorim et al., 2014).NS1 is a
glycoprotein expressed in infected mammaliancells as soluble
monomers that is subsequently transported tothe cell surface
where theproteinremains and/orit is releasedintothe
extra-cellularmilieubeenessentialforviralreplicationandviability.In
addition,thisproteinappearsintothebloodstreamandseveral
lab-oratorialtestsuseNS1todiagnoseDENVinfections(Amorimetal.,
2014).Inthisstudy,weshowedthatAFLsignificantlyreducedNS1
productioninDENV-2infectedHuh-7cells(Fig.3).
TodeterminethemechanismofactionofAFLoninvitrodengue
infectionwetestedtwomajoralkaloidsfromtheplant:warifteine
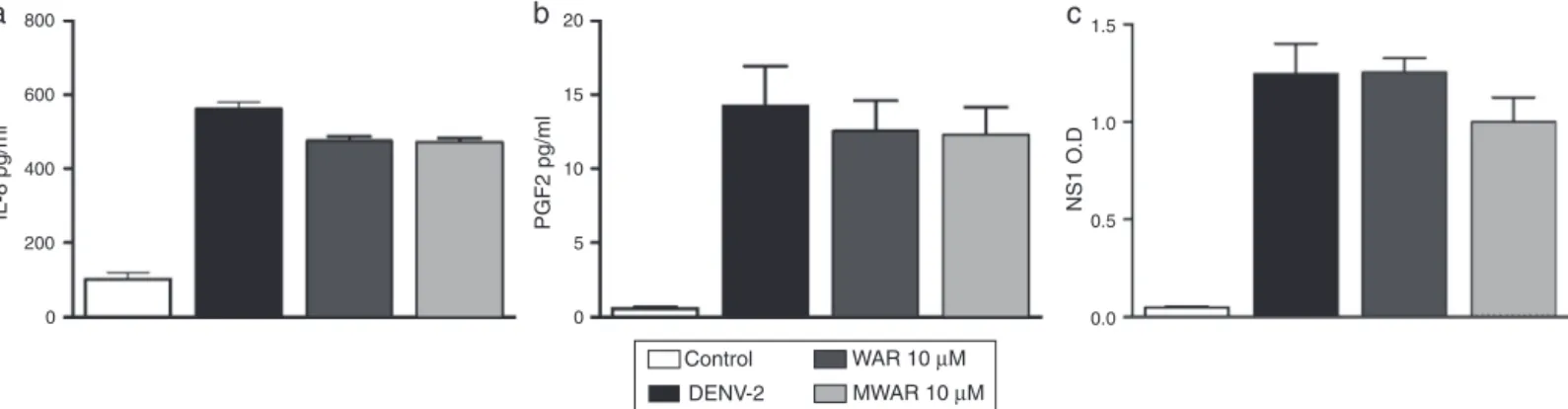
(WAR)anditsmethylateformmethylwarifteine(MWAR).Asitis
shownonFig.4A,WAR/MWARtreatmentdidnotreduceDENV-2
antigenininfectedHuh-7cellsanddidnotinhibitTNF-␣orMIF
production(Fig.4BandC).Inaddition,neitherWARnorMWAR
reducedIL-8,PGE2norNS1productionbyDENV-2infected
Huh-7cells(Fig.5A–C)indicatingthatAFLeffectisindependentlyof
2.0
*
NS1 O
.D
1.5
1.0
0.5
0.0
Control DENV–2 AFL 10 µg/ml
Fig.3.EffectofAFLonDENV-2inducedNS1productionininfectedHuh-7cells treatedwithAFL(0.1,1or10g/ml)for2hpre-infection.Thesupernatantofcells
werecollectedat24,48or72hpost-infectionwithdenguevirustype2 (DENV-2strain16681).NS1levelinDENV-infectedHuh-7cellstreatedwithAFLwere determinedbyELISAinthesupernatantsofHuh-7cells72hpostinfection,where *p<0.05,**p<0.01wasconsideredsignificantbyone-wayANOVAandaDunnett test,asapost-testwhenthetestsubstancewerecomparedwithDENV.Dataare representativeofthreeindependentexperimentsperformedintriplicates.
50
a
b
60c
Control DENV–2
WAR 0.1 µM
WAR 1 µM MWAR 0.1 µM
MWAR1 µM
MWAR10 µM
WAR 10µM
800
600
400
200
0 40
20
0 40
30
% inf
ection
MIF pg/ml
TNF-α
pg/ml
20
10
0
Fig.4.EffectofWARorMWARonDenguevirustype2(DENV-2strain16681)Dinfection,TNF-␣andMIFproductionininfectedHuh-7cells.(A)DENVantigenpositivecells
detectedbyflowcytometryanalysis.(B)TNF-␣and(C)MIF-␣levelsdetectedbyELISA.Resultswerepresentedasmean±SEMwhere*p<0.05wasconsideredsignificant byone-wayANOVAandaDunnetttest,asapost-testwhenthetestsubstancewerecomparedwithDENV-2.Dataarerepresentativeofthreeindependentexperiments performedintriplicates.
800
a
b
c
1.0 1.5
NS1 O
.D
PGF2 pg/ml
IL-8 pg/ml 0.5
0.0 15
20
10
5
0 600
400
200
0
Control DENV-2
WAR 10µM MWAR 10 µM
WAR/MWARandthattheplantextractpresentsothercompounds
responsiblefortheantiviraleffect.
Conclusion
Inconclusion,DENV2infectedHuh-7cells produceTNF,MIF,
IL-8andPGE2thatarecorrelatedwithNS1productionandAFL
suppressedtheviral-inducedinflammatoryresponsebydecreasing
the cytokine production as well as NS1 release, but failed to
affectvirusproduction.Conversely, warifteine/methylwarifteine
did not affect the inflammatory mediators nor virus
produc-tion in DENV-2 infected Huh-7 cells. These data shed light on
dengue virus infection and C. sympodialis extract capable to
reduceNS1and pro-inflammatorymediatorsdemonstratingthe
potentialanti-inflammatoryandantiviral therapyof thisherbal
medicine.
Ethicaldisclosures
Protectionofhumanandanimalsubjects. Theauthorsdeclare
thatnoexperimentswereperformedonhumansoranimalsfor
thisstudy.
Confidentialityofdata. Theauthorsdeclarethatnopatientdata
appearinthisarticle.
Righttoprivacyandinformedconsent. Theauthorsdeclarethat
nopatientdataappearinthisarticle.
Authors’contributions
FCL,CFMandLGF(PhDstudents)contributedtothe
develop-ment of thebiological protocols. CSMand ALAL(MS students)
contributedtotheELISAassay.JMBFcontributedinplant
identi-ficationandalkaloidisolation.CFKandMRPdesignedthestudy,
supervisedthelaboratoryworkandcontributedtocriticalreading
ofthemanuscript.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
This study was financially supported by CNPq-Universal14/
2012-472853/2012-0;CAPES/Brazil;PROEP, IOCand Plataforma
Tecnológica(RPT11D)/FIOCRUZCSM, LGF and CM are graduated
studentsatPosgbpIOC,Fiocruz.
References
AbdKadir,S.L.,Yaakob,H.,MohamedZulkifli,R.,2013.Potentialanti-dengue medic-inalplants:areview.J.Nat.Med.67,677–689.
Amorim,J.H.,Alves,R.P.,Boscardin,S.B.,Ferreira,L.C.,2014.Thedenguevirus non-structural1protein:risksandbenefits.VirusRes.181,53–60.
Assuncao-Miranda,I.,Amaral,F.A.,Bozza,F.A.,Fagundes,C.T.,Sousa,L.P.,Souza,D.G., Pacheco,P.,Barbosa-Lima,G.,Gomes,R.N.,Bozza,P.T.,DaPoian,A.T.,Teixeira, M.M.,Bozza,M.T.,2010.Contributionofmacrophagemigrationinhibitoryfactor tothepathogenesisofdenguevirusinfection.FASEBJ.24,218–228.
Braga,E.L.,Moura,P.,Pinto,L.M.,Ignacio,S.R.,Oliveira,M.J.,Cordeiro,M.T.,Kubelka, C.F.,2001.Detectionofcirculanttumornecrosisfactor-alpha,solubletumor necrosisfactorp75andinterferon-gammainBrazilianpatientswithdengue feveranddenguehemorrhagicfever.Mem.Inst.OswaldoCruz96,229–232.
Cerqueira-Lima,A.T.,Alcantara-Neves,N.M.,deCarvalho,L.C.,Costa,R.S., Barbosa-Filho,J.M., Piuvezam,M.,Momtchilo,R.,Barboza,R.,de JesusOliveira, E., Marinho,A.,Figueiredo,C.A.,2010.EffectsofCissampelossympodialisEichl.and itsalkaloidwarifteine,inanexperimentalmodelofrespiratoryallergytoBlomia
tropicalis.Curr.DrugTargets11,1458–1467.
Chen,H.-C.,Hofman,F.M.,Kung,J.T.,Lin,Y.-D.,Wu-Hsieh,B.A.,2007.Bothvirus andtumornecrosisfactoralphaarecriticalforendotheliumdamageinamouse modelofdenguevirus-inducedhemorrhage.J.Virol.81,5518–5526.
DeFreitas,M.R.,Cortes,S.F.,Thomas,G.,BarbosaFilho,J.M.,1996.Modificationof Ca2+metabolismintherabbitaortaasamechanismofspasmolyticactionof
warifteineabisbenzylisoquinolinealkaloidisolatedfromtheleavesof
Cissam-pelossympodialisEichl.(Menispermaceae).J.Pharm.Pharmacol.48,332–336.
Lima-Junior,R.S.,MelloCda,S.,Siani,A.C.,Valente,L.M.,Kubelka,C.F.,2013.Uncaria
tomentosaalkaloidalfractionreducesparacellularpermeability.IL-8andNS1
productiononhumanmicrovascularendothelialcellsinfectedwithdengue virus.Nat.Prod.Commun.8,1547–1550.
Liou,J.T.,Chen,Z.Y.,Ho,L.J.,Yang,S.P.,Chang,D.M.,Liang,C.C.,Lai,J.H.,2008. Differ-entialeffectsoftriptolideandtetrandrineonactivationofCOX-2.NF-kappaB, andAP-1andvirusproductionindenguevirus-infectedhumanlungcells.Eur. J.Pharmacol.589,288–298.
Melo,P.S.,deMedeirosCavalcante,H.M.,Barbosa-Filho,J.M.,deFatimaFormiga MeloDiniz,M.,deMedeiros,I.A.,Haun,M.,2003.Warifteineandmilonine, alka-loidsisolatedfromCissampelossympodialisEichl:cytotoxicityonrathepatocyte cultureandinV79cells.Toxicol.Lett.142,143–151.
Mosmann,T.,1983.Rapidcolorimetricassayforcellulargrowthandsurvival: appli-cationtoproliferationandcytotoxicityassays.J.Immunol.Methods65,55–63.
Perez,A.B.,Sierra,B.,Garcia,G.,Aguirre,E.,Babel,N.,Alvarez,M.,Sanchez,L.,Valdes, L.,Volk,H.D.,Guzman,M.G.,2010.Tumornecrosisfactor-alphatransforming growthfactor-beta1,andinterleukin-10genepolymorphisms:implicationin protectionorsusceptibilitytodenguehemorrhagicfever.Hum.Immunol.71, 1135–1140.
Piuvezam,M.R.,Bezerra-Santos,C.R.,Bozza,P.T.,Bandeira-Melo,C.,Vieira,G.C., Costa,H.F.,2012.Cissampelossympodialis(Menispermaceae):anovel phytother-apicweaponagainstallergicdiseases?In:Pereira,C.(Ed.),AllergicDiseases– HighlightsintheClinic,MechanismsandTreatment.InTech,Croatia,p.554.
Piuvezam,M.R., Pec¸anha,L.M.T., Alexander,J., Thomas, G.,1999. Cissampelos sympodialisEichlleafextractincreasestheproductionofIL-10by concanavalin-A-treatedBALB/cspleencells.J.Ethnopharmacol.67,93–101.
Purwati, Nasronudin, Kusumowidagdo, E.R., Rantam, F.A., 2011. The relation betweenlevelsofTNF-alpha,IL-1betaPGE2andPLA2withtheseveritydegree ofdenguehemorrhagic.Microbiol.Indones.5,5.
Raghupathy,R.,Chaturvedi,U.C.,Al-Sayer,H.,Elbishbishi,E.A.,Agarwal,R.,Nagar, R.,Kapoor,S.,Misra,A.,Mathur,A.,Nusrat,H.,Azizieh,F.,Khan,M.A.,Mustafa, A.S.,1998.ElevatedlevelsofIL-8indenguehemorrhagicfever.J.Med.Virol.56, 280–285.
Reis,S.R.,Valente,L.M.,Sampaio,A.L.,Siani,A.C.,Gandini,M.,Azeredo,E.L.,D’Avila, L.A.,Mazzei,J.L.,Henriques,M.,Kubelka,C.F.,2008.Immunomodulatingand antiviralactivitiesofUncariatomentosaonhumanmonocytesinfectedwith DengueVirus-2.Int.Immunopharmacol.8,468–476.
Rocca,B.,FitzGerald,G.A.,2002.Cyclooxygenasesandprostaglandins:shapingup theimmuneresponse.Int.Immunopharmacol.2,603–630.
Sellahewa,K.H.,2013.Pathogenesisofdenguehaemorrhagicfeveranditsimpact oncasemanagement.ISRNInfect.Dis.2013,6.
Shresta,S.,Sharar,K.L.,Prigozhin,D.M.,Beatty,P.R.,Harris,E.,2006.Murinemodel fordenguevirus-inducedlethaldiseasewithincreasedvascularpermeability.J. Virol.80,10208–10217.
Steer,S.A.,Corbett,J.A.,2003.TheroleandregulationofCOX-2duringviralinfection. ViralImmunol.16,447–460.
Tsatsanis, C., Androulidaki, A., Venihaki, M., Margioris, A.N., 2006. Signalling networksregulatingcyclooxygenase-2.Int.J.Biochem.CellBiol.38,1654–1661.