Stereotypy and variation of the mating call in the Lusitanian toadfish, Halobatrachus didactylus
10
0
0
Texto
Imagem

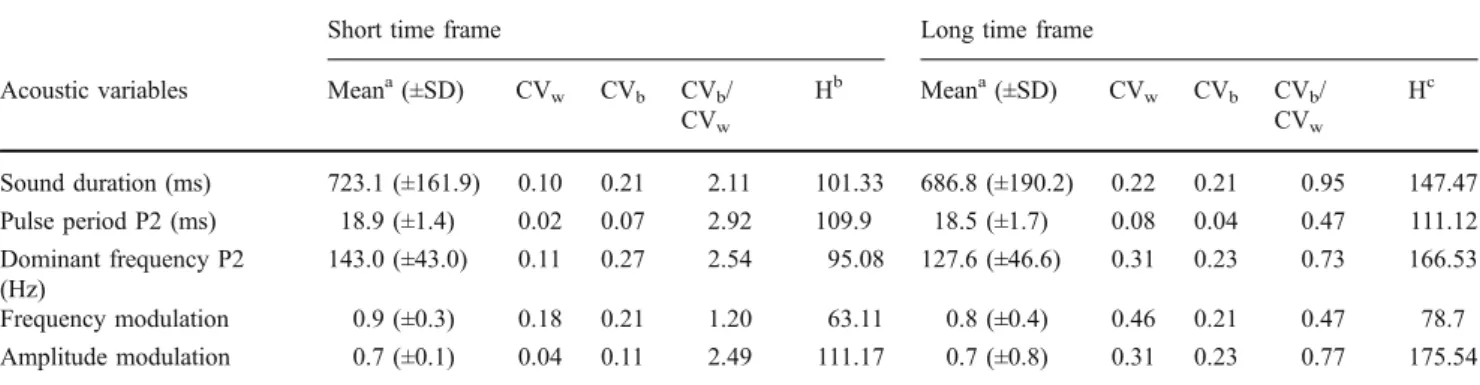
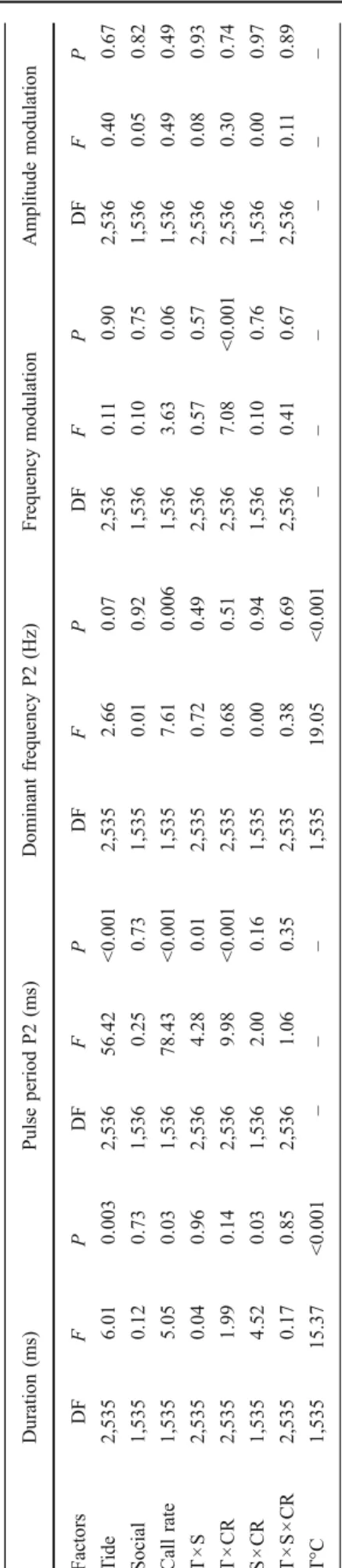
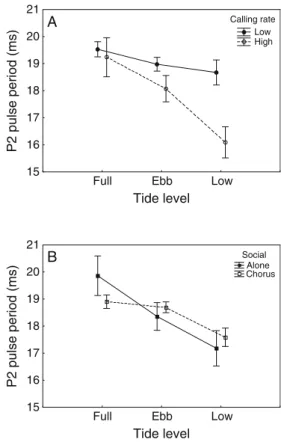
+2
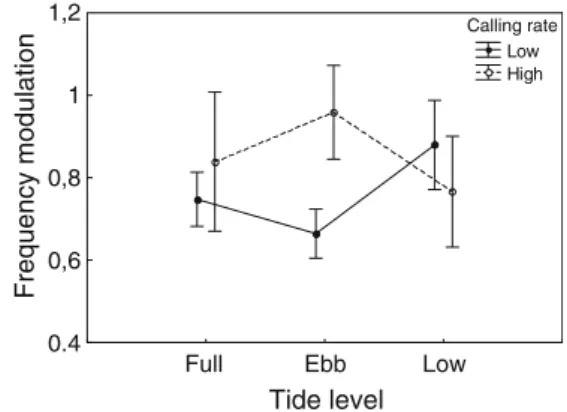
Documentos relacionados