A reverse transcription loop-mediated isothermal amplification assay optimized to detect multiple HIV subtypes.
Texto
Imagem
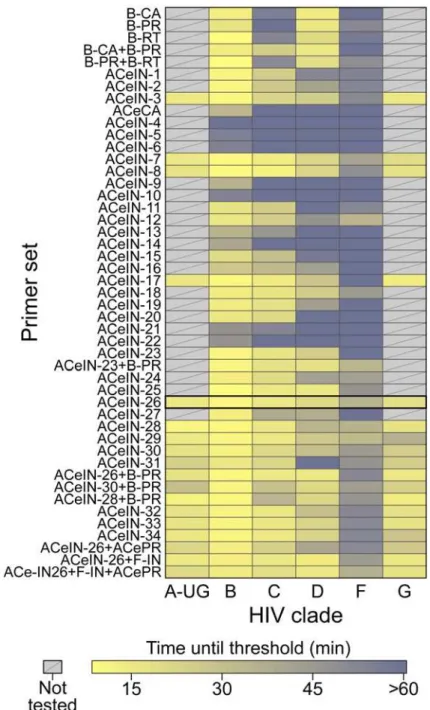
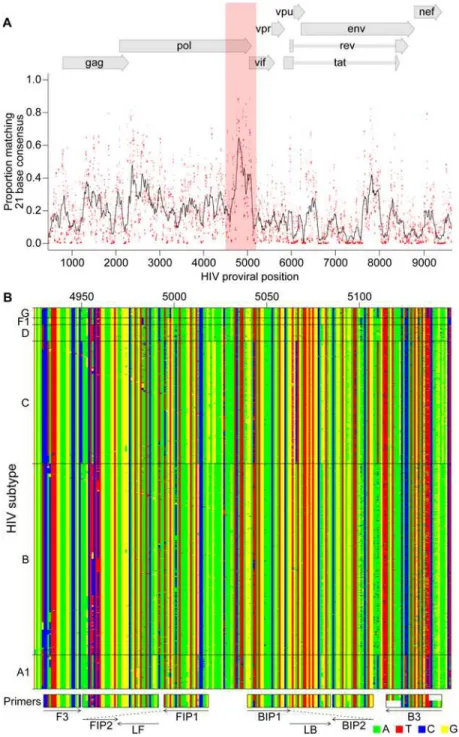
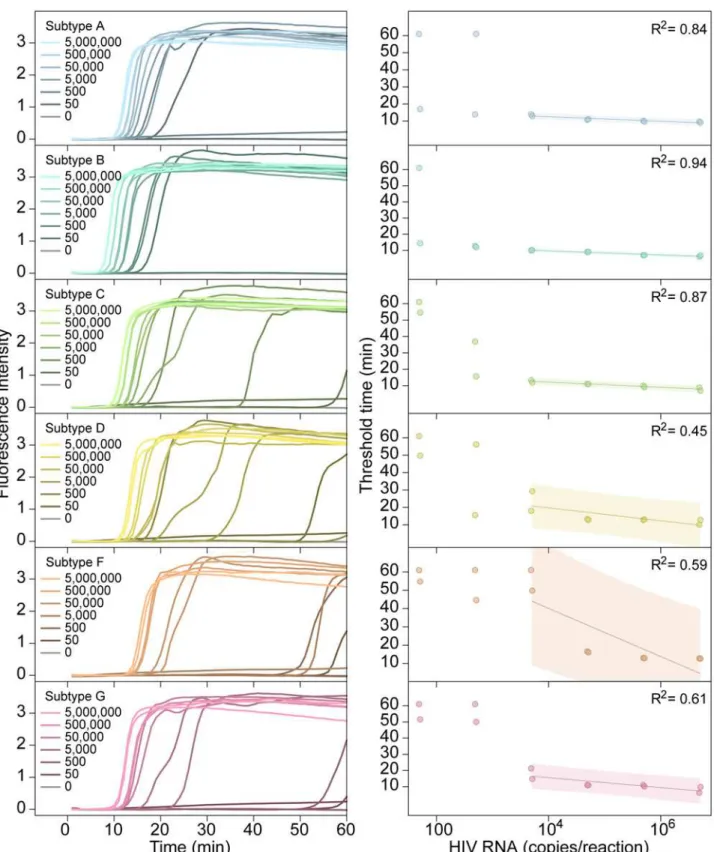
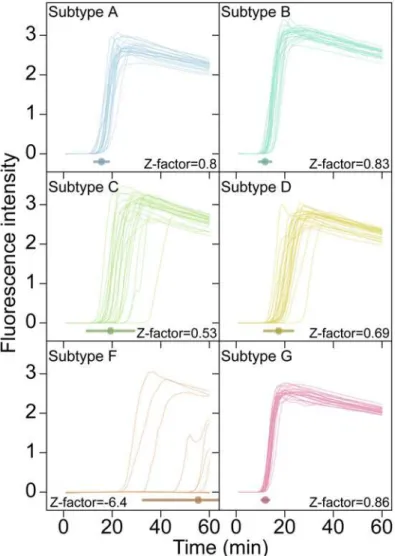
Documentos relacionados
From the 12 studies included (13 entries) in this meta-analysis, they all started from the same research assumption, in which the elderly with classifications according to the
Desse modo, o artigo apresenta um estudo sobre a deficiência visual, em espe- cífico a baixa visão em relação à iluminação artificial de ambientes internos, e busca.. apresentar
Comparison of the use of indirect immunofluorescence assay (IFA), immunochromatography assay (ICA-BD) and reverse transcription-polymerase chain reaction (RT-PCR) for
2: sensitivity of malachite green-loop-mediated isothermal amplification (MG-LAMP), polymerase chain reaction of minicircle kinetoplast DNA gene (PCR-mkDNA), and PCR-ITS1 to detect
A pre-enriched multiplex polymer- ase chain reaction (m-PCR) assay employing specific primers was developed and used to detect Salmonella at the genus level and to identify
These results showed that the TPC assay was capable to predict the ocular irritant potential of shampoos, and therefore was a promissory tool to be used as a preliminary assay
A loop-mediated isothermal amplification (LAMP) assay was developed for rapid, sensitive and specific detection of African swine fever virus (ASFV).. A set of LAMP primers
1 - A DIRCSI tem por missão planear, estudar, dirigir, coordenar e executar as atividades inerentes aos sistemas de informação e tecnologias de informação e
