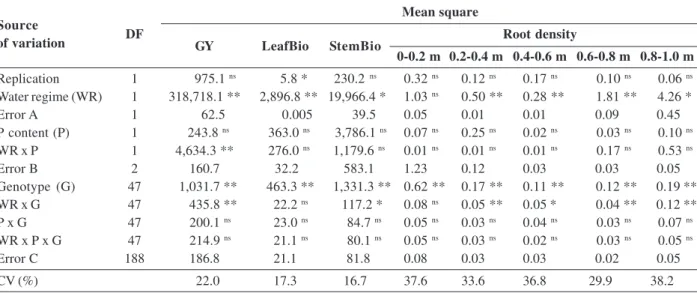
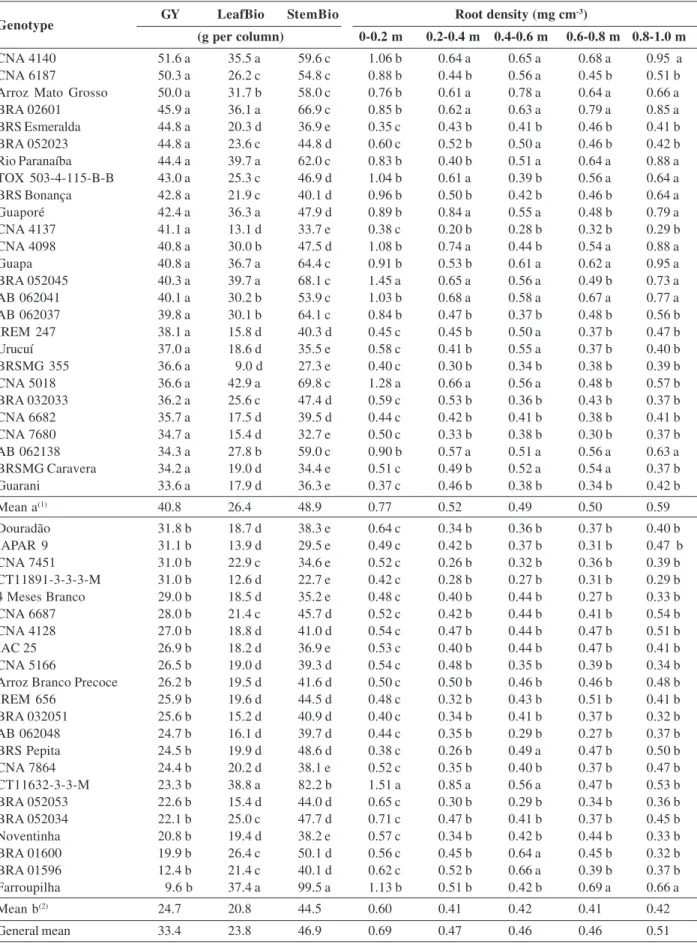
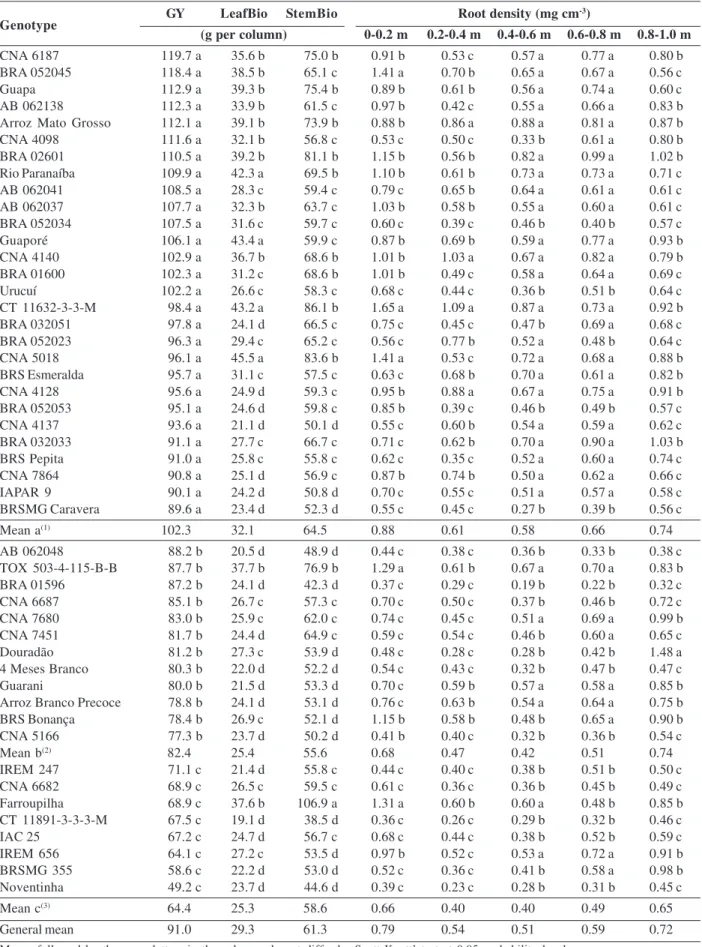
Performance of low phosphorus tolerant rice genotypes under drought stress
Texto
Imagem

Documentos relacionados
Para que se possa tentar direccionar com mais facilidade os sujeitos em estudos futuros, a fim de obtermos a valência das memórias, fica a proposta de alterar
A Figura 29 apresenta os resultados dos ensaios da concentração do rejeito de cromita para uma ampla faixa de tamanhos empregando-se um concentrador Falcon SB-40, onde foram
Regarding the importance of choice of rice cultivars tolerant to drought, the present study aimed to evaluate the traits related to productivity, in rice genotypes, under
O alargamento do campo de aplicação da denúncia espontânea no Direito Aduaneiro, proposto no texto legal para permitir a exclusão, também, da imposição de penalidades de
Contudo, apesar desta inovação, Freud ([1904/1905]1989) considerava o tratamento psicanalítico impróprio para os casos de psicose, mas não descartou que através
VI - apresentar as propostas de avaliação e atualização dos documentos com vistas a determinar sua destinação final, elaborando o Plano de Destinação de Documentos de Arquivo e
O projeto proposto para essas alunas do 6° semestre de Letras/Espanhol foi justamente unir vários saberes, contextualizando-o com a realidade da disciplina que é obter o profundo
Petrópolis fomos convencidos de que a prática não se apresenta salutar, por questões técnicas. Não somente técnicas, mas as que envolvem os Direitos Fundamentais”. Esse é um
