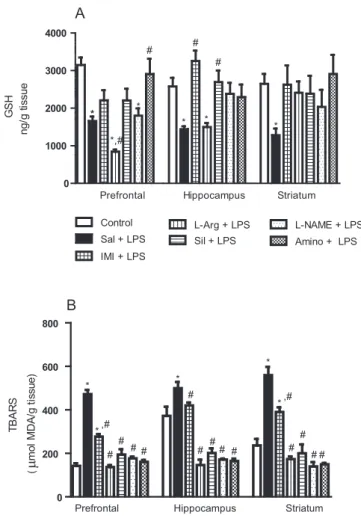
like effect of nitric oxide synthase inhibitors and sildenafil against depressivelike behavior in mice
Texto
Imagem
![Fig. 1. (A) Immobility time (s) in the forced swimming test (FST) and (B) number of crossings in the open field test (OFT) of animals treated with [ L -arginine (L-arg), sildenafil (sil), L -NAME, aminoguani-dine (amino)] or the antidepressant imipramine (I](https://thumb-eu.123doks.com/thumbv2/123dok_br/15361425.564949/4.892.467.813.627.1008/immobility-swimming-crossings-arginine-sildenafil-aminoguani-antidepressant-imipramine.webp)
![Fig. 2. Sucrose preference measured as% of sucrose consumption of animals treated with [ L -arginine (L-arg), sildenafil (sil), L -NAME, aminoguanidine (amino)] or the antidepressant imipramine (IMI) alone or prior to LPS administration](https://thumb-eu.123doks.com/thumbv2/123dok_br/15361425.564949/5.892.72.428.608.1011/sucrose-preference-consumption-sildenafil-aminoguanidine-antidepressant-imipramine-administration.webp)

Documentos relacionados
Abbreviations: 5-HT, serotonin; ACTH, adrenocorticotropic hormone; AMPA, α -amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; AMPK, AMP-activated protein kinase; BDNF,
Objective: To correlate neurotrophic factors – brain-derived neurotrophic factor (BDNF), glial cell line- derived neurotrophic factor (GDNF), and beta-nerve growth factor (beta-NGF)
Objectives: To compare levels of a marker of lipid peroxidation (thiobarbituric acid reactive substances, TBARS) and brain-derived neurotrophic factor (BDNF) in umbilical cord
The brain-derived neurotrophic factor (BDNF) gene Val66Met polymorphism affects memory performance in older adults.. Rev
Takeuchi, Role of nitric oxide in regulation of gastric acid secretion in rats: effects of NO donors and NO synthase inhibitor, Br. Kim, Effects of a nitric oxide donor and nitric
Inhibition of allergic airway inflammation in mice lacking nitric oxide synthase 2. Role of exhaled nitric oxide
Effect of intrathecal administration of nitric oxide/ cGMP/K ATP pathway inhibitors on the antinociception induced by acute aerobic exercise (AE) in the paw-withdrawal ( A ) and
Conclusions: Our results have demonstrated the presence of cyclic adenosine monophosphate-PDE and cyclic guanosine monophosphate-PDE in the human clitoris and may indicate a
