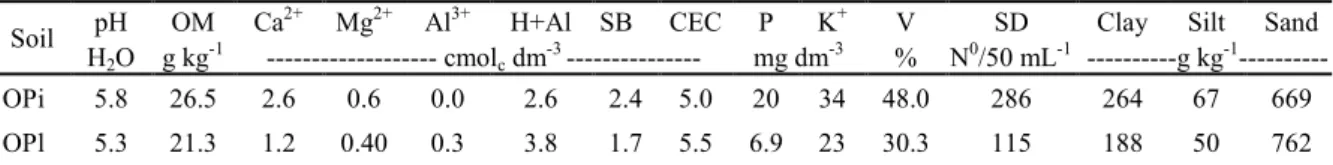
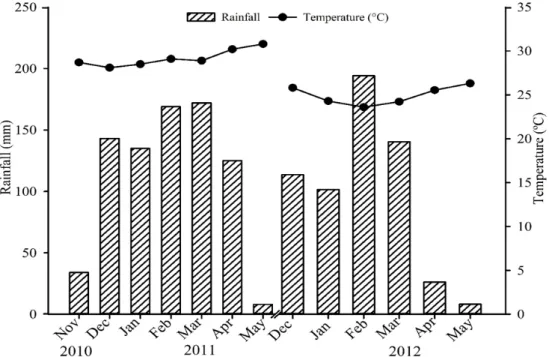
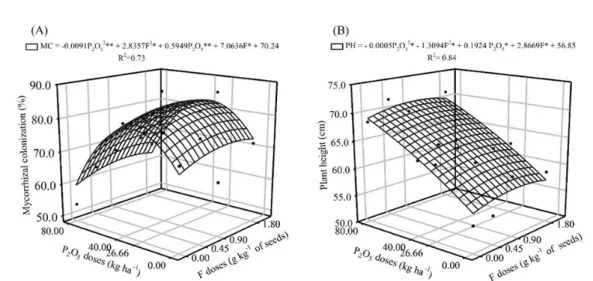
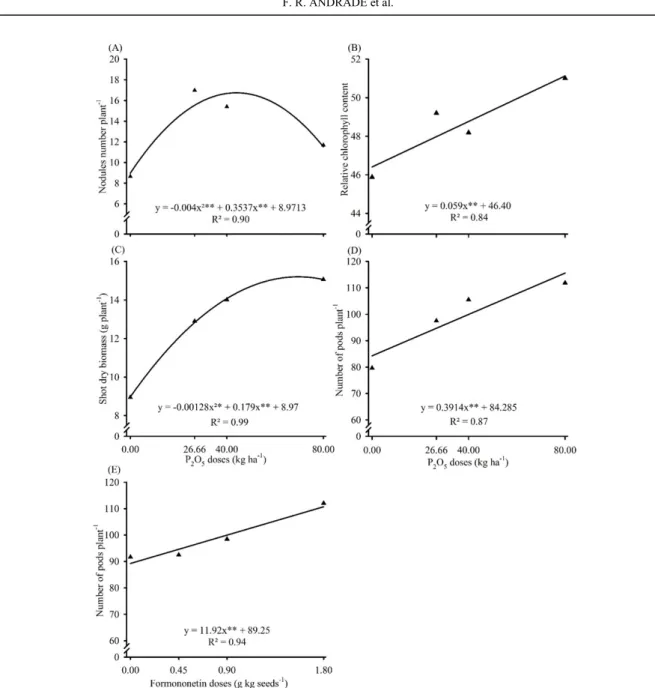
MYCORRHIZATION STIMULANT IN SOYBEAN ASSOCIATED WITH PHOSPHATE FERTILIZATION IN OXISOLS
Texto
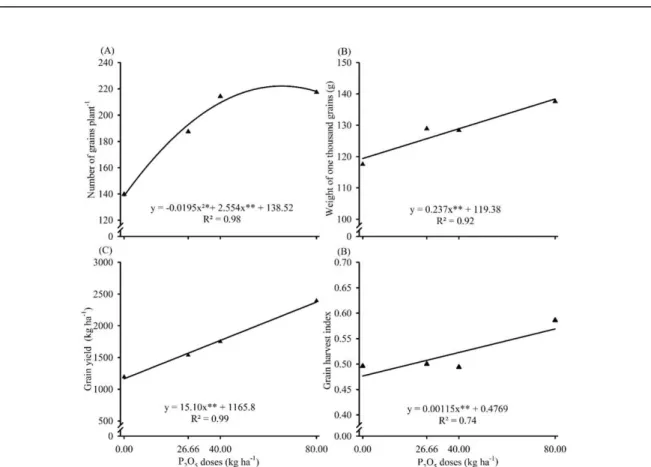
Imagem
Documentos relacionados
Aspects of morphology and productivity, such as plant height, root length, number of pods per plant, number of grains per pod, 100-grain weight, final grain yield and dry weight,
The nitrogen levels significantly affected dry weight, thousand grain weight and oil content in 2012, as well as plant height, number of pods, dry weight, grain yield and oil yield
Table 2 - Ryegrass dry mass and soybean plant population, number of pods per plantk, number of grains per plant, grain productivity, aerial part length (APL), root system
The following variables were measured: leaf N content, dry matter of plants, yield components (number of pods plant -1 , number of grains plant -1 , the average number of grains
GGE biplot vectors for the independent variables number of branches per plant (NBP), number of pods in branches per plant (NPPB), number of pods in stems per plant (NPPS), number
The results revealed that significantly higher shoot length, shoot fresh weight, shoot dry weight, root length, root fresh weigh, root dry weight, number of nodules per
Estimates of Pearson’s correlation coefficients of grain yield (GY) in relation to pod length, number of grains per pod, number of pods per plant, number of pods per area, and mass
Twenty-two QTLs were discovered for chlorophyll, leaf and stem fresh biomass, leaf biomass dry weight, leaf temperature, number of pods per plant, number of seeds per plant,
