Ana Rita Marques Lemos
68
0
0
Texto
Imagem


+7

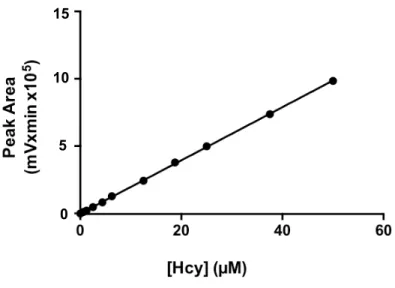
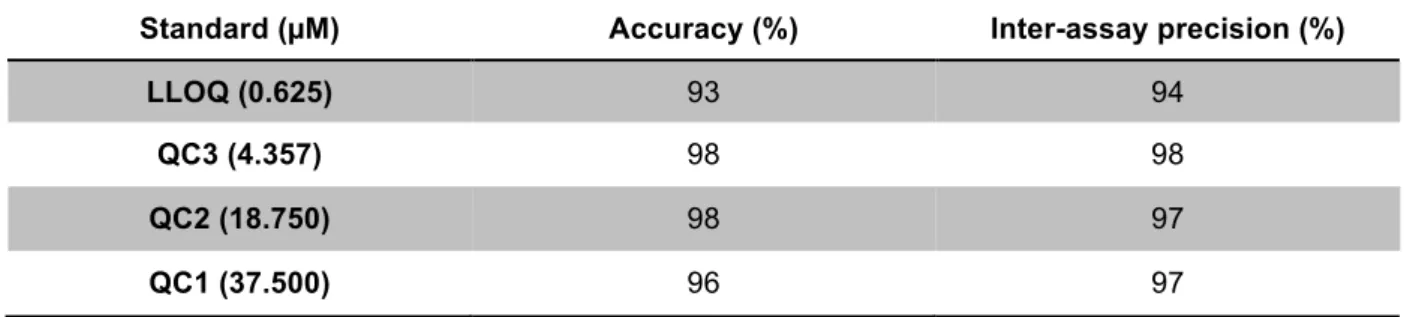
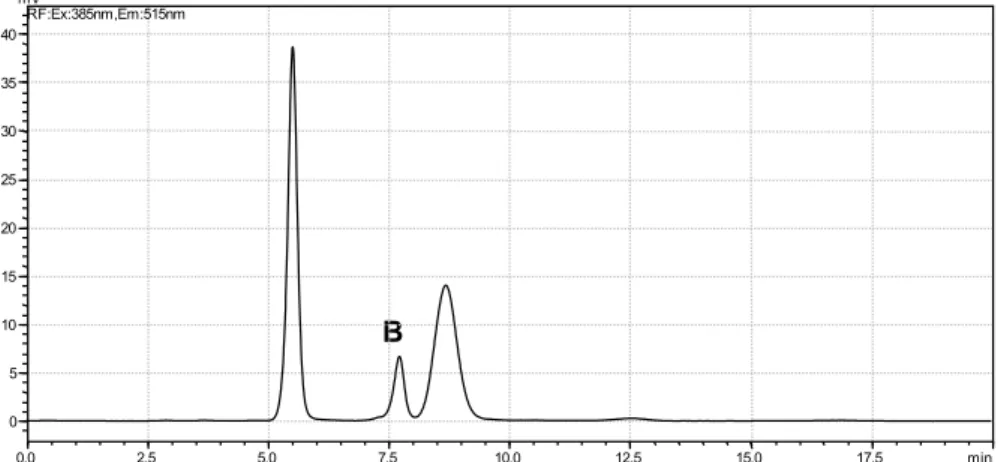
Documentos relacionados