AACL BI OFLUX
Aqu a cu lt u re , Aqu a riu m , Con se rv a t ion & Le gisla t ion
I n t e rn a t ion a l Jou rn a l of t h e Bioflu x Soci e t y
I n sit u a n d e x sit u p re se rva t ion of Pru ssia n ca rp
oocyt e s
M. Mehdi Taat i, Ali Shabani, and M. Reza I m anpoor
Depart m ent of Fishery, Gorgan Universit y of Agricult ural Sciences and Nat ural Resources, Gorgan, I ran. Corresponding aut hor: M. Taat i, Taat i.Mehdi@gmail. com
Abst ract. The decline of ova viabilit y aft er ovulation is one of the lim it ing fact ors in cont rolled reproduction of several fish especially in w arm w at er fish. The purpose of t his st udy w as evaluating the viabilit y of Prussian carp oocyt es ret ained w it hin t he ovarian cavit y (in sit u st orage) and out side of the ovarian cavit y ( ex situ st orage) at 21ْC and 4 ْC for 20, 30, 60 and 120 m in. Experim ent s w ere perform ed w ith 20 fem ale fish ( 10 fem ales for each experim ent al group) Carassius gibelio ( Bloch, 1782) induced t o spawn with 0.5 m g kg-1 Ovaprim ( sGnRH+ Dom pridon) . Prussian carp oocyt es could be successfully
st ored for 2 h in situ at 21ْC and 4ْC w it hout significant decrease in eyed em bryo and hatching rat e ( P> 0.05) . The percent age of eyed em bryo in kept eggs out side t he ovarian cavit y at 21ْC and 4ْC for all durations w as significantly different com pared t o cont rol group ( P< 0.05) . There w as not significantly different in deform ed larvae percent during in situ and ex sit u st orage at 21 ْC and 4 ْC as com pared with cont rol.
Key W ords: oocyt e, in sit u, ex sit u st orage, Prussian carp.
I n t rod u ct ion. Fish sperm at ozoa from various species have been successfully
cryopreserved, but cryopreservat ion has not been successful for unfert ilized eggs or fert ilized eggs at any st age of developm ent because of t heir large size, com plexit y, yolk cont ent and relative im permeabilit y t o cryoprot ect ant s ( Zhang et al 1993) . Oocyte viabilit y refers t o t he t ime period during which fert ilizat ion of ovulat ed oocyt es rem ains possible once t hey have been em it t ed by t he fem ale (Legendre et al 1996) . The fert ilizabilit y of m at ure ovulat ed ova gradually decrease and t hen disappears altoget her when t hey are ret ained in t he body cavit y of t he fem ale, kept in coelom ic fluid or in wat er, or under in vit ro condit ions ( Rot hbard et al 1996) . Fert ilit y of fish gam et es undergoing ext ernal fert ilizat ion is lim ited t o a few seconds or m inut es in wat er because oocyt es are act ivated and undergo cort ical react ion leading t o m icropyle closure (Hart 1990) . Short t erm preservat ion aim ed at increasing post spawning gam ete longevit y m ay im prove hat chery m anagement , m inim izing problem s result ing from inbreeding and asynchronous brooder m at urat ion (Brom age & Robert s 1995) . Ovulated oocyt es retained in t he ovarian cavit y undergo over ripening due t o gradual m orphological and biochem ical changes t hat negat ively affect fert ilit y and larval development ( Form acion 1993) . As over- ripening of eggs will inevit ably event ually occur in fish bloodst ocks, it is im port ant t o obt ain and fert ilize eggs at t he correct t im e aft er ovulat ion. I n fish hat cheries, over ripening represent s a problem when ovulated eggs are not st ripped on t im e, especially for t hose fish whose eggs can only be obt ained by m anual st ripping and fert ilized art ificially ( Kjorsvik et al 1990) . Unlike salm on and t rout , oocyt es of t he few warm wat er fishes exam ined t o date lose t heir viabilit y wit hin a few hours of st orage in t he ovarian cavit y ( Espinach et al 1984; Rizzo et al 2003) .
Aft er ovulat ion, t he ova rem ain arrest ed in met aphase I I of m eiosis unt il t hey becom e act ivated. Act ivat ion rem oves t his dorm ant st age and is associated wit h a num ber of physiological changes ( Ginsburg 1968; Linhart et al 1995b) . The post -ovulat ion st orage of ova in t he ovarian cavit y ( in vivo) of com m on carp Cyprinus carpio
Jahnichen 1981; Marcel 1981) . Ova st ripped aft er ovulat ion and st ored in vit ro progressively lose t heir fert ilit y aft er 4–6 h at t em perat ures of 15–20° C (Jahnichen 1978; Kiselev 1980; Marcel 1981; Zlabek & Linhart 1987) . Koi carp (Cyprinus carpio Linnaeus, 1758) ova st ored wit hout an ext ender for 6 h at a st able tem perat ure of 20° C had m ore t han 50% hat ching rat e ( Rot hbard et al 1996) . The fert ilit y of t ench Tinca t inca
( Linnaeus, 1758) ova st ored in vit ro for 1, 5. 5 and 8. 5 h at 13ْC significant ly decreased t o 45. 4, 10. 1 and 0. 6% , respect ively, as calculat ed by hat ching rat e (Linhart & Billard 1995a) .
The aim of t his st udy was to evaluat e t he effect of in sit u and ex sit u Prussian carp ova st orage on it s eyed em bryo and hat ching capacit y and m alform ed larvae percent .
M a t e ria l a n d M et h od. Broodst ocks ( 15 m at ure m ales and 20 m at ure females) were
obt ained from reared hat chery at Nahar Khoran, Gorgan, during t he spawning season of Prussian carp and t ransferred t o t he place of experim ent and acclim ated for 2 weeks in 1000 L t anks. The experim ent was carried out at t he aquacult ure Cent er of Agricult ural Science and Nat ural Resources, Universit y of Gorgan, I ran.
Ga m e t e s colle ct ion. To st im ulat e fish, we inj ected int raperit oneally: 0. 5 m L kg-1
b. w. Ovaprim ( sGnRHa+ dom pridon). Milt sam ples were collected during t he 2010 spawning season from 15 sexually m at ure t wo- years- old m ale Prussian carps ( m ean TW: 43. 6 ± 9. 84 g) . Sem en sam ples were collect ed by m assage from t he ant erior port ion of t he t est is towards t he genit al papilla. Care was t aken t o avoid cont am inat ion of t he sem en wit h wat er, m ucus, blood cells, faeces or urine. Sem en of each m ale was collect ed by using 2 cc polyet hylene syringes and sperm bat ches were st ored in a refrigerat or at 4ْC unt il use for fert ilizat ion. At fert ilizat ion, sperm of all m ales was pooled in t he sam e proport ions by volum e as t hat obt ained by st ripping. These m ixt ures were used for t he fert ilizat ion assay.
Also, fem ales ( m ean TW: 57. 23 ± 8. 61 g) were inj ect ed wit h 0. 5 m g kg- 1 Ovaprim
and 12 hours aft er inj ect ion t reat m ent half of t hem were st ripped and t heirs eggs were collect ed in a bowl. The ot her half fem ales ( 10 fem ales) were kept in t ank at 21° C ( 5 fem ales) and in refrigerat or at 4° C ( 5 fem ales) .
St ora g e of ov u la t e d oocy t e s. Recent ly obtained eggs and females were
subj ect ed t o one of t he following st orage t reat m ent s:
- Group 1 ( in sit u storage at 21° C) : 5 ovulat ed, not st ripped fem ales ( i. e. wit h t heir oocyt es ret ained in t he ovarian cavit y) were kept t oget her in one 300 L t ank wit h aerat ion and were st ripped 20, 30, 60 and 120 m in aft er ovulat ion.
- Group 2 ( in sit u st orage at 4° C) : 5 ovulat ed, not st ripped fem ales were kept in 2 aquarium s ( wit h 80 L aerat ed water) and were st ripped 20, 30, 60 and 120 m in aft er ovulat ion.
- Group 3 ( ex sit u st orage at 21° C and 4° C) : The all of fem ale ova were pooled. This t reat m ent had 8 t reat m ent s and t hree replicates for each t reat m ent . For each replicate 300 m g ( about 300) eggs were placed in Pet ri dish ( 10 cm diam eter) . Half of Pet ri dishes w e re pla ce d i n t o a re f ri ge ra t o r ( e xp osu re t o 4 º C) a nd h al f o f t he m h el d a t 2 1 º C. - Cont rol: Bat ches of about 300 ova ( 300 × 3 replicates) were fert ilized im m ediat ely aft er egg collect ion and was considered as cont rol group.
Fe rt iliz at ion a n d in cu b a t ion. Aft er storage for above t ime in t he t wo t est
t em perat ures and in sit u and ex sit u condit ions, undiluted sperm ( 0. 02 m L) was added to oocyt es, and water ( 10 m L) at am bient t em perat ure ( 21° C) was used t o act ivate t he gam et es, allowing fert ilizat ion and adhesion of eggs t o t he Pet ri dishes, t hen t ransferred t o aquarium ( cont aining 80 lit ers aerat ion water) and were placed in t he bot t om of t hem. The eggs were placed in a m onolayer t o ensure adequat e oxygen supply. The percent age of eyed em bryos, hat ching rate and percent age of m alform ed larvae were recorded.
St a t ist ica l a n a ly sis. Dat a analysis of variance ( t wo- way ANOVA) was done wit h
a significance level of P< 0. 05. Before analysis by ANOVA, dat a was used for norm alit y of dat a dist ribut ion and hom ogeneit y of variance. Result s are present ed as m ean ± SD. St at ist ical analyses were perform ed wit h SPSS 16 for windows st at ist ical package.
Re su lt s a n d Discu ssion
Effe ct s of e x sit u a n d in sit u st ora g e on e y e d e m b ry o a n d h a t ch in g ra t e. I n
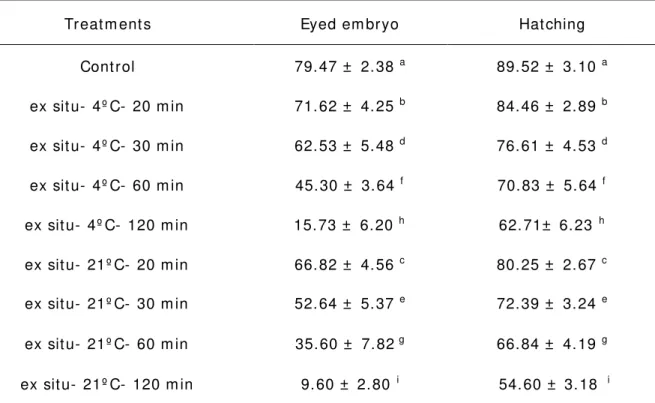
general, t here was a significant decline ( P< 0: 05) in t he rat e of eyed em bryo and hat ching rat e over t he period of ex sit u oocyt e st orage. Result s showed 64% decline in t he rat e of eyed em bryo at 4° C and 70% decrease at 21° C as com pared wit h cont rols aft er 120 m in in t his experim ent al group. Also, at 60 m in, t he eyed em bryo rat e had decreased by over 19% at 4 ° C and 23% at 21° C ( Table 1) . The percent age of hat ching of st ored ova in ex sit u condit ion decreased wit h increasing t im e at 4° C and 21 ° C ( P> 0. 05) . Tem perat ure had significant ly effect on eyed em bryo and hat ching rate; so t hat t hese param eters at 4° C were greater t han 21° C ( Table 1).
Table 1 Effect s of ex sit u ( out of ovarian cavit y) st orage, t em perat ure and t im e on
eyed em bryos and hat ching percent ( Mean ± SD) in Prussian carp ova (C. gibelio)
Treat m ent s Eyed em bryo Hat ching
Cont rol 79. 47 ± 2. 38 a 89. 52 ± 3. 10 a
ex sit u- 4º C- 20 m in 71. 62 ± 4. 25 b 84. 46 ± 2. 89 b
ex sit u- 4º C- 30 m in 62. 53 ± 5. 48 d 76. 61 ± 4. 53 d
ex sit u- 4º C- 60 m in 45. 30 ± 3. 64 f 70. 83 ± 5. 64 f
ex sit u- 4º C- 120 m in 15. 73 ± 6. 20 h 62. 71± 6. 23 h
ex sit u- 21º C- 20 m in 66. 82 ± 4. 56 c 80. 25 ± 2. 67 c
ex sit u- 21º C- 30 m in 52. 64 ± 5. 37 e 72. 39 ± 3. 24 e
ex sit u- 21º C- 60 m in 35. 60 ± 7. 82 g 66. 84 ± 4. 19 g
ex sit u- 21º C- 120 m in 9. 60 ± 2. 80 i 54. 60 ± 3. 18 i
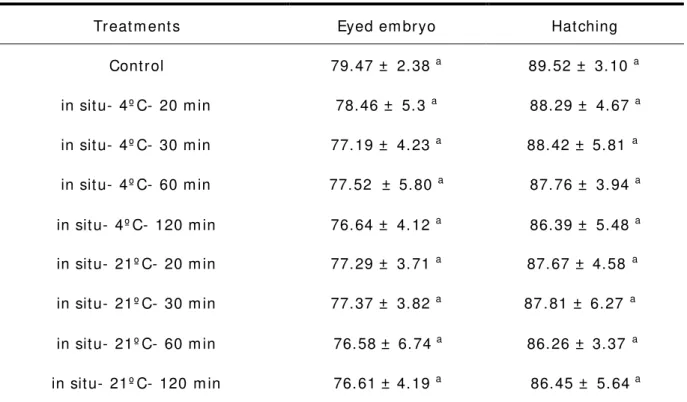
I n oocyt es st ored in sit u at 21° C and 4° C, t he eyed em bryo and hat ching percent at 20, 30, 60 and 120 m in of st orage were stat ist ically sim ilar t o t hose of t he cont rols ( Table 2). I n sit u st orage at 21° C and 4° C had not drast ic reduct ion in eyed em bryo and hat ching rat es as com pared wit h cont rol ( Fig. 2).
Effe ct of e g g st ora g e in sit u a n d e x sit u on m a lform e d la rv a e ra t e . The
Table 2 Effect s of in sit u ( in ovarian cavit y) st orage, t em perat ure and t im e on
eyed em bryos and hat ching percent ( m ean ± SD) in Prussian carp ova (C. gibelio)
Fig. 1. Larvae m alform ation in ex sit u st orage of Prussian carp eggs.
Fig. 2. Larvae m alform ation in t he case of in sit u st orage of Prussian carp eggs.
Treat m ent s Eyed em bryo Hat ching
Cont rol 79. 47 ± 2. 38 a 89. 52 ± 3. 10 a
in sit u- 4º C- 20 m in 78. 46 ± 5. 3 a 88. 29 ± 4. 67 a
in sit u- 4º C- 30 m in 77. 19 ± 4. 23 a 88. 42 ± 5. 81 a
in sit u- 4º C- 60 m in 77. 52 ± 5. 80 a 87. 76 ± 3. 94 a
in sit u- 4º C- 120 m in 76. 64 ± 4. 12 a 86. 39 ± 5. 48 a
in sit u- 21º C- 20 m in 77. 29 ± 3. 71 a 87. 67 ± 4. 58 a
in sit u- 21º C- 30 m in 77. 37 ± 3. 82 a 87. 81 ± 6. 27 a
in sit u- 21º C- 60 m in 76. 58± 6. 74 a 86. 26 ± 3. 37 a
Effe ct of t im e of st ora g e, h old ing con d it ion a nd t e m p e ra t u re on ey ed
e m b ry o, h a t ch ing a n d m a lform e d la rv a e p e rce n t. Result s shown t hat t he effect of
ex sit u st orage t im e on eyed em bryos and hat ching rate was significant . I n t his st at us, st orage tem perat ure had significant effect on eyed egg rate ( P< 0. 05) . Eyed egg and hat ching rat e decreased wit h increasing t im e ( 20, 30, 60 and 120 m in) but m alform ed larvae rat e had not changes wit h t im e in all t reat m ent s. Highest eyed em bryo and hat ching rat e bet ween t reat ment s of ex sit u st orage was observed in 20 m in st orage and 4° C and lowest eyed em bryo and hat ching rat es was observed in 12 0 m in st orage at 2 1
ْ
C. Also, any significant difference was not observed in eyed egg, hat ching and m alform ed larvae percent in ovarian cavit y st orage at 21° C and 4° C and 20, 30, 60 and 120 m in st orage.
I n t his work, t he viabilit y of C. gibelio was assessed t hrough eyed em bryo and hat ching rates evaluat ed aft er in sit u and ex sit u st orage. Percent age of eyed eggs and hat ching in ex sit u holding decreased rapidly as st orage t ime increased and it was recorded close t o 9. 6% , 54. 6% and 15% , 62% aft er 120 m in ex sit u st orage at 21° C and 4° C respect ively, sim ilar t o report s in several teleost species (Mollah & Tan 1983; Espinach et al 1984; Linhart & Billard 1995b; Linhart et al 2001; Rizzo t e al 2003) . Thus, in our experim ent s, C. gibelio oocyt es could only be successfully st ored for 1 h in ex sit u t reat m ent at 4° C. The highest eyed eggs and hat ching rat e were obtained when st ripping and fert ilizat ion were perform ed im m ediately aft er ovulat ion, as in t he Rham dia sapo
( Valenciennes, 1840) Prochilodus platensis ( Valenciennes, 1837) and Prochilodus
m arggravii ( Walbaum , 1792) ( Espinach et al 1984; Fort uny et al 1988; Rizzo et al 2003).
Anot her st udy on storage of com m on carp ova (Rot hbard et al 1996; Urbanyi et al 1998), wit hout any ext ender, showed a t im e- relat ed negat ive effect of decreased t em perat ure on fert ilizat ion and hat ching rat e. However, variabilit y of ova m ay be relat ed t o t he healt h of t he fem ale, t im e of ovulat ion, t em perat ure, feeding and qualit y of wat er ( Horvat h 1978; Linhart & Billard 1995a). A decrease in tem perat ure had a posit ive effect on t he fert ilizing potent ial of eggs t hrough t im e in t his experim ent . I n anot her st udy using com m on carp eggs ( Urbanyi et al 1998) and no ext ender, a decrease in t em perat ure had a negat ive effect on fert ilizat ion capacit y and hat ch t hrough t im e. These result s were not agreed wit h our st udy.
More research on st orage has been done wit h eggs of coldwat er fishes t han wit h eggs of warm wat er fishes. Eggs of chum salm on Oncorhynchus ket a ( Walbaum , 1792) had greater viabilit y when st ored undilut ed ( wit hout addit ion of ext ender solut ions) at 9 t o 15° C t han at 3 t o 6° C ( Jensen & Alderdice 1984) . Of t he warm wat er fishes, eggs of European cat fish Silurus glanis (Linnaeus, 1758) st ored undilut ed at 19° C for 3. 5 h had greater hat ching ( 54% ) t han did cont rol eggs st ored for 0 h ( 35% ) , while eggs st ored at 8° C yielded no hat ching (Linhart & Billard 1995b) . Eggs of com m on carp st ored bet ween 0 and 24° C for 60 m in wit hout an ext ender exhibit ed t im e dependent and t em perat ure-dependent sensit ivit y ( Urbanyi et al 1998) .
I n our observat ions ( Table 2) eyed em bryo and hat ching rate in ovarian cavit y st orage is bet t er t han ex sit u holding and offer a potent ial for ext ending t he st orage t ime of Prussian carp ova. Alt hough, st orage t ime can increase if we use of low t em perat ure.
These result sdid not confirm t he result s of Rizzo et al ( 2003) in Prochilodus m arggravii. Kj orsvik et al ( 1990) report ed t hat aft er st ripping, fish eggs gradually undergo changes sim ilar t o over ripening. On t he ot her hand, Form acion et al ( 1993) st ated t hat oocyt e over ripening m ay be evaluat ed on t he basis of t he decline in fert ilizat ion and hat ching rat es. I n addit ion, Lahnst einer ( 2000) report ed t hat eyeing rate in rainbow t rout eggs, ret ained inside of body cavit y, decreased wit h t he progress of over ripening process. Therefore, decline of eyed eggs may be considered as a sign of progress of over
( present st udy) had not significant different in all of keeping t im e t hat was not sim ilar t o findings in t he cat fish R. sapo ( Espinach et al 1984) and P. marggravii (Rizzo et al 2003). Morphological and biochem ical alterat ions during oocyt e over ripening m ay result from breakdown of yolk prot eins, loss of sm all organic m olecules t hrough oocyt e m em branes, and dephosphorylat ion of prot eins and lipids (Craik & Harvey 1984) . I n carps, a dist urbance in t he aerobic respirat ion process occurs t hat leads t o t he product ion of lact ic acid, which accum ulat es in t he ovarian fluid and reduces it s pH, wit h ult im at e loss of oocyt e m em brane int egrit y (Linhart et al 1995) . Alt hough t he eye eggs and hat ching rate
in C. gibelio ( in ovarian cavit y st orage) were not lower t han cont rol group aft er 2 h
st orage.
Con clu sion s. Result s shown t hat st orage of Prussian carp art ificial ova out of ovary and
wit hout ext ender is not very appropriat e for short - t erm st orage of Prussian carp eggs, but st orage of oocyt es of t his species in ovarian cavit y is appropriat e for longer durat ion t han 20, 30, 60 and 120 m in st orage. Also, m aint ain Pr ussian carp ova at 4° C had percent age of eyed em bryo and hat ching great er t han 21° C in ex sit u st orage. St orage durat ion out of ovary had significant effect on hat ching perform ance, but holding st at us ( ex sit u and in sit u) , t em perat ure and t im e had not significant ly influence on m alform ed larvae.
Re fe re n ce s
Brom age N. R. , Robert s R. J., 1995 Preservat ion of gamet es. I n: Brom age N. R. , Robert s R. J. , (eds), Brodst ock m anagem ent and egg and larval qualit y. Blackwell, Oxford. P. 53–75.
Craik J. C. A. , Harvey S. M. , 1984 Biochem ical changes associat ed wit h over ripening of t he eggs of rainbow t rout Salm o gairdneri Richardson. Aquacult ure 3 7: 347–57. Espinach R. A. , Am ut io V. G., Mest re Arceredillo J. P., Ort i G. , Nani A., 1984 I nduced
breeding of t he Sout h Am erican cat fish Rham dia sapo (C. & V. ) . Aquacult ure
3 7: 141–146.
Form acion M. J., Hori R., Lam T. J. , 1993 Overripening of ovulat ed eggs in goldfish. I . Morphological changes. Aquacult ure 1 1 4: 155–168.
Fort uny A. , Espinach R. A. , Am ut io V. G. , 1988 Horm onal induct ion of final m at urat ion and ovulat ion in t he sabalo, Prochilodus platensis Holm berg: t reat m ent s, latency and incubat ion t im es and viabilit y of ovules ret ained in t he ovary aft er ovulat ion.
7 3: 373–381.
Ginsburg A. S. , 1968 [ Fert ilization in Fishes and t he Problem of Polysperm y] . Nauka, Moscow. [ I n Russian]
Hart N. H., 1990 Fert ilizat ion in t eleost fishes: mechanism s of sperm –egg int eract ions. I nt Rev Cyt ol 1 2 1: 1–66.
Holcom b M., Cloud J. C. , I ngerm ann R. L. , 2005 I m pact of bact eria on short term st orage of salm onid eggs. Aquac Res 3 6: 1555–1561.
Horvat h L. , 1978 Experience in propagat ion of t he com m on carp (Cyprinus carpio L.) out t he spawning season. Aquacul Hungarica ( Szarvas) 1: 66–72.
Jahnichen H. , 1978 Unt ersuchungen uber Veranderungen der Qualit at ovuliert er Eier bei der kunst lichen Verm ehrung von Karpfen (Cyprinus carpio L.) . Zeit schrift fur die Binnenfischerei DDR 2 5: 23–25.
Jahnichen H., 1981 Kurzeit konservierung von Karpfensperm a als weit ere Moglichkeit der Rat ionalisierung der kunst lichen Vermehrung von Karpfen (Cyprinus carpio L.). Zeit schrift fur die Binnenfischerei DDR 2 8: 244–245.
Jensen J. O. T. , Alderdice D. F. , 1984 Effect of t em perat ure on short - term st orage of eggs and sperm of chum salm on (Oncorhynchus ket a) . Aquacult ure 3 7: 251- 265. Kiselev J. V. , 1980 [ The Biological Background of Fert ilizat ion and I ncubat ion of Fish
Eggs] . Naukova Dum ka, Kiev. [ I n Russian]
Kj orsvik E. , Mangor- jensen A., Holm fj ord I . , 1990 Egg qualit y in fishes. Adv Mar Biol
Lahnst einer F. , 2000 Morphological, physiological paramet ers charact erizing t he overripeningof rainbow t rout eggs. Fish Physiol Biochem 2 3: 107–118.
Legendre M. , Linhart O., Billard R., 1996 Spawning and m anagem ent of gam et es, fert ilized eggs and em bryos in Siluroidei. Aquat Living Resour 9: 59–80.
Linhart O. , Billard R., 1995a Biology of gam et es and art ificial reproduct ion in com m on t ench (Tinca t inca L. ). A review. Polskie Archivum Hydrobiologii 4 2: 37–56.
Linhart O. , Billard R. , 1995b Survival of ovulat ed oocyt es of t he European cat fish (Silurus
glanis) aft er in vivo and in vit ro st orage or exposure t o saline solut ions and urine.
Aquat Living Resour 8: 317–322.
Linhart O. , Kudo S., Billard R., Slecht a V. , Mikodina E. V. , 1995 Morphology, com posit ion and fert ilizat ion of carp eggs: a review. Aquacult ure 1 2 9: 75–93.
Linhart O., Gela D. , Rodina M., Rodriguez Gut ierrez M. , 2001 Short - t erm st orage of ova of com m on carp and t ench in ext enders. J Fish Biol 5 9: 616–623.
Marcel J. , 1981 Cont role de la Reproduct ion et Gest ion des Gam etes de Quelques Especes de Poissons Teleost eens. Diplom e, EPHE, Paris.
Mollah M. F. A., Tan E. S. P., 1983 Viabilit y of cat fish (Clarias m acrocephalus Gunt her) eggs fert ilized at varying post -ovulat ion t im es. J Fish Biol 2 2: 563–566.
Rizzo E. , Godinho H. P., Sat o Y. , 2003 Short - t erm st orage of oocyt es from t he neot ropical t eleost fish Prochilodus m arggravii. Theriogenology 6 0: 1059- 1070.
Rot hbard S. , Rubinst hein I . , Gelman E. , 1996 St orage of com m on carp (Cyprinus carpio
L. ) eggs for short durat ions. Aquac Res 2 7: 175–181.
Springat e J. R. C. , Brom age N. R. , Elliot J. A. K., Hudson D. L., 1984 The t im ing of ovulat ion and st ripping and t heir effect s on t he rates of fert ilizat ion and survival to eyeing, hat ch and swim- up in t he rainbow t rout (Salm o gairdneri R.) . Aquacult ure
4 3: 313–322.
Suzuki R. , 1980 Durat ion of developm ent al capabilit y of carp ova in t he ovarian cavit y aft er ovulat ion. Bullet in of t he Nat ional Research I nst it ut e of Aquacult ure 1: 1–6. Urbanyi B. , Baranyai B. , Dinnyes A., 1998 Chilling sensit ivit y of non- act ivat ed carp
(Cyprinus carpio) ova. Theriogenology 4 9: 175.
Zhang T. , Rawsen D.M. , Morris G.J. , 1993 Cryopreservat ion of pre- hat ch em bryo of zebrafish (Brachydanio rerio) . Aquat Living Resour 6: 145- 153.
Zlabek A., Linhart O. , 1987 [ Short - t erm storage of non- insem inat ed and unfert ilized ova of t he com m on carp, grass carp and silver carp] . Bullet in VURH Vodnany 2 3: 3–11. [ in Czech, wit h English sum m ary]
Received: 02 April 2011. Accept ed: 15 May 2011. Published online: 02 August 2011. Authors:
Moham m ad Mehdi Taati, Departm ent of Fishery, Gorgan Universit y of Agricultural Sciences and Nat ural Resources, I ran, Golest an, Gorgan, Shahid Behesht i Avenue, Post al code: 49138-15739, e-m ail: t aat i.m ehdi64@gm ail.com
Ali Shabani, Departm ent of Fishery, Gorgan Universit y of Agricult ural Sciences and Natural Resources, I ran, Golest an, Gorgan, Shahid Behesht i Avenue, Post al code: 49138- 15739.
Moham m ad Reza I m anpoor, Departm ent of Fishery, Gorgan Universit y of Agricultural Sciences and Natural Resources, I ran, Golest an, Gorgan, Shahid Behesht i Avenue, Post al code: 49138-15739.
How t o cite this art icle: