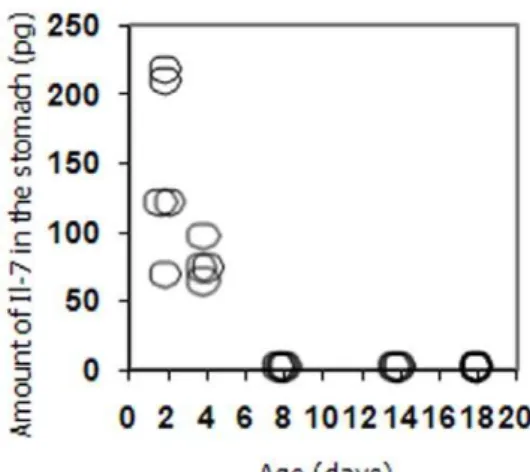
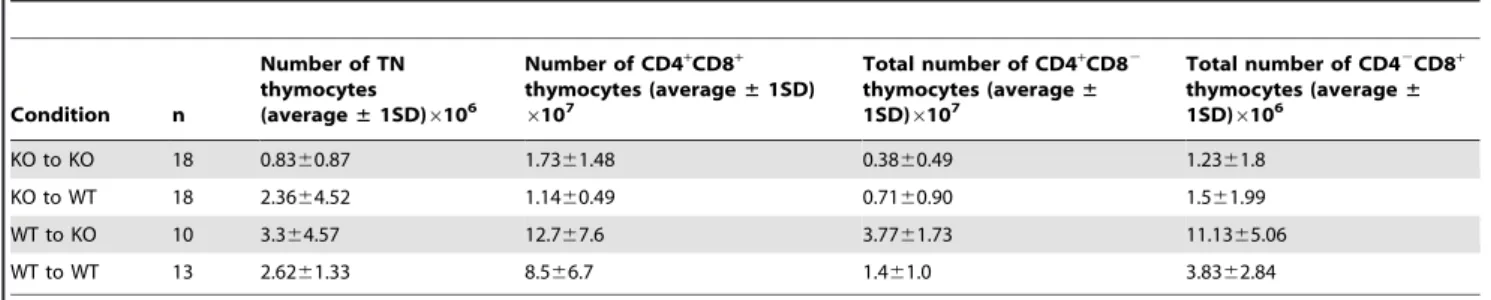
Interleukin 7 from maternal milk crosses the intestinal barrier and modulates T-cell development in offspring.
Texto
Imagem


Documentos relacionados
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Atividades dos homens e da igreja, se dão ao longo da história e em certo contexto. Sem considerarmos a historicidade dos homens e das instituições por elas
No caso e x p líc ito da Biblioteca Central da UFPb, ela já vem seguindo diretrizes para a seleção de periódicos desde 1978,, diretrizes essas que valem para os 7
Conheceremos nesta unidade os tipos de modais, suas infraestruturas, seus riscos, suas vantagens e desvantagens, assim como o papel da embalagem e da unitização para redução
Peça de mão de alta rotação pneumática com sistema Push Button (botão para remoção de broca), podendo apresentar passagem dupla de ar e acoplamento para engate rápido
Extinction with social support is blocked by the protein synthesis inhibitors anisomycin and rapamycin and by the inhibitor of gene expression 5,6-dichloro-1- β-
