KATTY ELENA BARRIOS ROJAS
BEHAVIOURAL DISCRIMINATION OF SOLITARY AND GREGARIOUS PHASES OF Anticarsia gemmatalis HÜBNER (LEPIDOPTERA:
NOCTUIDAE)
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Entomologia, para obtenção do título de Magister Scientiae.
VIÇOSA
ii
AGRADECIMENTOS
Ao Deus, por me dar saúde, energia e força todos os dias para realizar outro dos meus sonhos.
A Universidade Federal de Viçosa e ao Programa de Pós-Graduação em Entomologia pelos ensinamentos, os desafios e as experiências.
Á Organização de Estados Americanos (OEA) e a CAPES, pela concessão da bolsa de estudos no Brasil e por oferecer para mim uma oportunidade única na vida.
Ao meu orientador, o Professor Simon Luke Elliot pela confiança, por me dar a oportunidade de trabalhar no laboratório. Obrigada pelos ensinamentos, conselhos e ajuda durante o curso. Para mim foi uma experiência de aprendizagem em todos os sentidos. Serei sempre grata.
Ao meu coorientador, Farley William Souza pelos ensinamentos, confiança, paciência e pela ajuda aportada durante a realização do projeto.
A minha família por ser meu ponto de apoio em todo momento. Especialmente minha mãe por me apoiar, sem ela eu nunca teria conseguido. A minha irmã, obrigada por me manter focada e feliz, por me apoiar, por me dar razões a cada dia para lutar e conseguir tudo que nós merecemos. Amo vocês mais do que chocolate.
Ao Diego, meu amor. Obrigada por todos os momentos lindos vividos, pelas horas de risada, pelo apoio incondicional, pela companhia no laboratório, pela ajuda durante os experimentos, pelas aulas de português de graça, por tudo, te amo.
iii
Aos meus companheiros do laboratório de interações inseto-microrganismos, muito obrigada pela ajuda, pelas dicas e conselhos. Em especial agradeço a Verônica, Débora, Fernanda e Daniel pela ajuda nos experimentos.
A todas as pessoas que ajudaram na realização do projeto, muito obrigada.
iv SUMARIO
ABSTRACT ... vi
RESUMO ... viii
1. GENERAL INTRODUCTION ... 1
1.1 Phenotypic plasticity ... 1
1.2 Phase polyphenism-inducing factors ... 2
1.3 Phase change, behavioural characteristics ... 4
1.4 Adaptive significance of the phase change ... 5
2. REFERENCES ... 7
BEHAVIOURAL DISCRIMINATION OF SOLITARY AND GREGARIOUS PHASES OF Anticarsia gemmatalis HÜBNER (LEPIDOPTERA: NOCTUIDAE) 13 3. INTRODUCTION ... 14
4. METHODOLOGY ... 17
4.1 Anticarsia gemmatalis rearing conditions ... 17
4.2 Behavioural assays ... 18
4.2.1 Bioassay 1: Behavioural characterization of solitary and gregarious phases of Anticarsia gemmatalis. ... 18
4.2.2 Bioassay 2: Colour characterization of the head capsule and cuticle of different phases of Anticarsia gemmatalis. ... 20
4.2.3 Bioassay 3: Behavioural responses of Anticarsia gemmatalis to chemical stimulation. ... 21
4.2.4 Bioassay 4: Behavioural responses of Anticarsia gemmatalis to the mechanical stimulation... 22
4.3 Statistical analysis ... 22
4.3.1 Selection of behavioural discriminators of the solitary and gregarious phases of Anticarsia gemmatalis. ... 23
4.3.2 Selection and model fit ... 23
4.3.3 Probability of being classified as solitary ... 24
4.3.4 Colour characterization of the solitary and gregarious phases of Anticarsia gemmatalis. ... 24
v
5.1 Behavioural characterization of solitary and gregarious phases of Anticarsia
gemmatalis. ... 25
5.2 Colours characterization of solitary and gregarious phases of Anticarsia gemmatalis. ... 27
5.3 Phase change induction by mechanical and chemical stimulus ... 27
6. DISCUSSION ... 30
vi ABSTRACT
ROJAS, Katty Elena Barrios, M.Sc., Universidade Federal de Viçosa, February, 2015. Behavioural discrimination of solitary and gregarious phases of Anticarsia
gemmatalis Hübner (Lepidoptera: Noctuidae). Adviser: Simon Luke Elliot.
Co-adviser: Farley William Souza Silva.
Anticarsia gemmatalis is a species of Lepidoptera characterized by the capacity to
express differentially its genotype as part of adaptation strategies to environmental changes. This phenomenon is called phenotypic plasticity and is considered one of the main reasons for the evolutionary success. Phenotypes or "phases" described for larvae of Anticarsia gemmatalis are: solitary, gregarious and intermediary. These phases differ not only in morphological parameters but also in physiological, ecological and behavioural parameters. As regards the behaviour, many studies have confirmed that this is a basic component of phenotypic plasticity and the first to be expressed when there is a phase change. For Lepidoptera, although some clues are known, the factors that induce and maintain the phase change process have not been fully elucidated. Thus, the objectives of our study were to describe the behavioural repertoires of solitary and gregarious phases of Anticarsia gemmatalis and test if specific mechanical and/or chemical stimuli induce the phase change in this species. For the description of the behavioural repertoire were used 50 solitary and 50 gregarious phase larvae. In an experimental arena, each larva was observed for 780s in order to register the different components of its behaviour. For the induction of phase change, solitary phase larvae were selected and divided into groups in which stimuli were applied. To apply the mechanical stimulus 120 solitary larvae were selected and divided into 4 groups of 30 which were stimulated with a nylon thread on the head, on the third abdominal segment or on the last abdominal segment, according to the group. The control group remained without stimulation. To apply chemical stimulus 160 larvae were selected and divided into four groups of 40. For the three stimulus groups cuticular hydrocarbon extracts of Anticarsia gemmatalis solitary phase larvae, Anticarsia gemmatalis gregarious phase larvae and Spodoptera frugiperda larvae were used, according to the group. For the control group we
vii
viii RESUMO
ROJAS, Katty Elena Barrios, M.Sc., Universidade Federal de Viçosa, fevereiro de 2015.Discriminação comportamental das fases solitária e gregária de Anticarsia
gemmatalis Hübner (Lepidoptera: Noctuidae). Orientador: Simon Luke Elliot.
Coorientador: Farley William Souza Silva.
Anticarsia gemmatalis é uma espécie da ordem Lepidoptera caracterizada pela
capacidade de expressão diferencial de seu genótipo como parte de estratégias de adaptação às mudanças ambientais. O fenômeno é denominado plasticidade fenotípica e é considerado uma das principais razões do seu sucesso evolutivo. Os fenótipos ou “fases” descritas para as larvas de Anticarsia gemmatalis são: solitária, gregária e intermediária. Estas fases se diferenciam não só em parâmetros morfológicos, mas também em fisiológicos, ecológicos e comportamentais. No que respeita ao comportamento, diversos estudos confirmam que este é um componente básico da plasticidade fenotípica e uns dos primeiros em expressar-se quando ocorre uma mudança de fase. Para lepidópteros, embora se conheçam algumas pistas, os fatores que induzem e mantem o processo de mudança de fase não foram completamente elucidados. Diante disso os objetivos do nosso trabalho foram: descrever os repertórios comportamentais das fases solitária e gregária de Anticarsia gemmatalis e, testar se determinados estímulos mecânicos ou/e químicos induzem a
mudança de fase nesta espécie. Para a descrição do repertório comportamental foram utilizadas 50 larvas em fase solitária e 50 em fase gregária. Em uma arena experimental cada uma das larvas foi observada durante 780s, registrando assim diferentes componentes do comportamento. Para a indução da mudança de fase foram selecionadas larvas em fase solitária e divididas em grupos nos quais foram aplicados estímulos. Para aplicar o estimulo mecânico foram selecionadas 120 larvas solitárias e divididas em quatro grupos de 30. Foram feitas uma serie de toques com nylon na cabeça, no terceiro segmento abdominal ou no último segmento abdominal, segundo o grupo. O grupo controle permaneceu sem estimulo. Para aplicar o estimulo químico foram selecionadas 160 larvas e divididas em quatro grupos de 40. Foram usados extratos de hidrocarbonetos cuticulares de: larvas em fase solitária de Anticarsia gemmatalis, larvas em fase gregária de Anticarsia gemmatalis e larvas de
ix
1 1. GENERAL INTRODUCTION
1.1 Phenotypic plasticity
Environmental changes generate strong selective pressures in populations (Komers, 1997; Fusco & Minelli, 2010). Features that allow individuals to generate an adaptive response to these changes and to contribute to the fitness of future generations are favoured by natural selection (Pigliucci, 2005; Kowalski et al., 2014). One of these characteristics is phenotypic plasticity and has its genetic basis in the capacity of one genotype to produce more than one phenotype when exposed to different environments (Miner et al., 2005; Fusco & Minelli, 2010; Pfennig et al., 2010; Dingemanse & Wolf, 2013; Simões et al., 2013).
2
different stages of the life cycle in holometabolous insects; the presence or absence of winged females aphids; the castes of social insects and, the different phases in the development of some species of Lepidoptera and Orthoptera (Canfield et al., 2008; Suzuki & Nijhout, 2008; Fusco & Minelli, 2010; Beldade et al., 2011; Simpson et al., 2011). This last type of polyphenism is known “phase polyphenism” and was the
focus of our research.
1.2Phase polyphenism-inducing factors
The mechanism used by the individual to perceived and translated environmental stimuli, as well as the nature of these stimuli have been the focus of research and discussion for a long time (Kaiser et al., 2014). Within the phenomenon of phase polyphenism, some induction mechanisms are known (Badyaev, 2005; Whitman & Agrawal, 2009). Generally, hormonal processes are pointed out in the literature as induction mechanisms in several species (Nijhout, 1999; Boerjan et al., 2011). Hormones such ecdysteroids and juvenile hormone (JH) contribute to the phase change in Schistocerca gregaria (Hägele et al., 2004) and Anticarsia gemmatalis (Fescemyer & Hammond, 1988) respectively.
Other factors such as the availability of resources and population density have been suggested as factors (Gunn, 1998; Bailey et al., 2005). Population density has been considered as a key factor for the induction of several changes in the phenotype, genotype and behaviour of animals (Roessingh et al., 1998; Leo Lester et al., 2005; Cotter et al., 2008; Ma et al., 2011).
3
ever since this has been one of the most studied examples of phase polyphenism (Sword & Simpson, 2000; Simpson et al., 2011). Schistocerca gregaria belongs to a group of grasshoppers of the family Acrididae (Roessingh et al., 1993). One of the most distinctive characteristics of this group is the capacity to express two different phases through its development in response to changes in population density (Simpson et al., 1999). Uvarov defined these phases that vary in different features including morphology, physiology, ecology and behaviour (Simpson et al., 1999; Silva et al., 2013). Uvarov ascribed the solitary phase to that with green nymphs with relatively sedentary behaviour and a tendency to avoid conspecifics (Simpson et al., 1999). He ascribed the gregarious phase to that conformed by black nymphs with yellow or orange patches which have more active behaviour and a tendency to join with conspecifics (Bailey et al., 2005; Boerjan et al., 2011;.Simpson et al., 2011). Uvarov also noted that it is possible to find a large number of intermediate phenotypes between (Pener & Yerushalmi, 1998).
Some work with Schistocerca gregaria has demonstrated that density is in fact the main stimulus that induces the change (Simpson et al., 1999; Rogers et al., 2014). However, this process is not only about the number of individuals in a territory. An increase in density generates intraspecific competition for food, predation pressures and allows stimulus such as volatile and contact pheromones, visual and mechanical contact to contribute to this process (Sword et al., 2010). Of these stimuli, mechanical contact has emerged as one of the most important (Hägele & Simpson, 2000).
4
(Noctuidae). In aphids, a higher density of individuals on a plant usually facilitates or increases inter- and intraspecific interactions and generates responses such as the expression of a winged phenotype in females (Prokopy & Roitberg, 2001; Castañeda et al., 2010; Simpson et al., 2011).
In addition, species of Noctuidae (Lepidoptera) such as Anticarsia gemmatalis, have density-dependent phase polyphenism that is characterized by two
well-differentiated phases: solitary and gregarious (Silva et al., 2013). When the larvae are reared in isolation, their tendency is to be light green and when are reared together their tendency is to be black (Fescemyer & Erlandson, 1993). There is also a wide spectrum of intermediary phenotypes in the population and some of the phase features have been considered analogous to locusts (Silva et al., 2013). For Anticarsia gemmatalis, different factors such as plant age, humidity, temperature,
have been mentioned as inducers of morphological and physiological changes (Fescemyer & Hammond, 1986; Fescemyer & Erlandson, 1993). Recently, Silva et al (2013) suggested that another factor that could be critical in the phase change
induction is mechanical contact; however, this has not been studied in detail.
Finally, is also important to consider that besides these stimuli, the process can be also influenced by past experiences of parental generation, genotype variations and features of development (body size, reproductive patterns) (Dingemanse & Wolf, 2013).
1.3 Phase change, behavioural characteristics
5
et al., 1999). When behavioural characteristics are expressed, usually the species
show two or more types of behaviour. The capacity that allows individuals within a population to adjust their behavioural repertoire in response to changes in environmental conditions has been defined as “behavioural plasticity” (Geva et al., 2010; Kaiser et al., 2014).
There are two types of behavioural plasticity: developmental and activational. The first one refers to the adaptive change when a genotype expresses different behavioural phenotypes in different environments because of different developmental pathways induced by those environments. This process includes changes in morphology and physiology. The second refers to learning process that involves changes in the nervous system because of experience. This process includes structural and functional changes in neural networks (Snell-Rood, 2013).
For our work, we focus the attention on the behavioural plasticity related to development. Simpson et al (1999) pointed out that the study of behaviour represents improved tool to study phenotypic plasticity because despite other more conspicuous phase change characteristics such as colour, behavioural changes are the first to be expressed during the process and they are not dependent upon moulting, so can be studied over the life of the insect.
1.4 Adaptive significance of the phase change
6
diseases transmitted by contact. These factors could be considered adverse for the population fitness eventually (Cotter et al., 2004).
Depending on the situation, there are some hypotheses to explain the adaptive value of the phenotype change in high-density conditions (Cotter et al., 2004). For example, for morphological changes such colour, Simpson et al (1999) suggest that in locusts the colour change can serve as a visual cue to intraspecific processes such as aggregation of nymphs. Another alternative explanation for colour change is density-dependent prophylaxis (DDP) (Stearns, 1989; Barnes & Siva-Jothy, 2000; Sword, 2002; Cotter et al., 2004). In this hypothesis, the authors associated high levels of melanisation with high levels of the enzyme phenoloxidase enzyme and disease resistance (Elliot et al., 2003; Silva et al., 2013). They proposed that individuals belonging to the gregarious phase invest more in immune defence than those of the solitary phase and so combat the increased risk of infection at high densities (Elliot & Hart, 2010; Silva et al., 2013).
7 2. REFERENCES
Applebaum, S. W., & Heifetz, Y. (1999). Density-dependent physiological phase in insects. Annual Review of Entomology, 44, 317–341.
Badyaev, A. V. (2005). Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation. Proceedings of the Royal Society B: Biological Sciences, 272 (1566), 877–886.
Bailey, N., Gwynne, D., & Ritchie, M. (2005). Are solitary and gregarious mormon crickets (Anabrus simplex, Orthoptera, Tettigoniidae) genetically distinct?. Heredity, 95 (2), 166–73.
Barnes, A. I., & Siva-Jothy, M. T. (2000). Density-dependent prophylaxis in the mealworm beetle Tenebrio molitor L. (Coleoptera: Tenebrionidae): cuticular melanization is an indicator of investment in immunity. Proceedings of the Royal Society B: Biological Sciences, 267 (1439), 177–182.
Beldade, P., Mateus, A. R., & Keller, R. A. (2011). Evolution and molecular mechanisms of adaptive developmental plasticity. Molecular Ecology, 20 (7), 1347– 1363.
Boerjan, B., Sas, F., Ernst, U. R., Tobback, J., Lemière, F., Vandegehuchte, M. B., Janssen, C. R., Badisco, L., Marchal, E., Verlinden, H., & De Loof, A. (2011). Locust phase polyphenism : Does epigenetic precede endocrine regulation?.General and Comparative Endocrinology, 173 (1), 120–128.
Canfield, M. R., Greene, E., Moreau, C. S., Chen, N., & Pierce, N. E. (2008). Exploring phenotypic plasticity and biogeography in emerald moths : A phylogeny of the genus Nemoria (Lepidoptera : Geometridae). Molecular Phylogenetics and Evolution, 49 (2), 477–487.
8
Cotter, S. C., Hails, R. S., Cory, J. S., & Wilson, K. (2004). Density-dependent prophylaxis and condition-dependent immune function in lepidopteran larvae: a multivariate approach. Journal of Animal Ecology, 73 (2), 283–293.
Cotter, S. C., Myatt, J. P., Benskin, C. M. H., & Wilson, K. (2008). Selection for cuticular melanism reveals immune function and life-history trade-offs in Spodoptera littoralis. Journal of Evolutionary Biology, 21 (6), 1744–1754.
De Jong, G., & Bijma, P. (2002). Selection and phenotypic plasticity in evolutionary biology and animal breeding. Livestock Production Science, 78 (3), 195–214.
Dingemanse, N. J., & Wolf, M. (2013). Between-individual differences in behavioural plasticity within populations: causes and consequences. Animal Behaviour, 85 (5), 1031– 1039.
Elliot, S. L., & Hart, A. G. (2010). Density-dependent prophylactic immunity reconsidered in the light of host group living and social behavior. Ecology, 91 (1), 65–72.
Elliot, S. L., Blanford, S., Horton, C. M., & Thomas, M. B. (2003). Fever and phenotype: Transgenerational effect of disease on desert locust phase state. Ecology Letters, 6 (9), 830–836.
Fescemyer, H. W., & Erlandson, C. M. (1993). Influence of Diet on the Density-Dependent Phase Polymorphism of Velvetbean Caterpillars (Lepidoptera: Noctuidae). Physiological and Chemical Ecology, 22 (5), 933–941
Fescemyer, H. W., & Hammond, A. M. (1988). The relationship between population density, juvenile hormone, juvenile hormone esterase and phase variation in larvae of migrant insect, Anticarsia gemmatalis Hübner. Journal of Insect Physiology, 34 (1), 29–35.
9
Fusco, G., & Minelli, A. (2010). Phenotypic plasticity in development and evolution: facts and concepts. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 365 (1540), 547–556.
Geva, N., Guershon, M., Orlova, M., & Ayali, A. (2010). Memoirs of a locust : Density-dependent behavioral change as a model for learning and memory. Neurobiology of Learning and Memory, 93 (2), 175–182.
Gunn, A. (1998). The determination of larval phase coloration in the African armyworm, Spodoptera exempta and its consequences for thermoregulation and protection from UV light. Entomologia Experimentalis et Applicata, 86 (2), 125– 133.
Hägele, B. F., Wang, F.H., Sehnal, F., & Simpson, S. J. (2004). Effects of crowding, isolation, and transfer from isolation to crowding on total ecdysteroid content of eggs in Schistocerca gregaria. Journal of Insect Physiology, 50 (7), 621–628.
Hägele, B. F., & Simpson, S. J. (2000). The influence of mechanical, visual and contact chemical stimulation on the behavioural phase state of solitarious desert locusts (Schistocerca gregaria). Journal of Insect Physiology, 46 (9), 1295–1301. Hochkirch, A., Deppermann, J., & Gröning, J. (2008). Phenotypic plasticity in insects: the effects of substrate color on the coloration of two ground-hopper species. Evolution & Development, 10 (3), 350–359.
Kaiser, S. A., Sillett, T. S., & Webster, M. S. (2014). Phenotypic plasticity in hormonal and behavioural responses to changes in resource conditions in a migratory songbird. Animal Behaviour, 96, 19–29.
Komers, P. E. (1997). Behavioural plasticity in variable environments. Canadian Journal of Zoology, 75 (2), 161–169.
10
Leo Lester, R., Grach, C., Paul Pener, M., & Simpson, S. J. (2005). Stimuli inducing gregarious colouration and behaviour in nymphs of Schistocerca gregaria. Journal of Insect Physiology, 51 (7), 737–747.
Ma, Z., Guo, W., Guo, X., Wang, X., & Kang, L. (2011). Modulation of behavioral phase changes of the migratory locust by the catecholamine metabolic pathway. Proceedings of the National Academy of Sciences of the United States of America,
108 (10), 3882–3887.
Miner, B. G., Sultan, S. E., Morgan, S. G., Padilla, D. K., & Relyea, R. A. (2005). Ecological consequences of phenotypic plasticity. Trends in Ecology & Evolution, 20 (12), 685–692.
Nijhout, H. F. (1999). Control Mechanisms of Polyphenic Development in Insects. BioScience, 49 (3), 181–192.
Nijhout, H. F. (2003). Development and evolution of adaptive polyphenisms. Evolution & Development, 5(1), 9–18.
Pener, M. P. (1991). Locust Phase Polymorphism and its Endocrine Relations. Advances in Insect Physiology, 23, 1–79.
Pener, M.P., Yerushalmi, Y. (1998). The physiology of locust phase polymorphism: an update. Journal in Insect Physiology, 44 (5-6), 365-377.
Pfennig, D. W., Wund, M. A., Snell-Rood, E. C., Cruickshank, T., Schlichting, C. D., & Moczek, A. P. (2010). Phenotypic plasticity’s impacts on diversification and speciation. Trends in Ecology & Evolution, 25 (8), 459–467.
Pigliucci, M. (2005). Evolution of phenotypic plasticity: where are we going now ? Trends in Ecology & Evolution, 20 (9), 481–486.
11
Roessingh, P., Bouaïchi, A, & Simpson, S. J. (1998). Effects of sensory stimuli on the behavioural phase state of the desert locust, Schistocerca gregaria. Journal of Insect Physiology, 44 (10), 883–893.
Roessingh, P., Simpson, S. J., & James, S. (1993). Analysis of phase-related changes in behaviour of desert locust nymphs. Proceedings of the Royal Society B: Biological Sciences, 252 (1333), 43–49.
Rogers, S. M., Cullen, D. A., Anstey, M. L., Burrows, M., Despland, E., Dodgson, T., Matheson, T., Ott, S. R., Stettin, K., Sword, G. A., & Simpson, S. J. (2014). Rapid behavioural gregarization in the desert locust, Schistocerca gregaria entails synchronous changes in both activity and attraction to conspecifics. Journal of Insect Physiology, 65, 9–26.
Snell-Rood, E. C. (2013). An overview of the evolutionary causes and consequences of behavioural plasticity. Animal Behaviour, 85 (5), 1004–1011.
Silva, F.W, Viol, D. L., Faria, S. V., Lima, E., Valicente, F. H., & Elliot, S. L. (2013). Two’s a Crowd: phenotypic adjustments and prophylaxis in Anticarsia gemmatalis larvae are triggered by the presence of conspecifics. PLoS ONE, 8 (4),
1–10.
Simpson, S. J., McCaffery, A. R., & Hägele, B. F. (1999). A behavioural analysis of phase change in the desert locust. Biological Reviews, 74 (4), 461–480.
Simpson, S. J., Sword, G. A., & Lo, N. (2011). Polyphenism in insects. Current Biology, 21 (18), R738–R749.
Simões, P. M. V., Niven, J. E., & Ott, S. R. (2013). Phenotypic transformation affects associative learning in the desert locust. Current Biology, 23 (23), 2407– 2412.
12
Sword, G. A., & Simpson, S. J. (2000). Is there an intraspecific role for density-dependent colour change in the desert locust?. Animal Behaviour, 59 (4), 861–870. Sword, G. A., Lecoq, M., & Simpson, S. J. (2010). Phase polyphenism and preventative locust management. Journal of insect physiology, 56 (8), 949-957. Stearns, S. C. (1989). The evolutionary significance of phenotypic plasticity. BioScience, 39 (7), 436–445.
Suzuki, Y., & Nijhout, H. F. (2008). Genetic basis of adaptive evolution of a polyphenism by genetic accommodation. Journal of Evolutionary Biology, 21 (1), 57–66.
Whitman, D. W., & Agrawal, A. A. (2009). What is phenotypic plasticity and why is it important?. In Phenotypic Plasticity of Insects (pp. 1–63). Science publisher.
13
BEHAVIOURAL DISCRIMINATION OF SOLITARY AND
GREGARIOUS PHASES OF Anticarsia gemmatalis HÜBNER
(LEPIDOPTERA: NOCTUIDAE)
Katty Elena Barrios Rojas, Farley William Souza Silva, Simon Luke Elliot
_________________________________________________________
Departamento de Entomologia, Universidade Federal de Viçosa, Viçosa-MG, Brasil.
14 3. INTRODUCTION
For many organisms the capacity to deal with environment changes has an influence in their survival and reproductive success (Via et al., 1995; Komers, 1997; Foster, 2013). Many species have the capacity to express their genotype in a differential way and to show different phenotypes according to their interaction with the environment (Nijhout, 1999; Simpson et al., 2011). These species stand out for their great capacity to adapt to different selective pressures (Nijhout, 2003). The capacity is called "phenotypic plasticity” and is closely related to the evolutionary success of these species. It allows organisms to deal with a variety of environmental pressures, to occupy different ecological niches, to extend their geographical distribution, and to decrease the probability of extinction (Pfennig et al., 2010; Snell-Rood et al., 2010; Simpson et al., 2011).
One type of phenotypic plasticity is called “Phase polyphenism” and is a
widespread phenomenon in insects occurring in species of Lepidoptera, Orthoptera, Coleoptera and Hemiptera (Simpson et al., 2011; Silva et al., 2013). The phenomenon is induced by several stimuli (Komers, 1997). The increase in population density is one of the most mentioned in the literature as phase change-induction stimuli. The change-induction in this case may correlate with the increased likelihood of inter- or intraspecific interactions (Anazonwu & Johnson, 1986; Silva et al., 2013; Vilcinskas, 2013). Examples of insects that have different density
dependent phases are Spodoptera littoralis, Spodoptera exempta, Anticarsia gemmatalis (Lepidoptera: Noctuidae) and Schistocerca gregaria (Orthoptera:
15
The larvae of Anticarsia gemmatalis are characterized by expressing three principal phases in response to changes in the environmental conditions: solitary, gregarious and intermediary (Fescemyer & Hammond, 1988; Silva et al. 2013). The solitary phase is composed of individuals with cuticle and cephalic capsule light green and without any black patterns. Individuals that have black cuticle and yellowish cephalic capsule compose the gregarious phase. The intermediary phase has a spectrum of colour combinations (Fescemyer & Erlandson, 1993; Silva et al., 2013).
The phase change involves a complex set of characters including morphology, physiology and behaviour (Hägele & Simpson, 2000; Silva et al., 2013; Simpson et al., 1999). Within these characteristic one of the principal to be studied is behaviour because it is the first feature to be expressed in the process of phase change and is continuous throughout the life of an insect (Komers, 1997; Simpson et al., 1999; Sword, 2003). In species of locusts for example, one of the earliest
distinguishing features of phase change from solitary to gregarious is the interaction with conspecifics (Simpson et al., 1999; Simpson et al., 2001). When the locusts nymphs are in solitary phase they exhibit a clear repellence by conspecifics but after phase change is induced they are attracted to conspecifics (Sword & Simpson, 2000).
16
al.,2005; Praça et al., 2006). Despite this neither the induction mechanisms nor the
behavioural repertories of each phase have been described.
For these reasons, the objectives of our research were: (1) to determine the behavioural repertoires of the two extreme phases of Anticarsia gemmatalis: solitary and gregarious, and (2) to evaluate how mechanical and chemical stimuli applied to solitary phase larvae might induce changes in behavioural or morphological characteristics (cuticle and cephalic capsule larval colour).
17 4. METHODOLOGY
4.1 Anticarsia gemmatalis rearing conditions
In the laboratory, the Anticarsia gemmatalis population was establish using fist instar larvae. First, we put each larva in a 50 ml disposable plastic cup with artificial diet ad libitum. The diet was made according to Hoffman-Campo et al. (1985). Plastic cups were used to reduce the risk of contamination and to reduce labour in the laboratory (Hoffman-Campo et al., 1985).
The cups containing the larvae were kept in an incubator under controlled conditions (25 ± 5 °C and 12:12 light-dark photoperiod) until the final instar. The pupae were placed in glass cages and kept in a rearing room (25 ± 5 °C and 70 ± 5% RH with a 12:12 light-dark photoperiod), until adult emergence. Once emerged, moths remained in the glass cage and were fed daily with a water, beer, honey and sucrose solution (Hoffman-Campos et al., 1985). White paper sheets were placed inside the cage to serve as substrate for the oviposition of the moths. Paper sheets were exchanged approximately every two days with a new one. The paper sheets with eggs were separated, placed in a 500 ml plastic container and, maintained with artificial diet under controlled conditions until hatching. The whole process was repeated in order to maintain the laboratory population.
18 4.2 Behavioural assays
4.2.1 Bioassay 1: Behavioural characterization of solitary and gregarious phases of Anticarsia gemmatalis.
To accomplish the first objective we set up an experimental arena using a white plastic tray with the following dimensions: 32 cm x 24 cm x 7 cm. In the right and left sides of the tray, we made a hole in which were placed the stimulus and no-stimulus test tubes. The no-stimulus tube contained 25 gregarious phase larvae and the no-stimulus tube was empty (adapted from Simpson et al., 1999). The position of these tubes was changed between test individuals. The tray walls were covered with tape and with red cellophane paper, we made a cover to prevent light stimulus and at the same time facilitate observation.
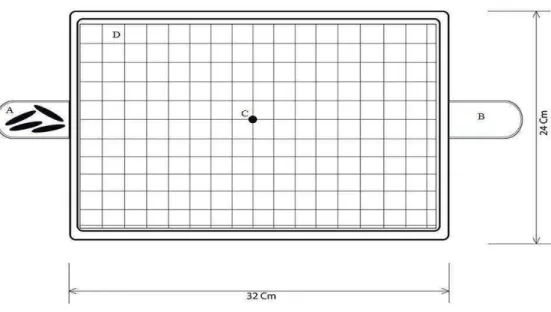
For the test, we selected a single larva and then we placed it in the centre of the experimental arena. We kept it under observation for 780 seconds and recorded by hand all the behaviours shown by the individual. In this fashion, 100 larvae were evaluated (50 of each phase). Figure 1 shows the experimental arena.
19
Figure 1. Basic schema of the experimental arena (top view). In the scheme the letters A, B, C and D. represent the principal points of the experimental arena. A) Tube with the stimulus group B) Tube without the stimulus group C) Point of larvae positioning in the beginning of each bioassay, and D) Graph paper located at the bottom of the arena to help with the record of the direction and total distance walked by the larvae. Were evaluated 100 larvae of Anticarsia gemmatalis, 50 reared alone and 50 reared in groups. The stimulus group was made up of 25 gregarious larvae.
20
Table 1. Description of the behavioural variables evaluated for Anticarsia gemmatalis larvae in the experimental arena.
Behavioural variables
Description
Stimulus position
Position of the stimulus in the sides of the experimental arena (right or left). Exchanged between each test.
Walking distance
Number of centimetres walked by larvae during the total time in the arena.
Number of walking
events
Number of times when the larva walked. A new event was considered when the larva did not walk for at least 5 seconds. Initial time at
rest
Time in seconds for which the larva remained motionless in the centre of the arena before its first walk event.
Number of head
movements
Number of times that the larva moved its head to one side or another.
Final Larva position
Position of the larva at the end of the bioassay. This variable was registered with values 0 or 1. The zero value indicated that the larva was in the opposite direction of the stimulus and the one value indicated that direction of the larva was in favour of the stimulus.
4.2.2 Bioassay 2: Colour characterization of the head capsule and cuticle of different phases of Anticarsia gemmatalis.
In order to have a control that help with the recognition and discrimination of the Anticarsia gemmatalis larvae used for the experiments, ninety larvae were selected, thirty from each phase: green, black and intermediary. These larvae were photographed one by one, with a Power Shot A640 camera attached to the Zeiss Axioskop 40 light microscope.
21
4.2.3 Bioassay 3: Behavioural responses of Anticarsia gemmatalis to chemical stimulation.
To evaluate the response of the Anticarsia gemmatalis larvae to chemical stimulation, 160 uniformly light green larvae were selected. Those larvae were divided into four groups of 40. Three groups were used for treatment and one for control. The three treatments were performed using cuticular hydrocarbons as stimulus. The cuticular hydrocarbons were extracted by submerging approximately 30 larvae in 50 ml of dichloromethane for 10 minutes (adapted from Hägele & Simpson, 2000). Using a graduated pipette the extract was placed on a filter paper that was at the bottom of a Petri dish and the larvae were placed to walk for 12 hours.
For treatment 1, the cuticular hydrocarbons were extracted from Anticarsia gemmatalis black phase larvae. For treatment 2 these were extracted from larvae of
Spodoptera frugiperda (reared under the same conditions as Anticarsia gemmatalis).
For treatment 3, these were extracted from Anticarsia gemmatalis green phase larvae. The control was done by just applying dichloromethane on filter paper. After 12 hours having installed the experiment, the larvae were evaluated in order to find indicators of a possible phase change. For this purpose we evaluated the behaviour and the final colour of the larvae after the completed bioassay.
22
4.2.4 Bioassay 4: Behavioural responses of Anticarsia gemmatalis to the mechanical stimulation
To evaluate the induction of phase change by a mechanical stimulus we selected 120 uniformly light green larvae. The larvae were divided into four groups, three with a stimulus and a fourth without stimulus (control). The groups with stimulus were denominated A, B and C. In group A, the stimulus was applied to the head of the larva. In group B this was applied in the third abdominal segment (at the height of the first pair of legs), and in group C this was applied in final segment of the abdomen (adapted from Simpson et al. 2001).
The stimulus consisted in 30 touches made with a nylon wire (simulating contact). The stimulus was done every half hour for 4 continuous hours (adapted from Simpson et al., 2001). After finishing the test, we evaluated the larvae in order to find indicators of a possible phase change. For this purpose we evaluated the behaviour and the final colour of the larvae. The behavioural repertoire of each larva was described using the same method and measuring the same variables as in the bioassay 4.2.1. After 24 hours, larvae were also photographed to measure the lightness of the cuticle and head capsule using the same method as in the bioassay 4.2.2. It was evaluated 24 hours later to try to register possible late effects on colour changes (Simpson et al., 2001).
23
4.3.1Selection of behavioural discriminators of the solitary and gregarious phases of Anticarsia gemmatalis.
In order to know if the solitary and gregarious phases of Anticarsia gemmatalis are different in their behaviours, the data collected in the experimental
arena was analysed. For this analysis we performed a logistic regression using the statistical program SPSS. The program regressed the response variable against a series of explanatory variables (behavioural parameters).The explanatory variables used were: stimuli position, walking distance, number of walking events, initial time at rest, number of head movements and final larva position. The response variable was the phase. This regression type is the most appropriate in this case because the response variable "phase" is binary or dichotomous. The logistic regression analyses are widely recommended for such behavioural studies (Roessingh et al., 1993; Rogers et al., 2014; Simpson et al., 1999). These analyses make no assumptions about the distribution of the data (Roessingh et al., 1993).
4.3.2 Selection and model fit
The first step in the logistic regression analysis was to create an initial model including all explanatory variables as a function of the response variable (Simpson et al., 1999). After obtain the p-values for each variable, the initial model was simplified to its most parsimonious using the ‘stepwise elimination’ procedure,
which is commonly recommended by statistical texts and manuals (Hardy & Field, 1998). That final version was called final model.
24
predicted values of the response variable. In this case, the best model adjustment is indicated by a smaller difference between the observed and predicted values. A good model fit is indicated by a chi-square value not significant (Hosmer et al., 1997).
4.3.3 Probability of being classified as solitary
After stabilizing the final model and identify the behavioural variables that help to discriminate between behaviours of the Anticarsia gemmatalis phases we used a logistic algorithm derived from the final model to calculate the probability of larvae resulting for the bioassays 3 and 4 to be classified as solitary after chemical and mechanical stimulus (Hardy & Field, 1998). We compared the effect of the stimulus evaluating changes in larvae behaviour and colour. For this we performed an analysis of variance (ANOVA) using statistical software R. Throughout this study we used the term “Psolitary” to indicate the probability of being classified as solitary larvae.
4.3.4 Colour characterization of the solitary and gregarious phases of Anticarsia gemmatalis.
25 5. RESULTS
5.1 Behavioural characterization of solitary and gregarious phases of Anticarsia gemmatalis.
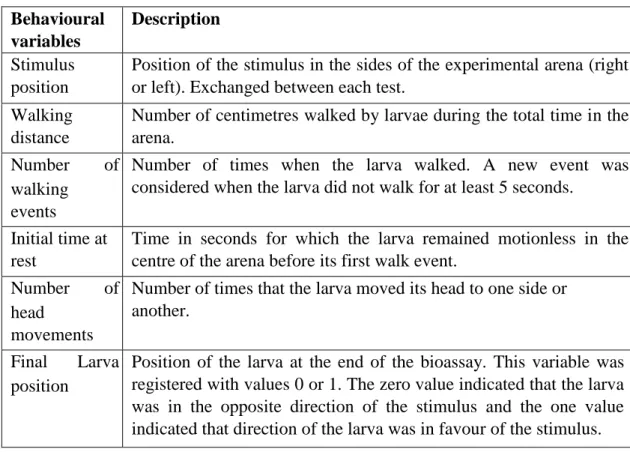
The test in the arena allowed discriminated the behaviour characteristics of the solitary and gregarious phases of Anticarsia gemmatalis. In the table number 2, we present the results obtained in the initial model.
Table 2. Behavioural variables retained in the logistic regression initial model from solitary and gregarious larvae of Anticarsia gemmatalis.
Behavioural variables S.E. Wald df p-value Exp(β)
Stimulus position 0,013 0,432 0,001 1 0,975 1,013
Walking distance 0,006 0,007 0,731 1 0,393 1,006
Number of walking events -0,075 0,028 7,008 1 0,008 0,928 Initial time at rest -0,013 0,008 2,817 1 0,093 0,987 Number of head movements 0,007 0,004 3,297 1 0,069 1,007 Final larval position -0,638 0,433 2,176 1 0,140 0,528
Constant 1,130 0,667 2,871 1 0,090 3,096
All these values are calculated for solitary phase Anticarsia gemmatalis larvae. The β value indicates
the relation between the explanatory and response variable, the sign (positive or negative) indicates the direction of the relationship. S.E values are the standard errors. Wald is a statistical that allows comparing the maximum likelihood estimate of the behavioural parameters and its estimate standard error, higher values than four indicate that the explanatory variable explains the variable response. df values are the degrees of freedom of the model. p-value is the measure of significance of variable in
the model. Exp(β) values indicates the strength of the relationship between variables, values more
distant of 1 indicate a stronger numerical relationship. For an explanation of the behavioural variables,
see table 1.
26
gregarious phase larvae. The Wald statistic value above 4 indicates that the explanatory variable is appropriate to describe the behaviour. Once after getting that result, they were removed one by one the variables in order to optimize the process. The results of the final model are in table 3.
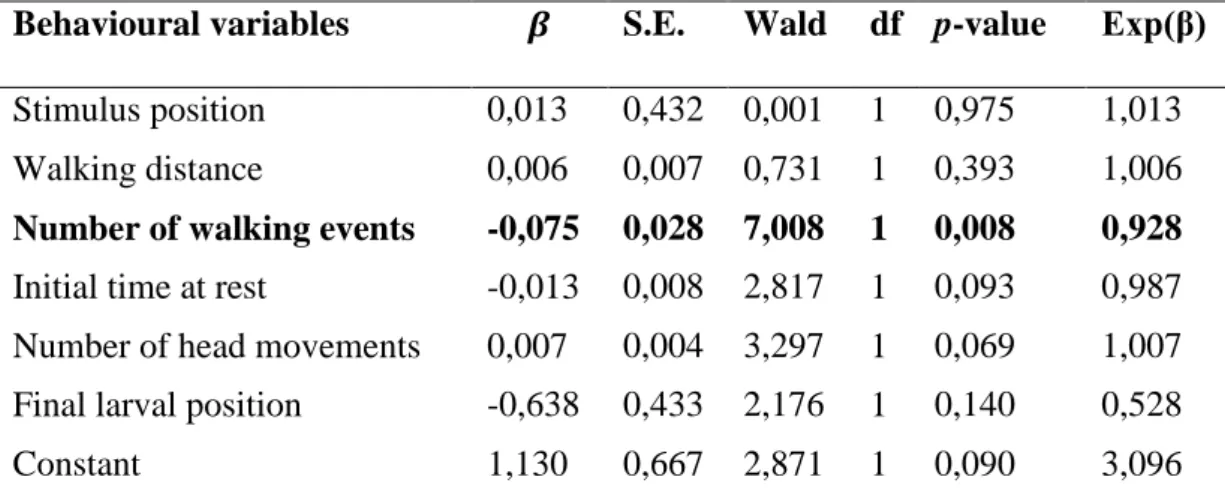
Table 3. Behavioural variables retained in the logistic regression final model from solitary and gregarious larvae of Anticarsia gemmatalis.
Behavioural variables S.E Wald df p-value Exp(
Number of walking events -0,061 0,023 6,968 1 0,008* 0,941
Number of head
movements
0,008 0,004 4,590 1 0,032* 1,008
Constant 0,316 0,480 0,433 1 0,511 1,371
All these values are calculated for solitary phase Anticarsia gemmatalis larvae. The β value indicates
the relation between the explanatory and response variable, the sign (positive or negative) indicates the direction of the relationship. S.E values are the standard errors. Wald is a statistical that allows comparing the maximum likelihood estimate of the behavioural parameters and its estimate standard error, higher values than four indicate that the explanatory variable explains the variable response. df values are the degrees of freedom of the model. p-value is the measure of significance of variable in
the model. Exp(β) values indicates the strength of the relationship between variables, values more
distant of 1 indicate a stronger numerical relationship. For an explanation of the behavioural variables see table 1.
Logistic regression final model enable to select two of the six variables included in the initial model. These two variables: number of walking events (p=0,008) and number of head movements (p=0,032) are considerate as predictive variables for the behaviour of the larvae (Table 3).The results showed that although the two phases share a similar behavioural repertoire, there have also differences. Solitary phase larvae have a lower number of walking events than the gregarious phase larvae. On the contrary, the solitary phase larvae differ from gregarious phase for the large number of head movements.
27
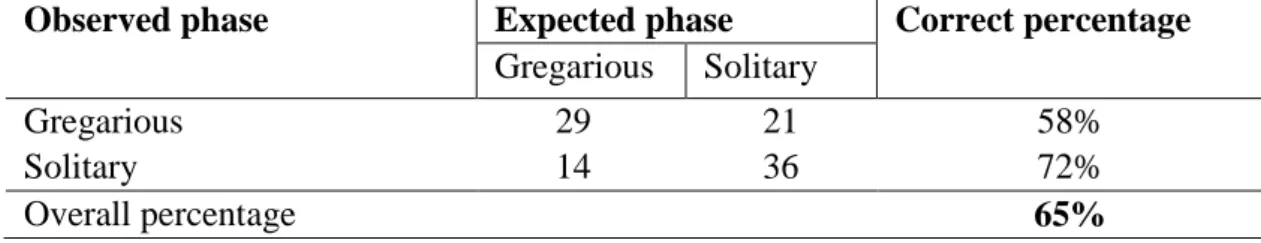
phase larvae (58%) and 36 as solitary phase larvae (72%) of the initial 100 larvae (table 4).
Table 4. Observed and expected phase predicted values to Anticarsia gemmatalis phases and its percentage of correct classification based on the logistic regression final model.
Observed phase Expected phase Correct percentage
Gregarious Solitary
Gregarious 29 21 58%
Solitary 14 36 72%
Overall percentage 65%
N=100 Anticarsia gemmatalis larvae (50 of each phase)
5.2 Colours characterization of solitary and gregarious phases of Anticarsia gemmatalis.
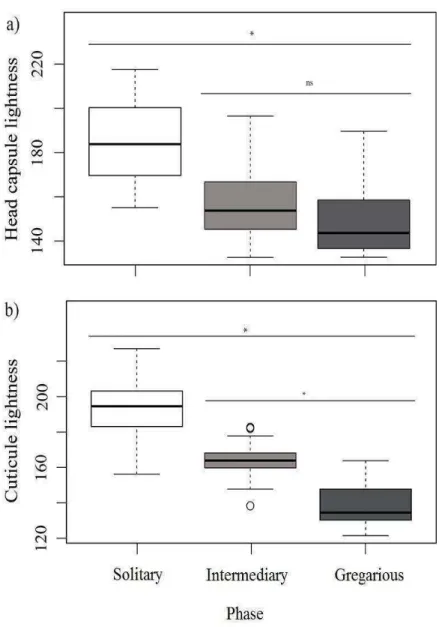
The results for a colour characterization of the Anticarsia gemmatalis phases are represent in the Figure 2. In this figure is possible to observe that indeed there is a significant difference between the lightness of the cuticle and the cephalic capsule from solitary, gregarious and intermediary phase larvae. These data reflects natural tendency of the species, so the solitary phase larvae are different both in cuticular and cephalic capsule lightness of the intermediary and gregarious phase larvae. The solitary phase larvae show lightness values closer to zero (or white) according with the established scale. In contrast, the gregarious phase larvae show lightness values closer to 255 (or black). The results were used as a control to have one objective morphological measure of each phase to compare them.
5.3 Phase change induction by mechanical and chemical stimulus
28
Figure 2. Distribution of lightness cuticle and the head capsule values calculated for three larval phases of Anticarsia gemmatalis: solitary, gregarious and intermediary. N = 30 for each phase. (*) significantly different (ns) nosignificantly different.
29
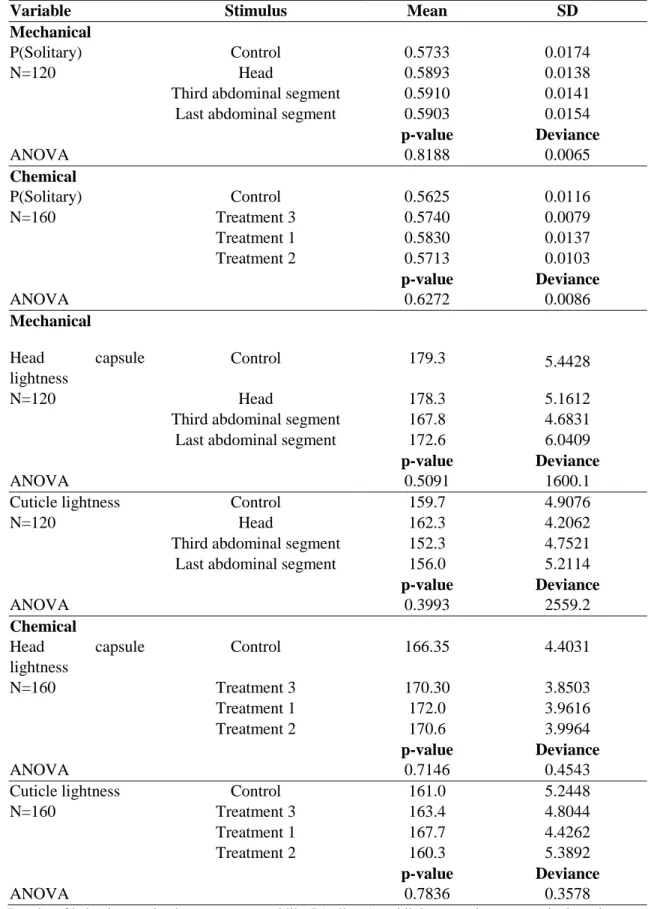
Table 5. Effect of mechanical and chemical stimulation on the induction
of Anticarsia gemmatalis phase change.
Variable Stimulus Mean SD
Mechanical
P(Solitary) Control 0.5733 0.0174
N=120 Head 0.5893 0.0138
Third abdominal segment 0.5910 0.0141
Last abdominal segment 0.5903 0.0154
p-value Deviance
ANOVA 0.8188 0.0065
Chemical
P(Solitary) Control 0.5625 0.0116
N=160 Treatment 3 0.5740 0.0079
Treatment 1 0.5830 0.0137
Treatment 2 0.5713 0.0103
p-value Deviance
ANOVA 0.6272 0.0086
Mechanical
Head capsule lightness
Control 179.3 5.4428
N=120 Head 178.3 5.1612
Third abdominal segment 167.8 4.6831
Last abdominal segment 172.6 6.0409
p-value Deviance
ANOVA 0.5091 1600.1
Cuticle lightness Control 159.7 4.9076
N=120 Head 162.3 4.2062
Third abdominal segment 152.3 4.7521
Last abdominal segment 156.0 5.2114
p-value Deviance
ANOVA 0.3993 2559.2
Chemical
Head capsule lightness
Control 166.35 4.4031
N=160 Treatment 3 170.30 3.8503
Treatment 1 172.0 3.9616
Treatment 2 170.6 3.9964
p-value Deviance
ANOVA 0.7146 0.4543
Cuticle lightness Control 161.0 5.2448
N=160 Treatment 3 163.4 4.8044
Treatment 1 167.7 4.4262
Treatment 2 160.3 5.3892
p-value Deviance
ANOVA 0.7836 0.3578
Results of behaviour and colour are presented like P(solitary) and lightness values respectively and are
30 6. DISCUSSION
During this investigation we evaluated both the behavioural characteristics of the larvae of solitary and gregarious phases of Anticarsia gemmatalis as well as the role some chemical and mechanical stimuli in phase change induction in this species. Our findings provided evidence that the solitary and gregarious phases of Anticarsia gemmatalis that our laboratory population can be separated based on their
behaviour in the bioassay arena.
The results indicated that the behavioural repertoires of both phases are composed of at least six conspicuous behavioural variables. Two of those variables can be used to discriminated the phases. One is the number of walking events and the other the number of head movements. Although these are common behaviours, the frequency of each was different in each phase. The solitary larvae have a greater number of head movements and the gregarious larvae a greater number of walking events. According to Ewing & Manning (1967), this may be due to differences in genotype expression.
31
On the other hand, gregarious phase individuals become more conspicuous to predators. The adaptive role of dark colour and active movement has been widely studied in locusts (Sword & Simpson, 2000). In this case, the colour could be associated with aposematism (Despland & Simpson, 2005b). Gregarious nymphs commonly feed on plants with toxins in contrast with solitary nymphs and are associated by predators with little or no palatability (Sword et al., 2000; Despland & Simpson, 2005a).
In the case of Anticarsia gemmatalis there is so far no evidence of an aposematic function, so the colour can be explained in other ways (Sword & Simpson, 2000). As pointed out by some authors the dark colour patterns may be functioning as visual cues for the localization of conspecifics (Sword & Simpson, 2000; Despland & Simpson, 2005b). As Spodoptera exempta, Anticarsia gemmatalis larvae have a tendency to express black coloration at low population densities (Silva et al., 2013) which might suggest that this is an important adaptive advantage for both species (Gunn, 1998).
32
With respect to head movements, our results show that Anticarsia gemmatalis solitary phase larvae performed a higher number than gregarious phase
larvae. Dethier (1942) pointed out that lepidopteran larvae are capable of detecting light using their sensory organs called stemmata. Both phases perform movements with their heads from one side to other. This behaviour is called searching in the literature and is done by the larvae in order to scanning their surrounding environment and orient themselves (Bell, 1990; Wicklein, 2003). Searching behaviour represents the confluence of three kinds of factors: biological characteristics and abilities to perceive sensory information of insects, environmental factors (availability of resources, risk of predation), internal factors, determining what an individual needs (Bell, 1990). Our results show differential performance of head movements between the phases which could suggest that there is a difference in the perception of stimuli between them (but have no evidence for this at the moment).
In general, these Anticarsia gemmatalis behavioural results are similar to the results found for Spodoptera larvae and Schistocerca gregaria. It has been reported that those larvae and nymphs are more active when they are reared in groups than when reared in isolation (Hodjat, 1970; Despland & Simpson, 2005b).
33
The second objective of or work was to assess whether certain mechanical and chemical stimuli contributed to the phase shift in this Anticarsia gemmatalis. Our results showed that neither chemical nor mechanical stimuli applied to solitary larvae had a primary role in the induction of phase change process and this may be due to several factors. For mechanical induction it is known that tactile stimuli is an important factor for phase changes in different species but their efficacy will depend on the stimulation site (Rogers et al., 2003).We applied the stimuli in the head, the third abdominal segment and in the last abdominal segment. These body regions were selected for methodological practicality, but the stimuli do not have an induction response. These results could be reflect that body areas as: head, legs, thorax and abdomen should not cause pronounced behavioural phase change, because these structures are regularly stimulated by the animal itself during normal feeding, and walking activity (Simpson et al., 2001). In addition, the intensity of the stimulus could be related, while some authors applied touch stimulus during five seconds every 60 seconds, we applied the touches every half hour.
34
About colour-change results, there is not a positive correlation between stimulus and the induction treatments. It has been reported the Anticarsia gemmatalis real phase did not appear until after it had moulted to the last instar (Fescemyer & Hammond, 1986). Because we measured the colour variations four hours later (mechanical stimulation) and 12 hours later (chemical stimulus) it could be that the time required for the result to be apparent and measurable was not enough in either case.
In general, other factors could be affecting our results. First, the rearing histories prior to capture of the Anticarsia gemmatalis larvae were unknown. There is a possibility that the variation in behaviour observed was due to uncontrolled differences in parental effects between populations (Sword, 2003). Secondly, it has been reported that larvae phase change occur in species that are actively gregarious, and although it has a tendency, Anticarsia gemmatalis is not considered a gregarious species (Anazonwu & Johnson, 1986). Third, the chance process could occur over many timescales some characters change rapidly while others change slowly through accumulation across generations (Simões et al., 2013), however we evaluated only one instar of one generation. Fourthly, many insects have a critical period in development when the individuals are sensitive to induction stimuli, so there is the possibility that the stimulation period chosen (3rd-4th instar) was not adequate for this species.
35
36 7. REFERENCES
Anazonwu, D. L., & Johnson, S. J. (1986). Effects of host and density on larval color, size and development of the velvethean caterpillar Anticarsia gemmatalis (Lepidoptera: Noctuidae). Enviromental Entomology, 15 (4), 779-783.
Applebaum, S. W., & Heifetz, Y. (1999). Density-dependent physiological phase in insects. Annual Review of Entomology, 44, 317–341.
Badyaev, A. V. (2005). Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation. Proceedings of the Royal Society B: Biological Sciences, 272 (1566), 877–886.
Bailey, N.W, Gwynne, D.T, & Ritchie, M. G. (2005). Are solitary and gregarious mormon crickets (Anabrus simplex, Orthoptera, Tettigoniidae) genetically distinct? Heredity, 95 (2), 166–73.
Barnes, A. I., & Siva-Jothy, M. T. (2000). Density-dependent prophylaxis in the mealworm beetle Tenebrio molitor L. (Coleoptera: Tenebrionidae): cuticular melanization is an indicator of investment in immunity. Proceedings of the Royal Society B: Biological Sciences, 267 (1439), 177–182.
Beldade, P., Mateus, A. R., & Keller, R. A. (2011). Evolution and molecular mechanisms of adaptive developmental plasticity. Molecular Ecology, 20 (7), 1347– 1363.
Bell, W. J (1990). Searching behavior patterns in insects. Annual Review of Entomology, 35, 447–467.
Boerjan, B., Sas, F., Ernst, U. R., Tobback, J., Lemière, F., Vandegehuchte, M. B., Janssen, C. R., Badisco, L., Marchal, E., Verlinden, H., Schoofs, L., & De Loof, A. (2011). Locust phase polyphenism : Does epigenetic precede endocrine regulation?. General and Comparative Endocrinology, 173(1), 120–128.
37
Hüebner, 1818 (Lepidoptera : Noctuidae) em soja, amendoim e dieta artificial. Boletin Sanidade Vegetal. Plagas, 31, 171–178.
Canfield, M. R., Greene, E., Moreau, C. S., Chen, N., & Pierce, N. E. (2008). Exploring phenotypic plasticity and biogeography in emerald moths : A phylogeny of the genus Nemoria (Lepidoptera : Geometridae). Molecular Phylogenetics and Evolution, 49 (2), 477–487.
Castañeda, L. E., Figueroa, C. C., Bacigalupe, L. D., & Nespolo, R. F. (2010). Effects of wing polyphenism, aphid genotype and host plant chemistry on energy metabolism of the grain aphid, Sitobion avenae. Journal of Insect Physiology, 56 (12), 1920–1924.
Chambers, P., Sword, G., Angel, J. E., Behmer, S., & Bernays, E. A. (1996). Foraging by generalist grasshoppers: two different strategies. Animal Behaviour, 52 (1), 155–165.
Cotter, S. C., Hails, R. S., Cory, J. S., & Wilson, K. (2004). Density-dependent prophylaxis and condition-dependent immune function in lepidopteran larvae: a multivariate approach. Journal of Animal Ecology, 73 (2), 283–293.
Cotter, S. C., Myatt, J. P., Benskin, C. M., & Wilson, K. (2008). Selection for cuticular melanism reveals immune function and life-history trade-offs in Spodoptera littoralis. Journal of Evolutionary Biology, 21 (6), 1744–1754.
De Jong, G., & Bijma, P. (2002). Selection and phenotypic plasticity in evolutionary biology and animal breeding. Livestock Production Science, 78 (3), 195–214.
Despland, E., & Simpson, S. J. (2005a). Food choices of solitarious and gregarious locusts reflect cryptic and aposematic antipredator strategies. Animal Behaviour, 69 (2), 471–479.
38
Dethier, V. G. (1942). The dioptric apparatus of lateral ocelli. Journal of Cellular and Comparative Physiology, 19 (3), 301–313.
Dingemanse, N. J., & Wolf, M. (2013). Between-individual differences in behavioural plasticity within populations: causes and consequences. Animal Behaviour, 85 (5), 1031– 1039.
Elliot, S. L., & Hart, A. G. (2010). Density-dependent prophylactic immunity reconsidered in the light of host group living and social behavior. Ecology, 91 (1), 65– 72.
Elliot, S. L., Blanford, S., Horton, C. M., & Thomas, M. B. (2003). Fever and phenotype: Transgenerational effect of disease on desert locust phase state. Ecology Letters, 6 (9), 830–836.
Ewing, A. W., & Manning, A. (1967). The evolution and genetics of insect behaviour. Annual Review of Entomology, 12, 471–494.
Fescemyer, H. W., & Erlandson, C. M. (1993). Influence of diet on the density-dependent phase polymorphism of velvetbean caterpillars (Lepidoptera: Noctuidae). Physiological and Chemical Ecology, 22 (5), 933-941.
Fescemyer, H. W., & Hammond, A. M. (1986). Effect of density and plant age on color phase variation and development of larval velvethean caterpillar, Anticarsia gemmatalis Hühner (Lepidoptera : Noctuidae). Enviromental Entomology, 15 (4),
784–789.
Fescemyer, H. W., & Hammond, A. M. (1988). The relationship between population density, juvenile hormone, juvenile hormone esterase and phase variation in larvae of migrant insect, Anticarsia gemmatalis Hübner. Journal of Insect Physiology, 34 (1), 29–35.
39
Fugi, C. G. Q., Lourenção, A. L., & Parra, J. R. P. (2005). Biology of Anticarsia gemmatalis on soybean genotypes with different degrees of resistance to insects.
Scientia Agricola, 62 (1), 31–35.
Fusco, G., & Minelli, A. (2010). Phenotypic plasticity in development and evolution: facts and concepts. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 365 (1540), 547–556.
Geva, N., Guershon, M., Orlova, M., & Ayali, A. (2010). Memoirs of a locust : Density-dependent behavioral change as a model for learning and memory. Neurobiology of Learning and Memory, 93 (2), 175–182.
Gunn, A. (1998). The determination of larval phase coloration in the African armyworm, Spodoptera exempta and its consequences for thermoregulation and protection from UV light. Entomologia Experimentalis et Applicata, 86 (2), 125– 133.
Hägele, B. F., & Simpson, S. J. (2000). The influence of mechanical, visual and contact chemical stimulation on the behavioural phase state of solitarious desert locusts (Schistocerca gregaria). Journal of Insect Physiology, 46 (9), 1295–1301. Hägele, B. F., Wang, F. H., Sehnal, F., & Simpson, S. J. (2004). Effects of crowding, isolation, and transfer from isolation to crowding on total ecdysteroid content of eggs in Schistocerca gregaria. Journal of Insect Physiology, 50 (7), 621–628.
Hardy, I. C. W, & Field, S. A (1998). Logistic analysis of animal contests. Animal Behaviour, 56 (3), 787–792.
Heifetz, Y., Voet, H., & Applebaum, S. W. (1996). Factors affecting behavioural phase transition in the desert locust, Schistocerca gregaria (Forskal) (Orthoptera: Acrididae). Journal of Chemical Ecology, 22 (9), 1717–1734.
40
Hodjat, S. H. (1970). Effects of crowding on colour, size and larval activity of Spodoptera littoralis (Lepidoptera: Noctuidae). Entomologia Experimentalis et
Applicata, 13 (1), 97–106.
Hoffman-Campo, C.B., Oliveira, E. B., & De Moscardi, F. (1985). Criação massal da lagarta da soja (Anticarsia gemmatalis). EMBRAPA.
Hosmer, D. W., Hosmer, T., Le Cessie, S., & Lemeshow, S. (1997). A comparison of goodness-of-fit tests for the logistic regression model. Statistics in Medicine, 16 (9), 965–980.
Howard, R. W., Blomquist, G. J. (2005). Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annual Review of Entomology, 50, 371-393.
Kaiser, S. A., Sillett, T. S., & Webster, M. S. (2014). Phenotypic plasticity in hormonal and behavioural responses to changes in resource conditions in a migratory songbird. Animal Behaviour, 96, 19–29.
Komers, P. E. (1997). Behavioural plasticity in variable environments. Canadian Journal of Zoology, 75 (2), 161–169.
Kowalski, K. N., Lakes-Harlan, R., Lehmann, G. U., & Strauß, J. (2014). Acoustic defence in an insect: characteristics of defensive stridulation and differences between the sexes in the tettigoniid Poecilimon ornatus (Schmidt 1850). Zoology, 117 (5), 329– 336.
Leo Lester, R., Grach, C., Paul Pener, M., & Simpson, S. J. (2005). Stimuli inducing gregarious colouration and behaviour in nymphs of Schistocerca gregaria. Journal of Insect Physiology, 51 (7), 737–747.
Ma, Z., Guo, W., Guo, X., Wang, X., & Kang, L. (2011). Modulation of behavioral phase changes of the migratory locust by the catecholamine metabolic pathway. Proceedings of the National Academy of Sciences of the United States of America,
41
Miner, B. G., Sultan, S. E., Morgan, S. G., Padilla, D. K., & Relyea, R. A. (2005). Ecological consequences of phenotypic plasticity. Trends in Ecology & Evolution, 20 (12), 685–692.
Nijhout, H. F. (1999). Control Mechanisms of Polyphenic Development in Insects. BioScience, 49 (3), 181–192.
Nijhout, H. F. (2003). Development and evolution of adaptive polyphenisms. Evolution & Development, 5 (1), 9–18.
Pener, M. P. (1991). Locust Phase Polymorphism and its Endocrine Relations. Advances in Insect Physiology, 23, 1–79.
Pener, M. P., & Yerushalmi, Y. (1998). The physiology of locust phase polymorphism : an update. Journal of Insect Physiology, 44 (5-6), 365–377.
Pfennig, D. W., Wund, M. A., Snell-Rood, E. C., Cruickshank, T., Schlichting, C. D., & Moczek, A. P. (2010). Phenotypic plasticity’s impacts on diversification and speciation. Trends in Ecology & Evolution, 25(8), 459–467.
Pigliucci, M. (2005). Evolution of phenotypic plasticity: where are we going now ? Trends in Ecology & Evolution, 20 (9), 481–486.
Praça, L. B., Moraes, S., & Monnerat, R. G. (2006). Anticarsia gemmatalis Hübner, 1818 (Lepidoptera:Noctuidae) biologia, amostragem e métodos de controle. Embrapa.
Prokopy, R. J., & Roitberg, B. D. (2001). Joining and avoidance behavior in nonsocial insects. Annual Review of Entomology, 46, 631–650.
Roessingh, P., Bouaïchi, A, & Simpson, S. J. (1998). Effects of sensory stimuli on the behavioural phase state of the desert locust, Schistocerca gregaria. Journal of Insect Physiology, 44 (10), 883–893.