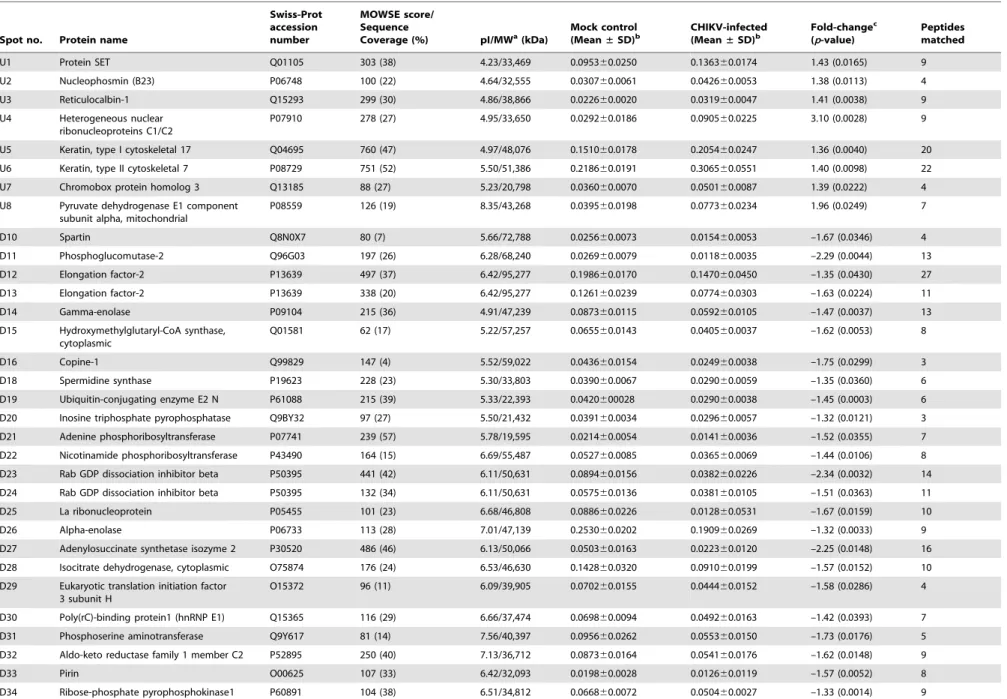
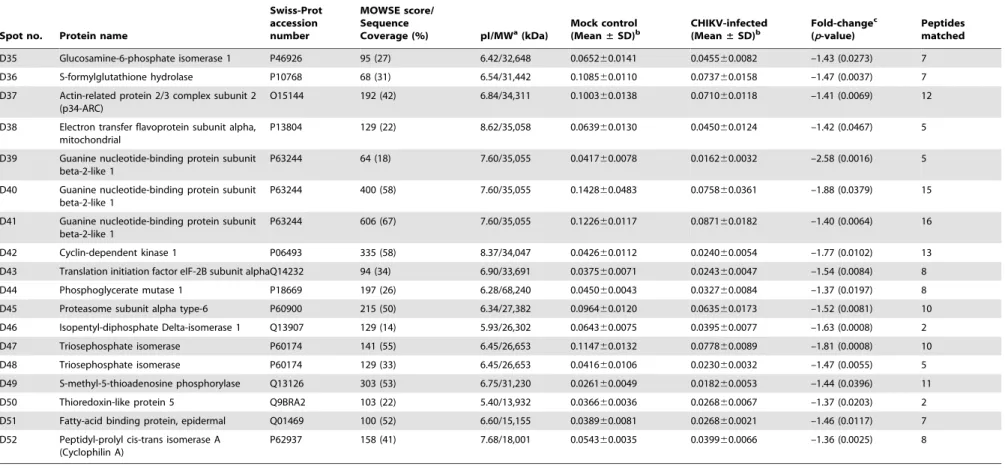
Differential proteome analysis of chikungunya virus infection on host cells.
Texto
Imagem

Documentos relacionados
Based on the Free Words Association Technique, data on evocations related to the Zika virus were modeled by social network analysis, allowing the characterization of the web
Com avanço e consolidação da área urbana ao redor do aeroporto, o aumento do número de pousos e decolagens, e da mudança na legislação sobre o zoneamento de ruído
The phenotypic diversity determined based on eight differential Brazilian cultivars was lower in commercial rice fields of ‘Primavera’ than in the fields of ‘Bonança,’
This paper reviews recent developments on biological processes developed on production of various products in solid state fermentation using sugarcane bagasse as the substrate
In the absence of targeted sequencing protocols for this novel virus, we applied a virus discovery strategy based on random amplification of purified nucleic acids in combination
Consistent with the cell cycle analysis, the results of EdU incorporation were not significantly different among P19CL6, P- c3.1 and P-499 cells at the early stage of
Correction for multiple testing using false discovery rate (FDR – q = 0.2) was performed separately in the unrelated, family and combined data accounting for 1,376 tests in
Multiple bioinformatics steps were utilized for data processing of MS/MS data by Spectrum Mill analyses for identification of DCSV proteins, FDR (false discovery rate) analyses
