Effect of temperature on embryonic development, larval viability and biomarkers in the first stages of life of Octopus Vulgaris (Cuvier, 1797)
46
0
0
Texto
Imagem
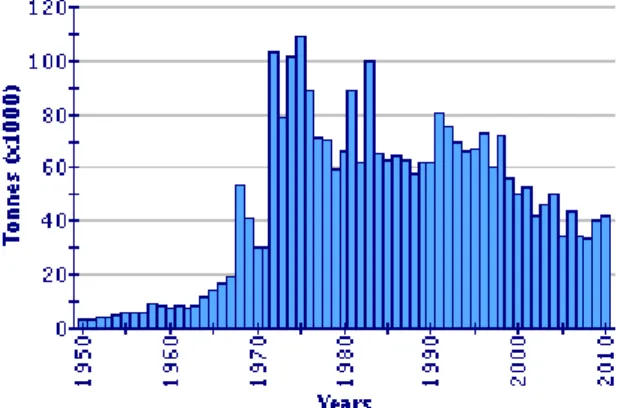
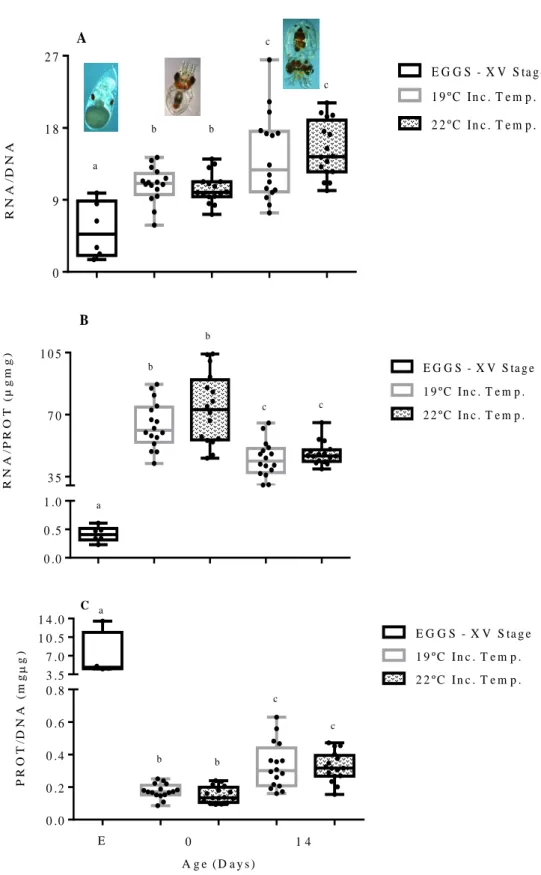
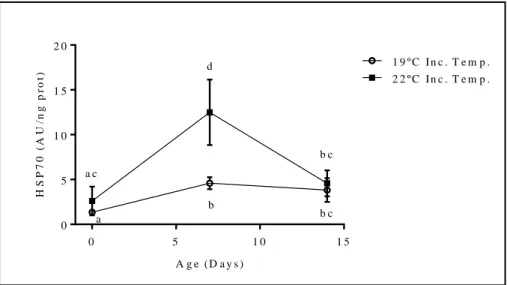
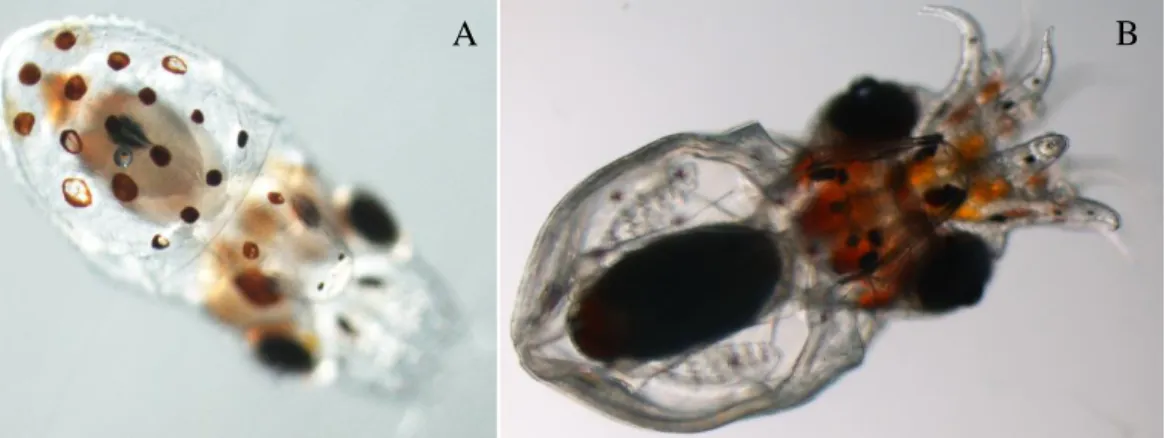
![Figure 3.3 - [A] Embryo of O. vulgaris at XIX – XX stage; [B] Premature paralarvae (stimulated to hatch); [C] Recently completed developed hatched paralarvae](https://thumb-eu.123doks.com/thumbv2/123dok_br/18602015.909340/14.892.82.808.898.1031/vulgaris-premature-paralarvae-stimulated-recently-completed-developed-paralarvae.webp)

+7



Documentos relacionados